
Integrated Genetic and Omics Approaches for the Regulation of Nutritional Activities in Rice (Oryza sativa L.)

Abstract
:1. Introduction
2. Nutritional Quality Enhancement by QTL Mapping in Rice
3. GWAS Analysis Improves Rice Nutritional Quality Traits
4. Efficient Nutrient-Rich Rice Breeding through Genome Selection (GS)
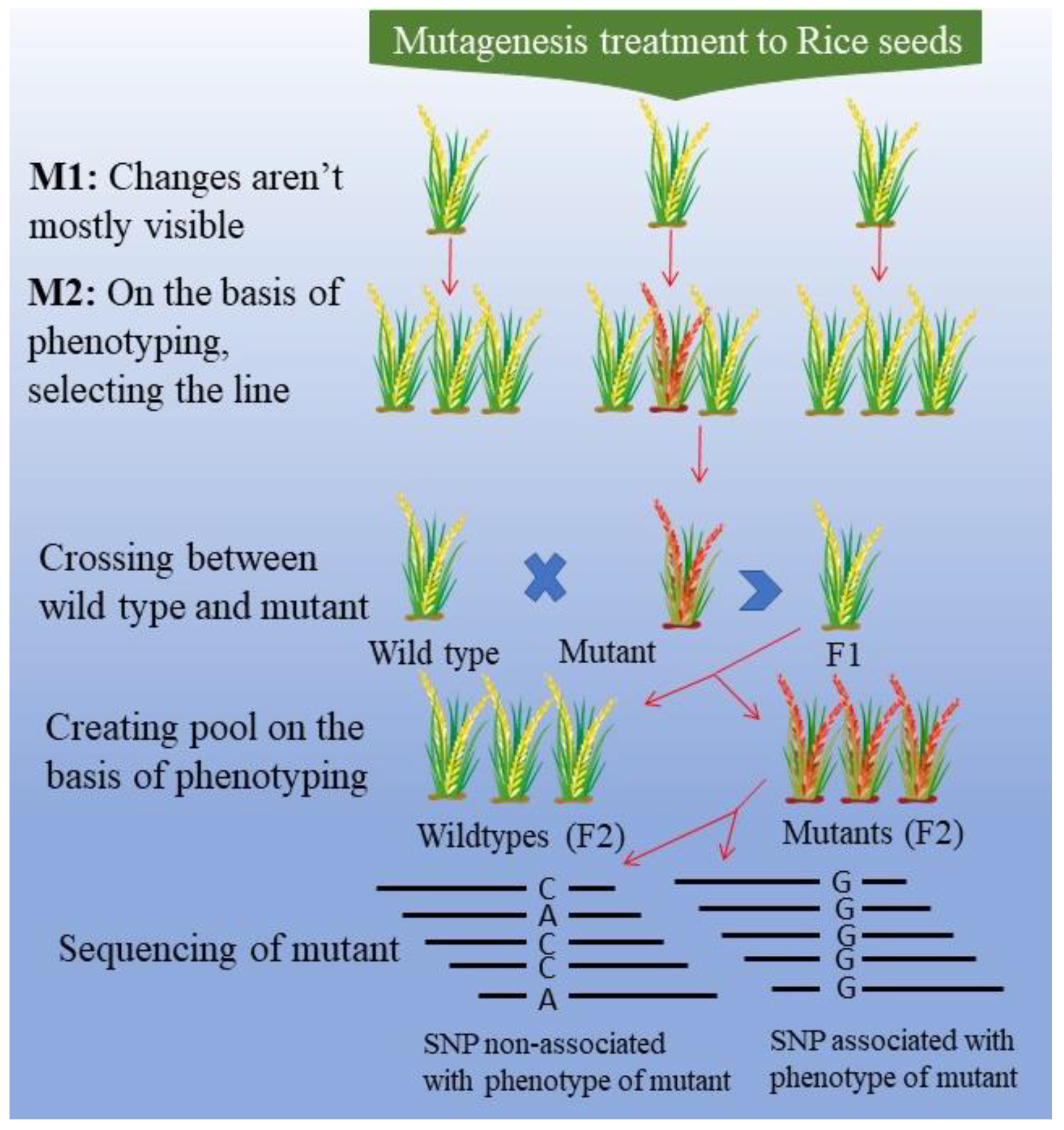
5. Mutation Mapping and Mutagenesis Techniques: Impact on Nutritional Quality of Rice
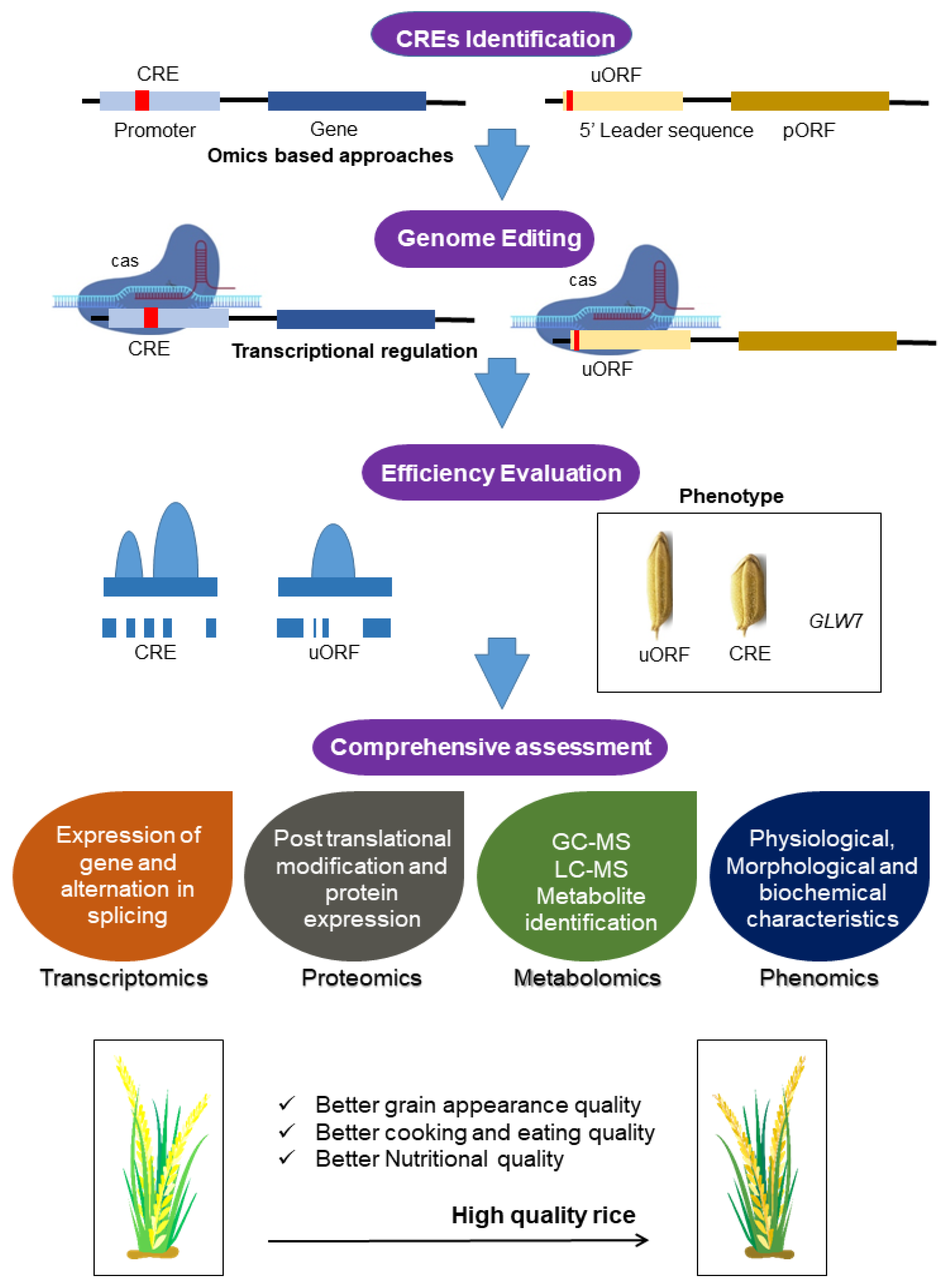
6. Integrative Omics Technologies for Enhancement of Rice Nutritional Quality Traits
6.1. Genomics and Pan-Genomics Analysis
6.2. Transcriptomics: Rice Nutritional Quality Enhancement through RNA Sequences
6.3. Proteomics: Rice Nutritional Quality Enhancement through Protein
6.4. Metabolomics: Rice Nutritional Quality Improvement through Metabolic Regulation
6.5. Nutrigenomics Approach in Rice
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Dobermann, A.; Witt, C.; Dawe, D.; Abdulrachman, S.; Gines, H.C.; Nagarajan, R.; Satawathananont, S.; Son, T.T.; Tan, P.S.; Wang, G.H.; et al. Site-specific nutrient management for intensive rice cropping systems in Asia. Field Crops Res. 2002, 74, 37–66. [Google Scholar] [CrossRef]
- Yankah, N.; Intiful, F.D.; Tette, E.M.A. Comparative study of the nutritional composition of local brown rice, maize (obaatanpa), and millet—A baseline research for varietal complementary feeding. Food Sci. Nutr. 2020, 8, 2692–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholova, J.; Sairam, R.K.; Meena, R.C. Osmolytes and metal ions accumulation, oxidative stress and antioxidant enzymes activity as determinants of salinity stress tolerance in maize genotypes. Acta Physiol. Plant. 2010, 32, 477–486. [Google Scholar] [CrossRef]
- Zakaria, Z.; Zulkafflee, N.S.; Mohd Redzuan, N.A.; Selamat, J.; Ismail, M.R.; Praveena, S.M.; Tóth, G.; Abdull Razis, A.F. Understanding potential heavy metal contamination, absorption, translocation and accumulation in rice and human health risks. Plants 2021, 10, 1070. [Google Scholar] [CrossRef]
- Ye, J.; Hu, X.; Luo, S.; McClements, D.J.; Liang, L.; Liu, C. Effect of endogenous proteins and lipids on starch digestibility in rice flour. Food Res. Int. 2018, 106, 404–409. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Meng, Q.; Yu, J.; Weng, L.; Li, J.; Deng, L.; Zhu, Q.; Gu, X.; Chen, C.; et al. Phenotypic and genetic dissection of cadmium accumulation in roots, nodes and grains of rice hybrids. Plant Soil 2021, 463, 39–53. [Google Scholar] [CrossRef]
- Noor, J.J.; Vinayan, M.T.; Umar, S.; Devi, P.; Iqbal, M.; Seetharam, K.; Zaidi, P.H. Morpho-physiological traits associated with heat stress tolerance in tropical maize (Zea mays L.) at the reproductive stage. Aust. J. Crop Sci. 2019, 13, 536–545. [Google Scholar] [CrossRef]
- Simkin, A.J. Genetic engineering for global food security: Photosynthesis and biofortification. Plants 2019, 8, 586. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yang, X.; Tzin, V.; Peng, Y.; Romeis, J.; Li, Y. Plant breeding involving genetic engineering does not result in unacceptable unintended effects in rice relative to conventional cross-breeding. Plant J. 2020, 103, 2236–2249. [Google Scholar] [CrossRef] [PubMed]
- Paine, J.A.; Shipton, C.A.; Chaggar, S.; Howells, R.M.; Kennedy, M.J.; Vernon, G.; Wright, S.Y.; Hinchliffe, E.; Adams, J.L.; Silverstone, A.L.; et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nat. Biotechnol. 2005, 23, 482–487. [Google Scholar] [CrossRef]
- Chen, H.; Lin, Y. Promise and issues of genetically modified crops. Curr. Opin. Plant Biol. 2013, 16, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I. Ionomics: Studying the social network of mineral nutrients. Curr. Opin. Plant Biol. 2009, 12, 381–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phing Lau, W.C.; Latif, M.A.; Rafii, M.Y.; Ismail, M.R.; Puteh, A. Advances to improve the eating and cooking qualities of rice by marker-assisted breeding. Crit. Rev. Biotechnol. 2016, 36, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, L.; Robb, S.M.C.; Collin, M.; Okumoto, Y.; Stajich, J.E.; Wessler, S.R. Genomic diversity generated by a transposable element burst in a rice recombinant inbred population. Proc. Natl. Acad. Sci. USA 2020, 117, 26288–26297. [Google Scholar] [CrossRef]
- Fragoso, C.A.; Moreno, M.; Wang, Z.; Heffelfinger, C.; Arbelaez, L.J.; Aguirre, J.A.; Franco, N.; Romero, L.E.; Labadie, K.; Zhao, H.; et al. Genetic architecture of a rice nested association mapping population. G3 Genes Genomes Genet. 2017, 7, 1913–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zou, W.; Lu, X.; Bian, J.; He, H.; Chen, J.; Ye, G. Genome-wide association study using a multiparent advanced generation intercross (Magic) population identified qtls and candidate genes to predict shoot and grain zinc contents in rice. Agriculture 2021, 11, 70. [Google Scholar] [CrossRef]
- Liu, C.; Ding, S.; Zhang, A.; Hong, K.; Jiang, H.; Yang, S.; Ruan, B.; Zhang, B.; Dong, G.; Guo, L.; et al. Development of nutritious rice with high zinc/selenium and low cadmium in grains through QTL pyramiding. J. Integr. Plant Biol. 2020, 62, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Ye, G.; Liang, S.; Wan, J. QTL mapping of protein content in rice using single chromosome segment substitution lines. Theor. Appl. Genet. 2010, 121, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Tondi Yacouba, N.; Chen, J.; Wang, Y.; Gao, G.; Zhang, Q.; Liu, X.; He, Y. Analysis of minor quantitative trait loci for eating and cooking quality traits in rice using a recombinant inbred line population derived from two indica cultivars with similar amylose content. Mol. Breed. 2014, 34, 2151–2163. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Kaladhar, K.; Anuradha, K.; Batchu, A.K.; Longvah, T.; Sarla, N. QTL Analysis for Grain Iron and Zinc Concentrations in Two O. nivara Derived Backcross Populations. Rice Sci. 2018, 25, 197–207. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Rahman, M.A.; Inabangan-Asilo, M.A.; Amparado, A.; Manito, C.; Chadha-Mohanty, P.; Reinke, R.; Slamet-Loedin, I.H. Advances in breeding for high grain Zinc in Rice. Rice 2016, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Bharati, B.; Chatopadhyay, K.; Singh, S.; Dash, P.; Reddy, J.N. Association mapping reveals multiple QTLs for grain protein content in rice useful for biofortification. Mol. Genet. Genom. 2019, 294, 963–983. [Google Scholar] [CrossRef]
- Pandit, E.; Tasleem, S.; Barik, S.R.; Mohanty, D.P.; Nayak, D.K.; Mohanty, S.P.; Das, S.; Pradhan, S.K. Genome-wide association mapping reveals multiple QTLs governing tolerance response for seedling stage chilling stress in Indica rice. Front. Plant Sci. 2017, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.K.; Pandit, E.; Pawar, S.; Naveenkumar, R.; Barik, S.R.; Mohanty, S.P.; Nayak, D.K.; Ghritlahre, S.K.; Sanjiba Rao, D.; Reddy, J.N.; et al. Linkage disequilibrium mapping for grain Fe and Zn enhancing QTLs useful for nutrient dense rice breeding. BMC Plant Biol. 2020, 20, 57. [Google Scholar] [CrossRef] [Green Version]
- Raza, Q.; Riaz, A.; Sabar, M.; Atif, R.M.; Bashir, K. Meta-analysis of grain iron and zinc associated QTLs identified hotspot chromosomal regions and positional candidate genes for breeding biofortified rice. Plant Sci. 2019, 288, 110214. [Google Scholar] [CrossRef]
- Smita, S.; Katiyar, A.; Lenka, S.K.; Dalal, M.; Kumar, A.; Mahtha, S.K.; Yadav, G.; Chinnusamy, V.; Pandey, D.M.; Bansal, K.C. Gene network modules associated with abiotic stress response in tolerant rice genotypes identified by transcriptome meta-analysis. Funct. Integr. Genom. 2020, 20, 29–49. [Google Scholar] [CrossRef]
- Norton, G.J.; Deacon, C.M.; Xiong, L.; Huang, S.; Meharg, A.A.; Price, A.H. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 2010, 329, 139–153. [Google Scholar] [CrossRef]
- Hong, W.J.; Yoo, Y.H.; Park, S.A.; Moon, S.; Kim, S.R.; An, G.; Jung, K.H. Genome-wide identification and extensive analysis of rice-endosperm preferred genes using reference expression database. J. Plant Biol. 2017, 60, 249–258. [Google Scholar] [CrossRef]
- Kishor, D.S.; Lee, C.; Lee, D.; Venkatesh, J.; Seo, J.; Chin, J.H.; Jin, Z.; Hong, S.K.; Ham, J.K.; Koh, H.J. Novel allelic variant of Lpa1 gene associated with a significant reduction in seed phytic acid content in rice (Oryza sativa L.). PLoS ONE 2019, 14, e209636. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhu, C.; Pang, J.; Zhang, X.; Yang, C.; Xia, G.; Tian, Y.; He, C. OsLOL1, a C2C2-type zinc finger protein, interacts with OsbZIP58 to promote seed germination through the modulation of gibberellin biosynthesis in Oryza sativa. Plant J. 2014, 80, 1118–1130. [Google Scholar] [CrossRef]
- Lee, S.; Jeon, U.S.; Lee, S.J.; Kim, Y.K.; Persson, D.P.; Husted, S.; Schjørring, J.K.; Kakei, Y.; Masuda, H.; Nishizawa, N.K.; et al. Iron fortification of rice seeds through activation of the nicotianamine synthase gene. Proc. Natl. Acad. Sci. USA 2009, 106, 22014–22019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xu, Y.H.; Yi, H.Y.; Gong, J.M. Vacuolar membrane transporters OsVIT1 and OsVIT2 modulate iron translocation between flag leaves and seeds in rice. Plant J. 2012, 72, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Saika, H.; Oikawa, A.; Matsuda, F.; Onodera, H.; Saito, K.; Toki, S. Application of gene targeting to designed mutation breeding of high-tryptophan rice. Plant Physiol. 2011, 156, 1269–1277. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Aung, M.S.; Nishizawa, N.K. Iron biofortification of rice using different transgenic approaches. Rice 2013, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Wang, X.; Zhou, D.; Ouyang, Y.; Yao, J. Overexpression of the 16-kDa α-amylase/trypsin inhibitor RAG2 improves grain yield and quality of rice. Plant Biotechnol. J. 2017, 15, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Zhang, C.Q.; Chan, M.L.; Zhao, D.S.; Chen, J.Z.; Wang, Q.; Li, Q.F.; Yu, H.X.; Gu, M.H.; Sun, S.S.M.; et al. Biofortification of rice with the essential amino acid lysine: Molecular characterization, nutritional evaluation, and field performance. J. Exp. Bot. 2016, 67, 4285–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.H.; Zhao, H.J.; Liu, Q.L.; Frank, T.; Engel, K.H.; An, G.; Shu, Q.Y. Mutations of the multi-drug resistance-associated protein ABC transporter gene 5 result in reduction of phytic acid in rice seeds. Theor. Appl. Genet. 2009, 119, 75–83. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Ma, A.; Xie, L.; Ramachandran, S. Improving protein content and quality by over-expressing artificially synthetic fusion proteins with high lysine and threonine constituent in rice plants. Sci. Rep. 2016, 6, 34427. [Google Scholar] [CrossRef] [Green Version]
- Blancquaert, D.; Van Daele, J.; Strobbe, S.; Kiekens, F.; Storozhenko, S.; De Steur, H.; Gellynck, X.; Lambert, W.; Stove, C.; Van Der Straeten, D. Improving folate (Vitamin B 9) stability in biofortified rice through metabolic engineering. Nat. Biotechnol. 2015, 33, 1076–1078. [Google Scholar] [CrossRef]
- Lei, L.; Zheng, H.L.; Wang, J.G.; Liu, H.L.; Sun, J.; Zhao, H.W.; Yang, L.M.; Zou, D.T. Genetic dissection of rice (Oryza sativa L.) tiller, plant height, and grain yield based on QTL mapping and metaanalysis. Euphytica 2018, 214, 109. [Google Scholar] [CrossRef]
- Korte, A.; Ashley, F. The advantages and limitations of trait analysis with GWAS: A review Self-fertilisation makes Arabidopsis particularly well suited to GWAS. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Xu, K.; Li, Z.; Hu, Y.; Hu, Z.; Zheng, X.; Song, S.; Tang, Z.; Li, L. Genome-wide association study and Mendelian randomization analysis provide insights for improving rice yield potential. Sci. Rep. 2021, 11, 6894. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A. A strategy for identifying quantitative trait genes using gene expression analysis and causal analysis. Genes 2017, 8, 347. [Google Scholar] [CrossRef] [Green Version]
- Norton, G.J.; Douglas, A.; Lahner, B.; Yakubova, E.; Guerinot, M.L.; Pinson, S.R.M.; Tarpley, L.; Eizenga, G.C.; McGrath, S.P.; Zhao, F.J.; et al. Genome wide association mapping of grain arsenic, copper, molybdenum and zinc in rice (Oryza sativa L.) grown at four international field sites. PLoS ONE 2014, 9, e89685. [Google Scholar] [CrossRef] [Green Version]
- Eizenga, G.C.; Jackson, A.K.; Edwards, J.D. Prototype for developing SNP markers from GWAS and biparental QTL for rice panicle and grain traits. Agric. Environ. Lett. 2021, 6, e20047. [Google Scholar] [CrossRef]
- Zhang, P.; Zhong, K.; Zhong, Z.; Tong, H. Mining candidate gene for rice aluminum tolerance through genome wide association study and transcriptomic analysis. BMC Plant Biol. 2019, 19, 490. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.-L. Plant Marker-Assisted Breeding and Conventional Breeding: Challenges and Perspectives. Adv. Crop Sci. Technol. 2013, 1, e106. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Zhao, K.; Wright, M.; Kimball, J.; Eizenga, G.; McClung, A.; Kovach, M.; Tyagi, W.; Ali, M.L.; Tung, C.W.; Reynolds, A.; et al. Genomic diversity and introgression in O. sativa reveal the impact of domestication and breeding on the rice genome. PLoS ONE 2010, 5, e10780. [Google Scholar] [CrossRef]
- Spindel, J.; Begum, H.; Akdemir, D.; Virk, P.; Collard, B.; Redoña, E.; Atlin, G.; Jannink, J.L.; McCouch, S.R. Correction: Genomic Selection and Association Mapping in Rice (Oryza sativa): Effect of Trait Genetic Architecture, Training Population Composition, Marker Number and Statistical Model on Accuracy of Rice Genomic Selection in Elite, Tropical Rice Breeding. PLoS Genet. 2015, 11, e1005350. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; He, H.; Zhou, F.; Yu, H.; Deng, X.W. Development of genomics-based genotyping platforms and their applications in rice breeding. Curr. Opin. Plant Biol. 2013, 16, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Su, G.; Jiang, L.; Bao, Z. An efficient unified model for genome-wide association studies and genomic selection. Genet. Sel. Evol. 2017, 49, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spindel, J.E.; Begum, H.; Akdemir, D.; Collard, B.; Redoña, E.; Jannink, J.L.; McCouch, S. Genome-wide prediction models that incorporate de novo GWAS are a powerful new tool for tropical rice improvement. Heredity 2016, 116, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Rathna Priya, T.S.; Raeboline, A.; Eliazer, L.; Ravichandran, K.; Antony, U. Nutritional and functional properties of coloured rice varieties of South India: A review. J. Ethn. Foods 2019, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Descalsota, E.G.I.; Noraziyah, A.A.S.; Navea, I.P.; Chung, C.; Dwiyanti, M.S.; Labios, R.J.D.; Ikmal, A.M.; Juanillas, V.M.; Inabangan-Asilo, M.A.; Amparado, A.; et al. Genetic dissection of grain nutritional traits and leaf blight resistance in rice. Genes 2019, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Bollinedi, H.; Yadav, A.K.; Vinod, K.K.; Gopala Krishnan, S.; Bhowmick, P.K.; Nagarajan, M.; Neeraja, C.N.; Ellur, R.K.; Singh, A.K. Genome-Wide Association Study Reveals Novel Marker-Trait Associations (MTAs) Governing the Localization of Fe and Zn in the Rice Grain. Front. Genet. 2020, 11, 213. [Google Scholar] [CrossRef] [Green Version]
- Swamy, B.P.M.; Shamsudin, N.A.A.; Rahman, S.N.A.; Mauleon, R.; Ratnam, W.; St. Cruz, M.T.; Kumar, A. Association Mapping of Yield and Yield-related Traits under Reproductive Stage Drought Stress in Rice (Oryza sativa L.). Rice 2017, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Grenier, C.; Cao, T.V.; Ospina, Y.; Quintero, C.; Châtel, M.H.; Tohme, J.; Courtois, B.; Ahmadi, N. Accuracy of genomic selection in a rice synthetic population developed for recurrent selection breeding. PLoS ONE 2015, 10, e136594. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zhu, D.; Zhang, Q. Predicting hybrid performance in rice using genomic best linear unbiased prediction. Proc. Natl. Acad. Sci. USA 2014, 111, 12456–12461. [Google Scholar] [CrossRef]
- Onogi, A.; Watanabe, M.; Mochizuki, T.; Hayashi, T.; Nakagawa, H.; Hasegawa, T.; Iwata, H. Toward integration of genomic selection with crop modelling: The development of an integrated approach to predicting rice heading dates. Theor. Appl. Genet. 2016, 129, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Mba, C.; Afza, R.; Bado, S.; Jain, S.M. Induced Mutagenesis in Plants using physical and chemical agents. Plant Cell Cult. Essent. Methods 2010, 20, 111–130. [Google Scholar]
- Phanchaisri, B.; Chandet, R.; Yu, L.D.; Vilaithong, T.; Jamjod, S.; Anuntalabhochai, S. Low-energy ion beam-induced mutation in Thai jasmine rice (Oryza sativa L. cv. KDML 105). Surf. Coat. Technol. 2007, 201, 8024–8028. [Google Scholar] [CrossRef]
- Semsang, N.; Kawaree, R.; Cutler, R.W.; Chundet, R.; Yu, L.D.; Anuntalabhochai, S. Improved antioxidant activity of BKOS Thai jasmine rice. Nat. Prod. Res. 2012, 26, 1145–1151. [Google Scholar] [CrossRef]
- Liu, Q.L.; Xu, X.H.; Ren, X.L.; Fu, H.W.; Wu, D.X.; Shu, Q.Y. Generation and characterization of low phytic acid germplasm in rice (Oryza sativa L.). Theor. Appl. Genet. 2007, 114, 803–814. [Google Scholar] [CrossRef]
- Zhou, X.; Bai, X.; Xing, Y. A rice genetic improvement boom by next-generation sequencing. Curr. Issues Mol. Biol. 2018, 27, 109–126. [Google Scholar] [CrossRef]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Takagi, H.; Uemura, A.; Yaegashi, H.; Tamiru, M.; Abe, A.; Mitsuoka, C.; Utsushi, H.; Natsume, S.; Kanzaki, H.; Matsumura, H.; et al. MutMap-Gap: Whole-genome resequencing of mutant F2 progeny bulk combined with de novo assembly of gap regions identifies the rice blast resistance gene Pii. New Phytol. 2013, 200, 276–283. [Google Scholar] [CrossRef]
- Peng, H.; Wang, K.; Chen, Z.; Cao, Y.; Gao, Q.; Li, Y.; Li, X.; Lu, H.; Du, H.; Lu, M.; et al. MBKbase for rice: An integrated omics knowledgebase for molecular breeding in rice. Nucleic Acids Res. 2020, 48, D1085–D1092. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Qi, M.; Liu, J.; Zhang, Y. CARMO: A comprehensive annotation platform for functional exploration of rice multi-omics data. Plant J. 2015, 83, 359–374. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, S.; Satish Chhapekar, S.; Sreedharan, S.; Singh, S.; Ryun Choi, S.; Ramchiary, N.; Pyo Lim, Y. Integrating omics and gene editing tools for rapid improvement of traditional food plants for diversified foods and sustainable food security. Int. J. Mol. Sci. 2021, 22, 8093. [Google Scholar] [CrossRef]
- Li, Q.; Yan, J. Sustainable agriculture in the era of omics: Knowledge-driven crop breeding. Genome Biol. 2020, 21, 5–9. [Google Scholar] [CrossRef]
- Matsumoto, T.; Wu, J.; Kanamori, H.; Katayose, Y.; Fujisawa, M.; Namiki, N.; Mizuno, H.; Yamamoto, K.; Antonio, B.A.; Baba, T.; et al. The map-based sequence of the rice genome. Nature 2005, 436, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Wenger, A.M.; Peluso, P.; Rowell, W.J.; Chang, P.C.; Hall, R.J.; Concepcion, G.T.; Ebler, J.; Fungtammasan, A.; Kolesnikov, A.; Olson, N.D.; et al. Accurate circular consensus long-read sequencing improves variant detection and assembly of a human genome. Nat. Biotechnol. 2019, 37, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chebotarov, D.; Kudrna, D.; Llaca, V.; Lee, S.; Rajasekar, S.; Mohammed, N.; Al-Bader, N.; Sobel-Sorenson, C.; Parakkal, P.; et al. A platinum standard pan-genome resource that represents the population structure of Asian rice. Sci. Data 2020, 7, 113. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Wang, C.H.; Xu, Q.; Feng, Y.; Yuan, X.P.; Yu, H.Y.; Wang, Y.P.; Tang, S.X.; Wei, X.H. Genome-wide copy number variations in Oryza sativa L. BMC Genom. 2013, 14, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrell, P.L.; Buckler, E.S.; Ross-Ibarra, J. Crop genomics: Advances and applications. Nat. Rev. Genet. 2012, 13, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.C.; Yu, Y.; Copetti, D.; Zwickl, D.J.; Zhang, L.; Zhang, C.; Chougule, K.; Gao, D.; Iwata, A.; Goicoechea, J.L.; et al. Genomes of 13 domesticated and wild rice relatives highlight genetic conservation, turnover and innovation across the genus Oryza. Nat. Genet. 2018, 50, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Mondal, T.K.; Rawal, H.C.; Chowrasia, S.; Varshney, D.; Panda, A.K.; Mazumdar, A.; Kaur, H.; Gaikwad, K.; Sharma, T.R.; Singh, N.K. Draft genome sequence of first monocot-halophytic species Oryza coarctata reveals stress-specific genes. Sci. Rep. 2018, 8, 13698. [Google Scholar] [CrossRef]
- Xie, X.; Du, H.; Tang, H.; Tang, J.; Tan, X.; Liu, W.; Li, T.; Lin, Z.; Liang, C.; Liu, Y.G. A chromosome-level genome assembly of the wild rice Oryza rufipogon facilitates tracing the origins of Asian cultivated rice. Sci. China Life Sci. 2021, 64, 282–293. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Feng, Q.; Lu, H.; Li, Y.; Wang, A.; Tian, Q.; Zhan, Q.; Lu, Y.; Zhang, L.; Huang, T.; et al. Pan-genome analysis highlights the extent of genomic variation in cultivated and wild rice. Nat. Genet. 2018, 50, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; et al. Rational design of high-yield and superior-quality rice. Nat. Plants 2017, 3, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, T.; Meng, X.; Du, H.; Zhang, J.; Liu, G.; Chen, M.; Jing, Y.; Kou, L.; Li, X.; et al. A route to de novo domestication of wild allotetraploid rice. Cell 2021, 184, 1156–1170. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zou, D.; Sang, J.; Xu, X.; Yin, H.; Li, M.; Wu, S.; Hu, S.; Hao, L.; Zhang, Z. Rice Expression Database (RED): An integrated RNA-Seq-derived gene expression database for rice. J. Genet. Genom. 2017, 44, 235–241. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Qi, D.; Pugh, N.; Li, K.; Wen, B.; Zhou, R.; Xu, S.; Liu, S.; Jones, A.R. Improvements to the rice genome annotation through large-scale analysis of RNA-seq and proteomics data sets. Mol. Cell. Proteom. 2019, 18, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Huang, F.; Narsai, R.; Wu, J.; Giraud, E.; He, F.; Cheng, L.; Wang, F.; Wu, P.; Whelan, J.; et al. Physiological and transcriptome analysis of iron and phosphorus interaction in rice seedlings. Plant Physiol. 2009, 151, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Anacleto, R.; Badoni, S.; Parween, S.; Butardo, V.M.; Misra, G.; Cuevas, R.P.; Kuhlmann, M.; Trinidad, T.P.; Mallillin, A.C.; Acuin, C.; et al. Integrating a genome-wide association study with a large-scale transcriptome analysis to predict genetic regions influencing the glycaemic index and texture in rice. Plant Biotechnol. J. 2019, 17, 1261–1275. [Google Scholar] [CrossRef] [Green Version]
- Takehisa, H.; Sato, Y.; Igarashi, M.; Abiko, T.; Antonio, B.A.; Kamatsuki, K.; Minami, H.; Namiki, N.; Inukai, Y.; Nakazono, M.; et al. Genome-wide transcriptome dissection of the rice root system: Implications for developmental and physiological functions. Plant J. 2012, 69, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Lei, G.; Zhou, H.W.; He, C.; Liao, J.L.; Huang, Y.J. Quantitative iTRAQ-based proteomic analysis of rice grains to assess high night temperature stress. Proteomics 2017, 17, 1600365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksup, S.; Pongpakpian, S.; Roytrakul, S.; Chaum, S.; Supaibulwatana, K. Comparative proteomics and protein profile related to phenolic compounds and antioxidant activity in germinated Oryza sativa KDML105 and Thai brown rice Mali Dang for better nutritional value. J. Sci. Food Agric. 2018, 28, 303–325. [Google Scholar] [CrossRef]
- Graziano, S.; Marmiroli, N.; Gullì, M. Proteomic analysis of reserve proteins in commercial rice cultivars. Food Sci. Nutr. 2020, 8, 1788–1797. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, R.A.; Boff, T.; Duarte, G.L.; Santos, L.S.; Grusak, M.A.; Fett, J.P. Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. J. Plant Physiol. 2010, 167, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.W.; Komatsu, S.; Furukawa, T.; Koshiba, T.; Kohno, Y. Proteome analysis of proteins responsive to ambient and elevated ozone in rice seedlings. Agric. Ecosyst. Environ. 2008, 125, 255–265. [Google Scholar] [CrossRef]
- Ernst, M.; Silva, D.B.; Silva, R.R.; Vêncio, R.Z.N.; Lopes, N.P. Mass spectrometry in plant metabolomics strategies: From analytical platforms to data acquisition and processing. Nat. Prod. Rep. 2014, 31, 784–806. [Google Scholar] [CrossRef]
- Heuberger, A.L.; Lewis, M.R.; Chen, M.H.; Brick, M.A.; Leach, J.E.; Ryan, E.P. Metabolomic and functional genomic analyses reveal varietal differences in bioactive compounds of cooked rice. PLoS ONE 2010, 5, e12915. [Google Scholar] [CrossRef]
- Kusano, M.; Fukushima, A.; Kobayashi, M.; Hayashi, N.; Jonsson, P.; Moritz, T.; Ebana, K.; Saito, K. Application of a metabolomic method combining one-dimensional and two-dimensional gas chromatography-time-of-flight/mass spectrometry to metabolic phenotyping of natural variants in rice. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 855, 71–79. [Google Scholar] [CrossRef]
- Gong, L.; Chen, W.; Gao, Y.; Liu, X.; Zhang, H.; Xu, C.; Yu, S.; Zhang, Q.; Luo, J. Genetic analysis of the metabolome exemplified using a rice population. Proc. Natl. Acad. Sci. USA 2013, 110, 20320–20325. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Shi, J.; Quan, S.; Cui, B.; Kleessen, S.; Nikoloski, Z.; Tohge, T.; Alexander, D.; Guo, L.; Lin, H.; et al. Metabolic variation between japonica and indica rice cultivars as revealed by non-targeted metabolomics. Sci. Rep. 2014, 4, 5067. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. A comparative UHPLC-QqQ-MS-based metabolomics approach for evaluating Chinese and North American wild rice. Food Chem. 2019, 275, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.C.; Zhang, Y.X.; Sun, S.Y.; Xie, M.X.; Hu, C.Y.; Shi, Y.Q.; Shi, J.X.; Li, J.Y. Identification of the biochemical characteristics of developing giant embryo rice grains using non-targeted metabolomics. J. Cereal Sci. 2019, 85, 70–76. [Google Scholar] [CrossRef]
- Ravichanthiran, K.; Ma, Z.F.; Zhang, H.; Cao, Y.; Wang, C.W.; Muhammad, S.; Aglago, E.K.; Zhang, Y.; Jin, Y.; Pan, B. Phytochemical profile of brown rice and its nutrigenomic implications. Antioxidants 2018, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Romero, C.; Lopez-Perez, P.A.; Toledo, A.K.M.; Perez-Espana, V.H.; Aparicio-Burgos, J.E.; Cuervo-Parra, J.A. Nutrigenomics and Nutrigenetics in Functional Foods. Int. J. Bio-Resour. Stress Manag. 2018, 9, 661–672. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Function | Locus | References |
|---|---|---|---|
| glu4a | Gene involved in storage proteins of seed | Os01g55690 | [28] |
| lpa1 | Metabolizes the phytic acid | Os02g57400 | [29] |
| OsbZIP58, OsSMF1 | Helps in accumulating the storage protein | Os07g08420 | [30] |
| OsNAS3 | Improves the fortification of iron in rice seed | Os07g48980 | [31] |
| OsVIT2 | Involved in translocation of iron | Os09g23300 | [32] |
| OASA2 | Synthesis and accumulation of Tryptophan | [33] | |
| OsYSL2 | Transportation of manganese and iron at long distance | Os02g43370 | [34] |
| RAG2 | Functioning in yield and grain quality | Os07g11380 | [35] |
| LRP, RLRH1, and RLRH2 | Accumulation of lysin content | [36] | |
| XS-lpa2-1 | Involved with phytic acid accumulation | Os03g04920 | [37] |
| TKTKK1 and TKTKK2 | Regulation and synthesis of Methionine and cysteine | [38] | |
| AtGTPCH | Synthesis folate | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaghum, M.J.; Ali, K.; Teng, S. Integrated Genetic and Omics Approaches for the Regulation of Nutritional Activities in Rice (Oryza sativa L.). Agriculture 2022, 12, 1757. https://doi.org/10.3390/agriculture12111757
Zaghum MJ, Ali K, Teng S. Integrated Genetic and Omics Approaches for the Regulation of Nutritional Activities in Rice (Oryza sativa L.). Agriculture. 2022; 12(11):1757. https://doi.org/10.3390/agriculture12111757
Chicago/Turabian StyleZaghum, Muhammad Junaid, Kashir Ali, and Sheng Teng. 2022. "Integrated Genetic and Omics Approaches for the Regulation of Nutritional Activities in Rice (Oryza sativa L.)" Agriculture 12, no. 11: 1757. https://doi.org/10.3390/agriculture12111757