
Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Condition
2.2. Determination of NH4+ and NO3−
2.3. Determination of Nitrate Reductase (NR) Activity
2.4. Determination of Glutamine Synthetase (GS), Glutamate Synthase Activity (GOGAT) and Glutamate Dehydrogenase (GDH) Activity
2.5. Glutamate Oxaloacetate Aminotransferase (GOT) and Glutamate Pyruvate Aminotransferase (GPT) Activities
2.6. Dry Matter Accumulation and Determination of Root Characteristic Parameters
2.7. Determination of Root Activity and Root Exudate
2.8. Determination of SPAD Value and Net Photosynthetic Rate
2.9. Determination of Leaf Area Index and Leaf Senescence Characteristics
2.10. Determination of Yield
2.11. Data Analysis
3. Results
3.1. NO3− Concentration in Xylem Bleeding Sap
3.2. Contents of NO3− and NH4+ in Leaves
3.3. NR Activity in Leaves
3.4. Activities of GS, GOGAT and GDH in Leaves
3.5. Activities of GOT and GPT in Leaves
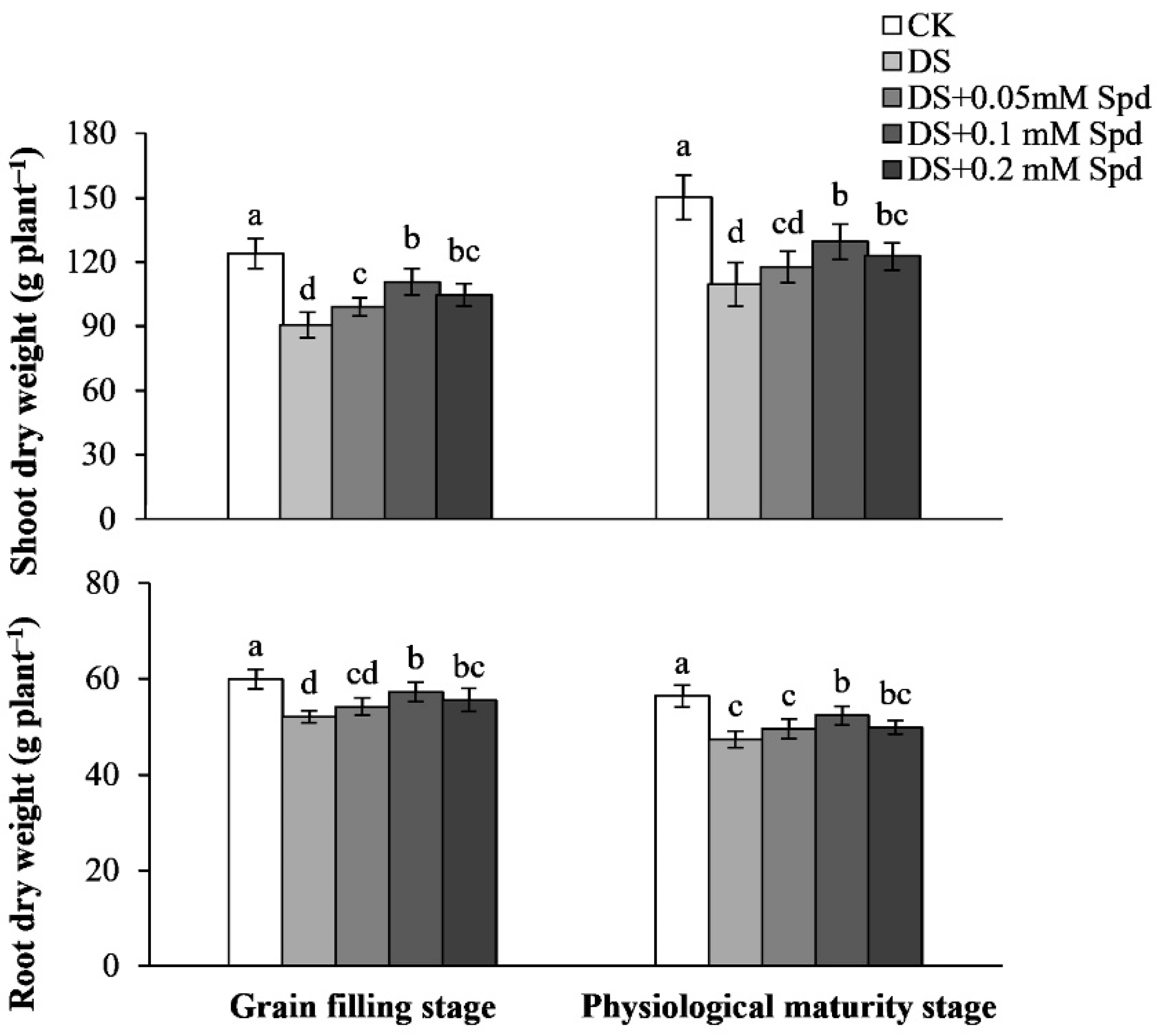
3.6. Dry Matter Accumulation
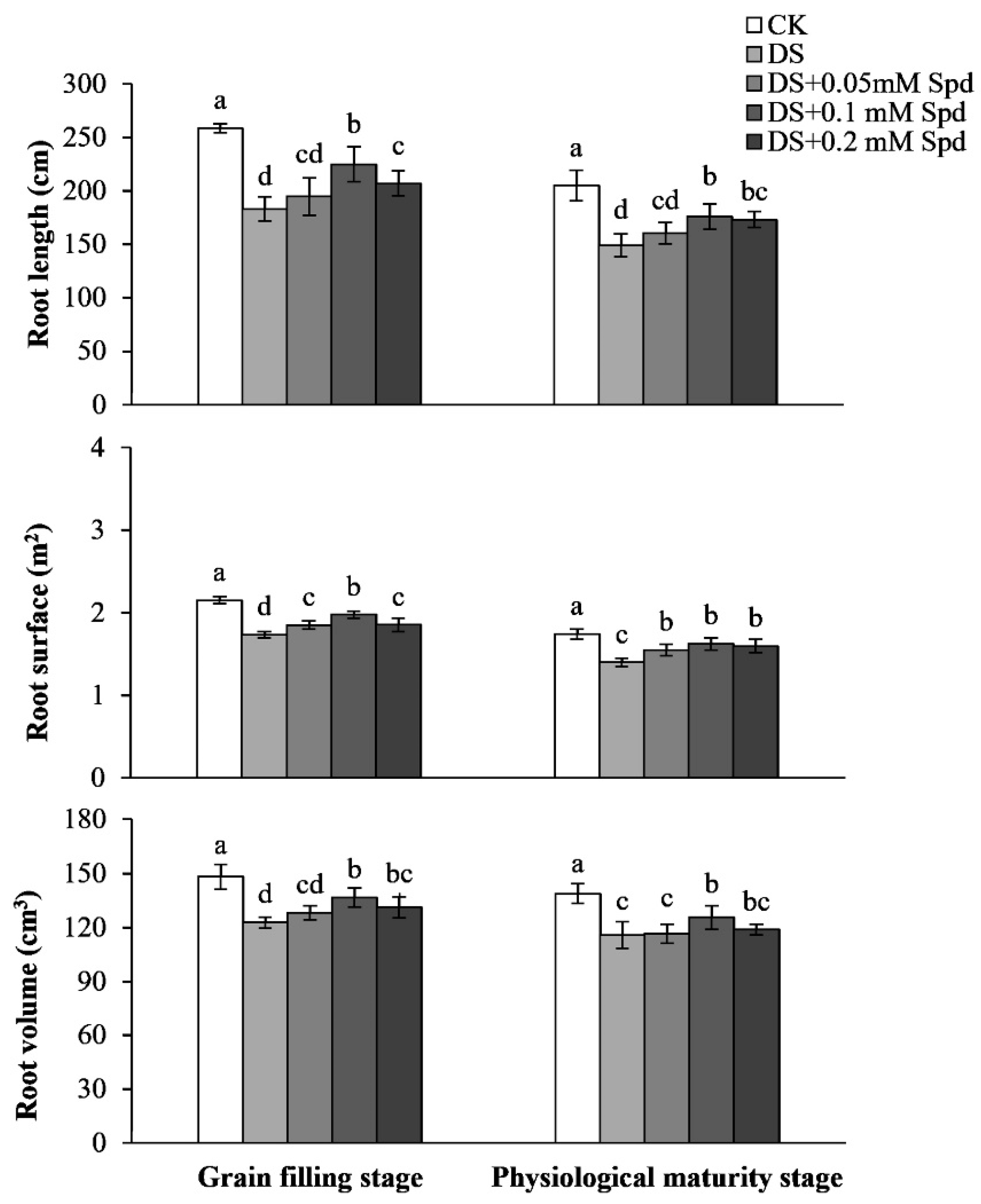
3.7. Root Characteristic Parameters
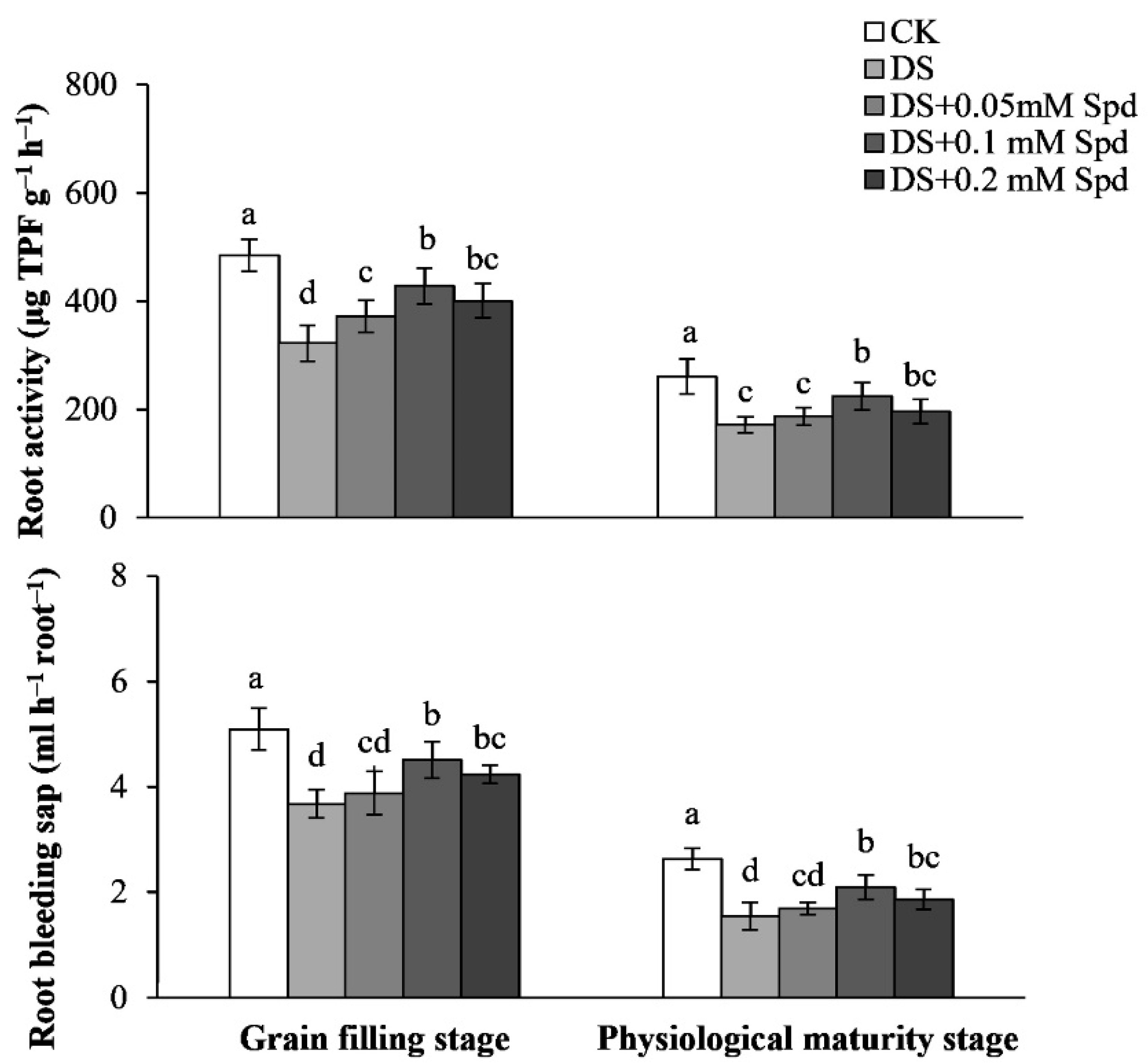
3.8. Root Activity and Root Bleeding Sap Rate
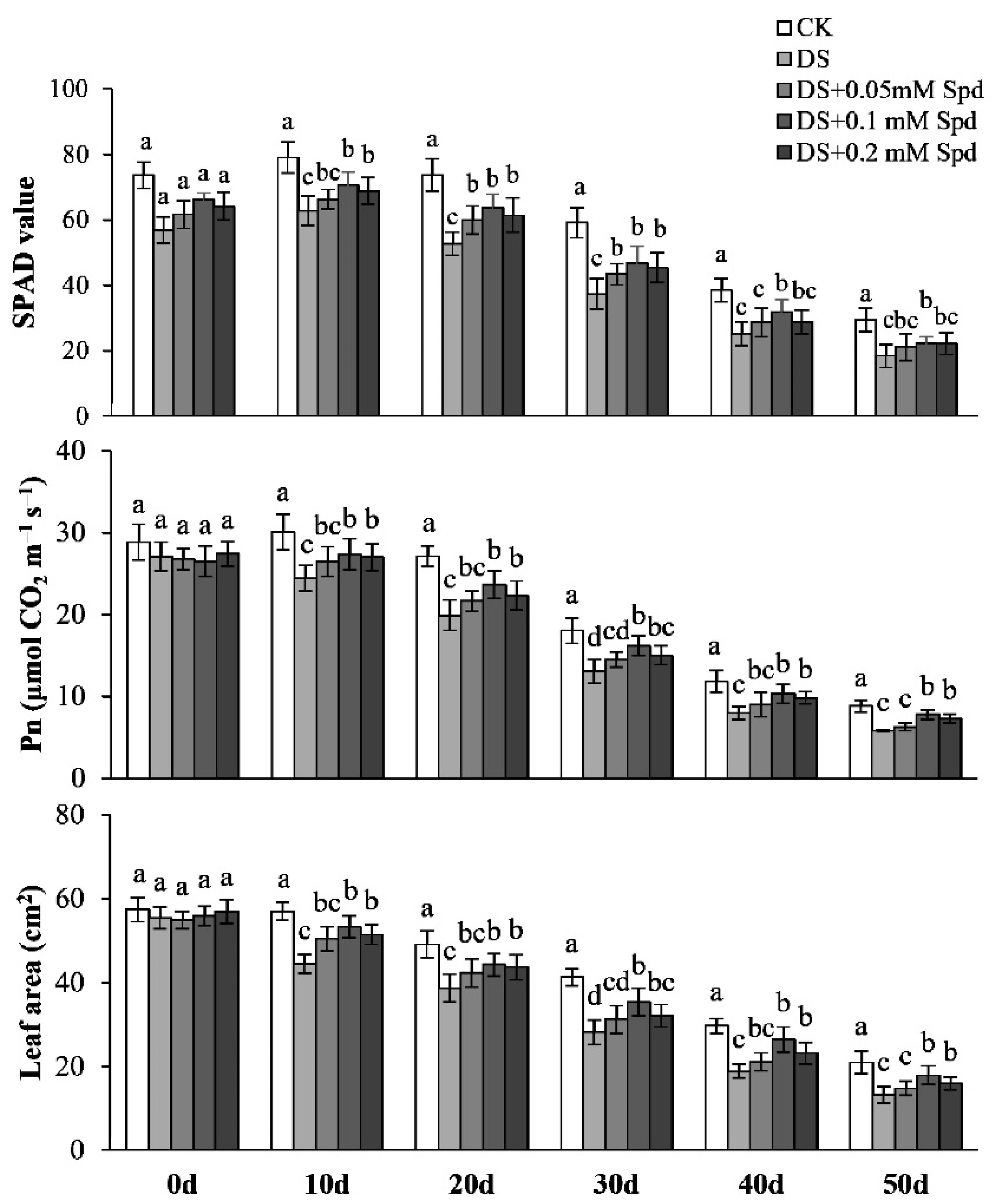
3.9. Leaf Photosynthesis and Leaf Area at Post-Anthesis Stage
3.10. Leaf Senescence Characteristics
3.11. Maize Yield and It’s Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, G.S. Research prospect on impact of climate change on agricultural production in China. Meteor. Environ. Sci. 2015, 38, 80–94. [Google Scholar] [CrossRef]
- Song, H.; Jiang, Y.L.; Xu, Z.Z.; Zhou, G.S. Response of photosynthetic physiological parameters of maize to drought during the whole growth period and after the jointing stage. Acta Ecol. Sin. 2019, 39, 2405–2415. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Dual application of 24-epibrassinolide and spermine confers drought stress tolerance in maize (Zea mays L.) by modulating polyamine and protein metabolism. J. Plant Growth Regul. 2016, 35, 518–533. [Google Scholar] [CrossRef]
- Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- O’Connell, E. Towards adaptation of water resource Systems to climatic and socio-economic Chang. Water Resour. Manag. 2017, 31, 2965–2984. [Google Scholar] [CrossRef]
- Meng, Q.; Hou, P.; Wu, L.; Chen, X.; Cui, Z.; Zhang, F. Understanding production potentials and yield gaps in intensive maize production in China. Field Crop. Res. 2013, 143, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Jiao, H.Y.; Zhou, G.S.; Chen, Z.L. Blue paper about agricultural issues on climate change. In Assessment Report on Effect of Climate Change on China’agriculture (No.1); Social Science Literature: Beijing, China, 2014; pp. 1–6. [Google Scholar]
- Avramova, V.; Abdelgawad, H.; Zhang, Z.; Fotschki, B.; Vergauwen, L.; Knapen, D.; Taleisnik, E.; Guisez, Y.; Asard, H. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol. 2015, 169, 1382–1396. [Google Scholar] [CrossRef]
- Bai, X.L.; Sun, S.X.; Yang, G.H.; Liu, M.; Zhang, Z.P.; Qi, H. Effect of water stress on maize yield during different growing stages. J. Maize Sci. 2009, 17, 60–63. [Google Scholar]
- Benari, T.; Adrian, J.; Klein, T.; Calanca, P.; Van der Velde, M.; Makowski, D. Identifying indicators for extreme wheat and maize yield losses. Agric. Forest Meteorol. 2016, 220, 130–140. [Google Scholar] [CrossRef]
- Lobell, D.B.; Roberts, M.J.; Wolfram, S.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Ribaut, J.M.; Betran, J.; Monneveux, P.; Setter, T. Handbook of Maize: Its Biology. In Drought Tolerance in Maize; Bennetzen, J.L., Hake, S.C., Eds.; Springer: New York, NY, USA, 2009; pp. 311–344. [Google Scholar]
- Jin, Z.N.; Zhuang, Q.L.; Wang, J.L.; Archontoulis, S.V.; Zobel, Z.; Kotamarthi, V.R. The combined and separate impacts of climate extremes on the current and future US rainfed maize and soybean production under elevated CO2. Glob. Chang. Biol. 2017, 23, 2687–2704. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.S.; Smith, M.R.; Guth, S.; Golden, G.D.; Vaitla, B.; Mueller, N.D.; Dangour, A.D.; Huybers, P. Climate change and global food systems: Potential impacts on food security and undernutrition. Annu. Rev. Public Health 2017, 38, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.; Nawaz, F.; Naeem, M.; Ashraf, M.Y. Effect of exogenous nitric oxide on sulfur and nitrate assimilation pathway enzymes in maize (Zea mays L.) under drought stress. Acta Physiol. Plantarum. 2018, 40, 206. [Google Scholar] [CrossRef]
- Sezgin, A.; Altuntas, C.; Demiralay, M.; Cinemre, S.; Terzi, S. Exogenous alpha lipoic acid can stimulate photosystem II activity and the gene expressions of carbon fixation and chlorophyll metabolism enzymes in maize seedlings under drought. J. Plant Physiol. 2019, 232, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Takaya, M. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [Green Version]
- Tavladoraki, P.; Cona, A.; Federico, R.; Tempera, G.; Viceconte, N.; Saccoccio, S.; Saccoccio, S.; Battaglia, V.; Toninello, A.; Agostinelli, E. Polyamine catabolism: Target for antiproliferative therapies in animals and stress tolerance strategies in plants. Amino Acids 2012, 42, 411–426. [Google Scholar] [CrossRef]
- Alcázar, R.; Bitrián, M.; Bartels, D.; Koncz, C.; Altabella, T.; Tiburcio, A.F. Polyamine metabolic canalization in response to drought stress in Arabidopsis and the resurrection plant Craterostigma plantagineum. Plant Signal. Behav. 2011, 6, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.X.; Zhang, W.H.; Liu, Y.L. Involvement of hydrogen peroxide generated by polyamine oxidative degradation in the development of lateral roots in soybean. J. Integr. Plant Biol. 2010, 48, 426–432. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Tebartz, F.; Scherer, G.F.E. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011, 181, 593–603. [Google Scholar] [CrossRef]
- Alet, A.I.; Sánchez, D.H.; Cuevas, J.C.; Marina, M.; Carrasco, P.; Altabella, T.; Tiburcio, A.F.; Ruiz, O.A. New insights into the role of spermine in Arabidopsis thaliana under long-term salt stress. Plant Sci. 2012, 182, 94–100. [Google Scholar] [CrossRef]
- Garwood, E.A.; Williams, T.B. Growth, water use and nutrient uptake from the subsoil by grass swards. J. Agric. Sci. 1967, 69, 125–130. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological responses of forest trees to heat and drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.L.; Li, M.J.; Zhou, K.; Sun, T.T.; Hu, L.Y.; Li, C.Y.; Ma, F.W. Uptake and metabolism of ammonium and nitrate in response to drought stress in malus prunifolia. Plant Physiol. Biochem. 2018, 127, 185–193. [Google Scholar] [CrossRef]
- Huang, L.; Li, M.J.; Shao, Y.; Sun, T.T.; Li, C.Y.; Ma, F.W. Ammonium uptake increases in response to PEG-induced drought stress in Malus hupehensis Rehd. Environ. Exp. Bot. 2018, 151, 32–42. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Oaks, A.; Long, D.M.; Zoumadakis, M.; Li, X.Z.; Hertig, C. The Role of NO3− and NH4+ Ions in the Regulation of Nitrate Reductase in Higher Plants; Springer-Verlag: Berlin/Heidelberg, Germany, 1990; pp. 165–170. [Google Scholar] [CrossRef]
- Xiong, D.L.; Yu, T.T.; Liu, X.; Li, Y.; Peng, S.B.; Huang, J.L. Heterogeneity of photosynthesis within leaves is associated with alteration of leaf structural features and leaf N content per leaf area in rice. Funct. Plant. Biol. 2015, 42, 687–696. [Google Scholar] [CrossRef]
- Wilkinson, S.; Bacon, M.A.; Davies, W.J. Nitrate signalling to stomata and growing leaves: Interactions with soil drying, ABA, and xylem sap pH in maize. J. Exp. Bot. 2007, 58, 1705–1716. [Google Scholar] [CrossRef]
- Bloom, A.J.; Sukrapanna, S.S.; Warner, R.L. Root respiration associated with ammonium and nitrate absorption and assimilation by barley. Plant Physiol. 1992, 99, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.P.; Zhang, Y.L.; Yao, H.S.; Zhang, X.J.; Luo, H.H.; Guo, L.; Zhang, W.F. Alternative electron sinks are crucial for conferring photoprotection in field-grown cotton under water deficit during flowering and boll setting stages. Funct. Plant Biol. 2014, 41, 737. [Google Scholar] [CrossRef]
- Cai, K.Z.; Wu, X.Z.; Luo, S.M. Effects of water stress on osmolytes at different growth stages in staces in rice leaves and roots. J. Plant Ecol. 2008, 32, 491–500. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Li, L.J.; Gu, W.R.; Li, J.; Li, C.F.; Xie, T.L.; Qu, D.Y.; Meng, Y.; Li, C.F.; Wei, S. Exogenously applied spermidine alleviates photosynthetic inhibition under drought stress in maize (Zea mays L.) seedlings associated with changes in endogenous polyamines and phytohormones. Plant Physiol. Biochem. 2018, 129, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.F.; Song, J.; Li, Y.R.; Yue, M.K.; Xiong, J.; Tao, L.X. Alterations of panicle antioxidant metabolism and carbohydrate content and pistil water potential involved in spikelet sterility in rice under water-deficit stress. Rice Sci. 2010, 17, 303–310. [Google Scholar] [CrossRef]
- Shi, H.T.; Chan, Z.L. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J. Integr. Plant Biol. 2014, 56, 114–121. [Google Scholar] [CrossRef]
- Hussain, S.; Farooq, M.; Wahid, M.; Ashfaq, A. Seed priming with putrescine improves the drought resistance of maize hybrids. Int. J. Agric. Biol. 2013, 15, 1560–8530. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.T.; Ye, T.T.; Chen, F.F.; Cheng, Z.M.; Wang, Y.P.; Yang, P.F.; Zhang, Y.S.; Chan, Z.L. Manipulation of arginase expression modulates abiotic stress tolerance in Arabidopsis: Effect on arginine metabolism and ROS accumulation. J. Exp. Bot. 2013, 8, 1367–1379. [Google Scholar] [CrossRef] [Green Version]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; RoubelakisAngelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine is involved in arabidopsis freezing tolerance and cold acclimation by regulating abscisic acid levels in response to low temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Pottosin, I.; Shabala, S. Polyamines control of cation transport across plant membranes: Implications for ion homeostasis and abiotic stress signaling. Front. Plant Sci. 2014, 5, 154. [Google Scholar] [CrossRef] [Green Version]
- Blatt, M.R. Cellular signaling and volume control in stomatal movements in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Fu, H.; Bei, Q.X.; Luan, S. Inward potassium channel in guard cells as a target for polyamine regulation of stomatal movements. Plant Physiol. 2000, 124, 1315–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Fu, X.Z.; Peng, T.; Huang, X.S.; Liu, J.H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Aharon, G.S.; Sottosanto, J.B.; Blumwald, E. Vacuolar Na+/H+ antiporter cation selectivity is regulated by calmodulin from within the vacuole in a Ca2+- and pH-dependent manner. Proc. Nati. Acad. Sci. USA 2005, 102, 16107–16112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.; Suzuki, N.; Ciftciyilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Toumi, I.; Moschou, P.N.; Paschalidis, K.A.; Bouamama, B.; Ben, S.A.; Ghorbel, A.W.; Mliki, A.; Roubelakis-Angelakis, K.A. Abscisic acid signals reorientation of polyamine metabolism to orchestrate stress responses via the polyamine exodus pathway in grapevine. J. Plant Physiol. 2010, 167, 519–525. [Google Scholar] [CrossRef]
- Wang, B.Q.; Zhang, Q.F.; Liu, J.H.; Li, G.H. Overexpression of PtADC confers enhanced dehydration and drought tolerance in transgenic tobacco and tomato: Effect on ROS elimination. Biochem. Biophys. Res. Commun. 2011, 413, 10–16. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Lee, I.J. Spermine promotes acclimation to osmotic stress by modifying antioxidant, abscisic acid, and jasmonic acid signals in soybean. J. Plant Growth Regul. 2013, 32, 22–30. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Li, Y.P.; Zhang, X.Q.; Ma, X.; Huang, L.K.; Yan, Y.H.; Peng, Y. Chitosan and spermine enhance drought resistance in white clover, associated with changes in endogenous phytohormones and polyamines, and antioxidant metabolism. Funct. Plant. Biol. 2018, 45, 1205–1222. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Romero, L.; Ruiz, J.M. Accumulation of free polyamines enhances the antioxidant response in fruits of grafted tomato plants under water stress. J. Plant Physiol. 2016, 190, 72–78. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, H.; Peng, Y.; Zhang, X.Q.; Xiao, Y.H. Exogenously applied spermidine improves drought tolerance in creeping bentgrass associated with changes in antioxidant defense, endogenous polyamines and phytohormones. Plant Growth Regul. 2015, 76, 71–82. [Google Scholar] [CrossRef]
- Espasandin, F.D.; Maiale, S.J.; Calzadilla, P.; Ruiz, O.A.; Sansberro, R.A. Transcriptional regulation of 9-cis-epoxycarotenoid dioxygenase (NCED) gene by putrescine accumulation positively modulates ABA synthesis and drought tolerance in Lotus tenuis plants. Plant Physiol. Biochem. 2014, 76, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.X.; Li, Q.; Dou, P.; Kong, F.L.; Ma, X.J.; Cheng, Q.B.; Yuan, J.C. Effect of low nitrogen stress on bleeding sap characters and root activity of maize cultivars with different low N tolerance. J. Plant Nutr. Fertil. 2017, 23, 278–288. [Google Scholar] [CrossRef]
- Jia, Q.M.; Yang, L.Y.; An, H.Y.; Dong, S.; Chang, S.H.; Zhang, C.; Liu, Y.J.; Hou, F.J. Nitrogen fertilization and planting models regulate maize productivity, nitrate and root distributions in semi-arid regions. Soil Tillage Res. 2020, 200, 104636. [Google Scholar] [CrossRef]
- Natali, S.M.; Sañudo-Wilhelmy, S.A.; Lerdau, M.T. Effects of elevated carbon dioxide and nitrogen fertilization on nitrate reductase activity in sweetgum and loblolly pine trees in two temperate forests. Plant Soil. 2009, 314, 197–210. [Google Scholar] [CrossRef]
- Yu, X.Z.; Zhang, F.Z. Activities of nitrate reductase and glutamine synthetase in rice seedlings during cyanide metabolism. J. Hazard. Mater. 2012, 225–226, 190–194. [Google Scholar] [CrossRef]
- Singh, R.P.; Srivastava, H.S. Increase in glutamate synthase (GOGAT) activity in maize seedlings in response to nitrate and ammonium nitrogen. Physiol. Plant. 1986, 66, 413–416. [Google Scholar] [CrossRef]
- Magalhaes, J.R.; Huber, D.M. Response of ammonium assimilation enzymes to nitrogen form treatments in different plant species. J. Plant Nutr. 1991, 14, 175–185. [Google Scholar] [CrossRef]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y.F. Nitrate signaling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, O. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.Q.; Young, J.; Crawford, N.M. The nitrate transporter AtNRT1.1 (CHL1) functions in stomatal opening and contributes to drought susceptibility in Arabidopsis. Plant Cell. 2003, 15, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Valadier, M.H.; Migge, A.; Becker, T.W. Drought-induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiol. 1998, 117, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaut, Z. Nitrate Reductase activity of wheat seedlings during exposure to and recovery from water stress and salinity. Physiol. Plantarum. 2010, 30, 212–217. [Google Scholar] [CrossRef]
- Miura, K. Nitrogen and phosphorus nutrition under salinity stress. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M., Prasad, M., Eds.; Springer: New York, NY, USA, 2013; pp. 425–441. [Google Scholar]
- Sunil, B.; Talla, S.K.; Aswani, V.; Raghavendra, A.S. Optimization of photosynthesis by multiple metabolic pathways involving interorganelle interactions: Resource sharing and ROS maintenance as the bases. Photosynth. Res. 2013, 117, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Dugo, V.; Durand, J.L.; Gastal, F. Water Deficit and Nitrogen Nutrition of Crops. In Sustainable Agriculture; Lichtfouse, E., Hamelin, M., Navarrete, M., Debaeke, P., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 2, pp. 557–575. [Google Scholar]
- Hoai, N.T.T.; Shim, I.S.; Kobayashi, K.; Kenji, U. Accumulation of some nitrogen compounds in response to salt stress and their relationships with salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2003, 41, 159–164. [Google Scholar] [CrossRef]
- Márquez, A.J.; Betti, M.; García-Calderón, M.; Estivill, G. Nitrate and ammonium assimilatory enzymes. In Lotus Japonicus Handbook; Márquez, A.J., Ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 315–328. [Google Scholar]
- Du, J.; Shu, S.; Shao, Q.S.; An, Y.H.; Zhou, H.; Guo, S.R.; Sun, J. Mitigative effects of spermidine on photosynthesis and carbon–nitrogen balance of cucumber seedlings under Ca(NO3)2 stress. J. Plant Res. 2016, 129, 79–91. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef]
- Labboun, S.; Terce-Laforgue, T.; Roscher, A.; Bedu, M.; Restivo, F.M.; Velanis, C.N.; Skopelitis, D.S.; Moschou, P.N.; Roubelakis-Angelakis, K.A.; Suzuki, A.; et al. Resolving the role of plant glutamate dehydrogenase. I. in vivo real time nuclear magnetic resonance spectroscopy experiments. Plant Cell Physiol. 2009, 50, 1761–1773. [Google Scholar] [CrossRef] [Green Version]
- King, J.; Waygood, E.R. Glyoxylate aminotransferases from wheat leaves. Can. J. Biochem. 1968, 46, 771–779. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.H.; Feraud, M.; Jouglet, T.; Suzuki, A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar] [CrossRef] [Green Version]
- Joshi, V.; Joung, J.G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, D.H.; Al-Kaisi, M.M.; Zhang, Y.S.; Duan, L.S.; Tan, W.M.; Zhang, M.C.; Li, Z.H. Tillage practices affect biomass and grain yield through regulating root growth, root-bleeding sap and nutrients uptake in summer maize. Field Crop. Res. 2014, 157, 89–97. [Google Scholar] [CrossRef]
- Fageria, N.K. Influence of dry matter and length of roots on growth of five field crops at varying soil zinc and copper levels. J. Plant Nutr. 2004, 27, 1517–1523. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, A.W.; Qiu, X.Q.; Liu, Z.G.; Sun, J.S.; Zhang, J.P.; Wang, H.Z. Distribution of roots and root length density in a maize/soybean strip intercropping system. Agr. Water Manage. 2010, 98, 199–212. [Google Scholar] [CrossRef]
- Wasson, A.P.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Richards, R.A.; Watt, M. Soil coring at multiple field environments can directly quantify variation in deep root traits to select wheat genotypes for breeding. J. Exp. Bot. 2014, 65, 6231–6249. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Zhu, D.; Lin, X.; Han, W. Effects of polyamines on tobacco root growth and development. Plant Physlol. Commun. 1992, 28, 124–126. [Google Scholar] [CrossRef]
- Peuke, A.D. The chemical composition of xylem sap in Vitis vinifera L. cv. Riesling during vegetative growth on three different Franconian vineyard soils and as influenced by nitrogen fertilizer. Am. J. Enol. Vitic. 2000, 51, 329–339. [Google Scholar] [CrossRef]
- Amos, B.; Walters, D.T. Maize root biomass and net rhizodeposited carbon. Soil Sci. Soc. Am. J. 2006, 70, 1489–1503. [Google Scholar] [CrossRef]
- Morita, S.; Okamoto, M.; Abe, J.; Yamagishi, J. Bleeding rate of field-grown maize with reference to root system development. Jpn. J. Crop Sci. 2000, 69, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, A.; Kageyama, M.; Shinmachi, F.; Schmidhalter, U.; Hasegawa, I. Potential for using plant xylem sap to evaluate inorganic nutrient availability in soil. Soil Sci. Plant Nutr. 2005, 51, 343–350. [Google Scholar] [CrossRef]
- Ansari, T.H.; Yamamoto, Y.; Yoshida, T.; Sakagami, K. Relation between bleeding rate during panicle formation stage and sink size in rice plant. Soil Sci. Plant Nutr. 2004, 50, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Ji, B.; Li, J.; Xin, L.; Wang, F.; Li, B. Effects of spermidine on antioxidant properties and root activity of rice stubble wheat seedlings under waterlogging stress. J. Henan Agric. Sci. 2018, 47, 29–33. [Google Scholar] [CrossRef]
- Seng, S.S.; Wang, Q.; Zhang, Y.E.; Li, C.H.; Liu, T.X.; Zhao, L.F.; Liu, H.P. Effects of exogenous spermidine on physiological regulatory of maize after waterlogging stress. Acta Agron. Sin. 2012, 38, 1042–1050. [Google Scholar] [CrossRef]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Liu, M.Z.; Gao, L.M. Photosynthetic rate and yield formation in different maize hybrids. Biol. Plant 2007, 51, 165–168. [Google Scholar] [CrossRef]
- Marchiori, P.E.R.; Machado, E.C.; Ribeiro, R.V. Photosynthetic limitations imposed by self-shading in field-grown sugarcane varieties. Field Crop. Res. 2014, 155, 30–37. [Google Scholar] [CrossRef]
- Chen, X.C.; Chen, F.J.; Chen, Y.L.; Gao, Q.; Yang, X.L.; Yuan, L.X.; Zhang, F.S.; Mi, G.H. Modern maize hybrids in Northeast China exhibit increased yield potential and resource use efficiency despite adverse climate change. Glob. Chang Biol. 2013, 19, 923–936. [Google Scholar] [CrossRef]
- Sun, X.; Ding, Z.; Wang, X.; Ming, Z. Subsoiling practices change root distribution and increase post-anthesis dry matter accumulation and yield in summer maize. PLoS ONE 2017, 12, e174952. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.F.; Ding, Z.S.; Hou, H.P.; Ge, J.Z.; Tang, L.; Zhao, M. Post-anthesis photosynthetic assimilation and the changes of carbon and nitrogen in different varieties of spring maize. Acta Agron. Sin. 2013, 39, 1284–1292. [Google Scholar] [CrossRef]
- Martínez, D.; Guiamet, J. Distortion of the SPAD 502 chlorophyll meter readings by changes in irradiance and leaf water status. Agronomie 2004, 24, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Duvick, D.N. The contribution of breeding to yield advances in maize (Zea mays L.). Adv. Agron. 2005, 86, 83–145. [Google Scholar] [CrossRef]
- Zhou, B.Y.; Yue, Y.; Sun, X.F.; Wang, X.B.; Wang, Z.M.; Ma, W.; Zhao, M. Maize grain yield and dry matter production responses to variations in weather conditions. Agron. J. 2015, 108, 196–204. [Google Scholar] [CrossRef]
- Khan, N.A.; Murayama, S.; Ishimine, Y.; Tsuzuki, E.; Nakamura, I. Physio-morphological studies of f1 hybrids in rice (Oryza sativa L.): Photosynthetic ability and yield. Plant Prod. Sci. 1998, 1, 233–239. [Google Scholar] [CrossRef]
- Schussler, J.R.; Westgate, M.E. Assimilate flux determines kernel set at low water potential in maize. Crop Sci. 1995, 35, 1074–1080. [Google Scholar] [CrossRef]
- Zhang, J.P.; Sun, J.S.; Liu, Z.G.; Gao, Y. Effect of moisture and mulching on filling characteristics and yield of summer maize. Chin. J. Eco.-Agric. 2010, 18, 501–506. [Google Scholar] [CrossRef]
- Bräutigam, A.; Gagneul, D.; Weber, A.P.M. High-throughput colorimetric method for the parallel assay of glyoxylic acid and ammonium in a single extract. Anal. Biochem. 2007, 362, 151–153. [Google Scholar] [CrossRef] [PubMed]




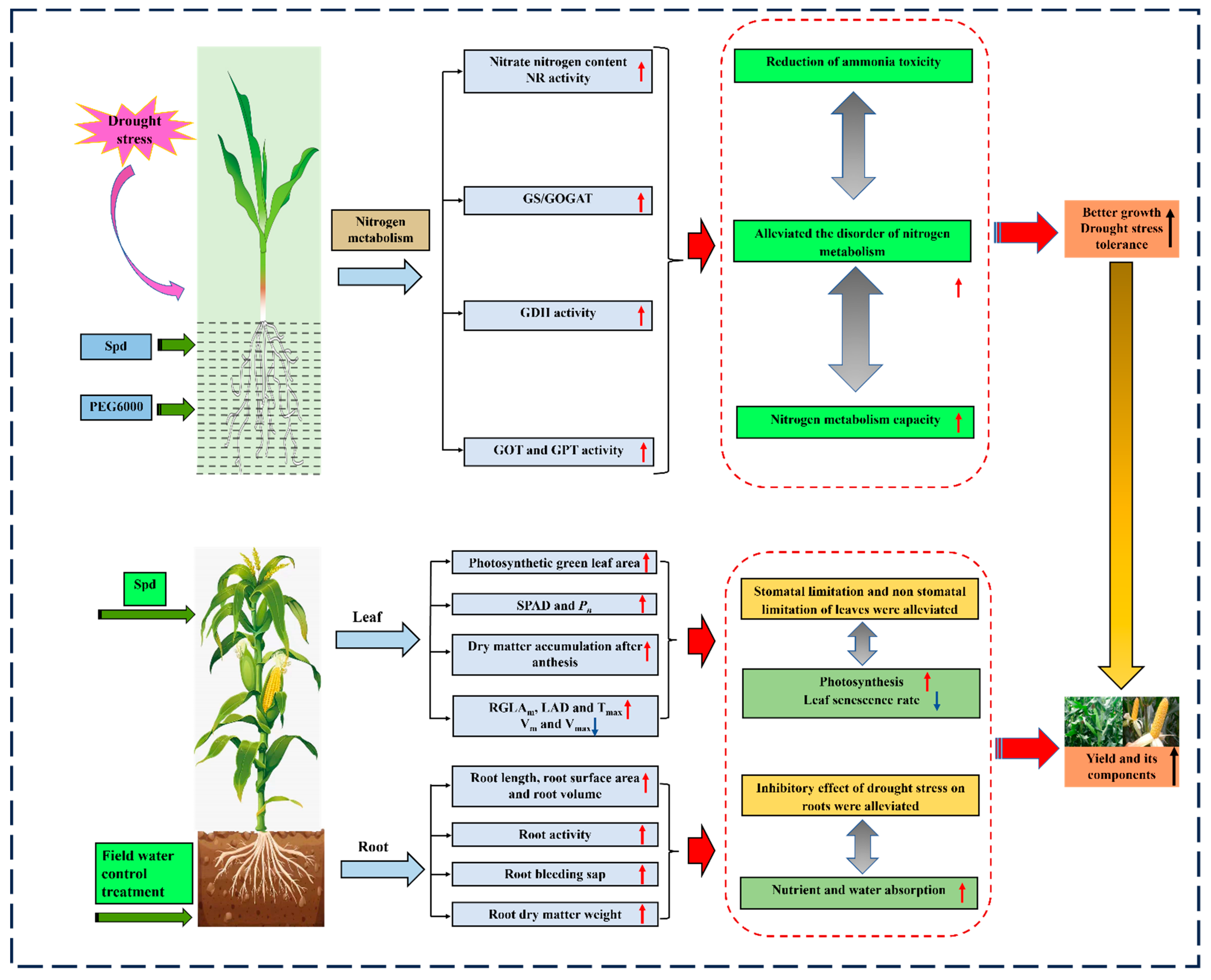
 ) and the blue arrows (
) and the blue arrows (  ) represent the positive and passive roles of spermidine, respectively.
) and the blue arrows ( ) represent the positive and passive roles of spermidine, respectively.
) represent the positive and passive roles of spermidine, respectively.
) and the blue arrows ( ) represent the positive and passive roles of spermidine, respectively.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Xianyu 335 | Fenghe 1 | ||
|---|---|---|---|---|
| 2d | 4d | 2d | 4d | |
| CK | 28.83 ± 2.13 ab | 27.08 ± 1.63 a | 30.00 ± 2.21 a | 31.13 ± 2.21 a |
| Spd | 31.31 ± 1.49 a | 26.88 ± 1.73 a | 32.27 ± 1.76 a | 31.43 ± 2.73 a |
| PEG | 23.69 ± 2.14 c | 15.71 ± 1.28 c | 17.23 ± 2.37 c | 10.93 ± 1.56 c |
| Spd + PEG | 26.16 ± 1.56 bc | 20.13 ± 1.24 b | 22.13 ± 1.96 b | 15.97 ± 1.98 b |
| Treatment | Varieties | Parameters | 0d | 1d | 2d | 3d | 4d |
|---|---|---|---|---|---|---|---|
| CK | Xianyu 335 | NO3− contents (mg g−1 FW) | 3.60 ± 0.27 a | 3.18 ± 0.23 ab | 3.23 ± 0.18 a | 3.39 ± 0.13 a | 3.21 ± 0.22 a |
| Spd | 3.91 ± 0.19 a | 3.36 ± 0.22 a | 3.06 ± 0.26 ab | 3.13 ± 0.25 a | 3.33 ± 0.24 a | ||
| PEG | 3.67 ± 0.22 a | 2.46 ± 0.28 c | 1.96 ± 0.25 c | 1.67 ± 0.10 c | 1.89 ± 0.10 c | ||
| Spd + PEG | 3.94 ± 0.17 a | 2.93 ± 0.15 b | 2.70 ± 0.18 b | 2.35 ± 0.20 b | 2.47 ± 0.20 b | ||
| CK | Fenghe 1 | NO3− contents (mg g−1 FW) | 3.00 ± 0.22 a | 3.11 ± 0.22 a | 2.58 ± 0.22 a | 2.71 ± 0.16 a | 2.96 ± 0.19 a |
| Spd | 3.23 ± 0.18 a | 3.14 ± 0.27 a | 2.72 ± 0.25 a | 2.93 ± 0.15 a | 2.70 ± 0.18 a | ||
| PEG | 3.06 ± 0.26 a | 1.73 ± 0.22 c | 1.52 ± 0.16 c | 1.31 ± 0.11 c | 0.90 ± 0.13 c | ||
| Spd + PEG | 3.28 ± 0.14 a | 2.60 ± 0.20 b | 2.22 ± 0.30 b | 2.70 ± 0.18 b | 1.64 ± 0.15 b | ||
| CK | Xianyu 335 | NH4+ contents (μg g−1 FW) | 294.24 ± 21.47 a | 247.25 ± 20.88 b | 261.93 ± 26.51 b | 293.76 ± 26.22 c | 281.52 ± 27.45 c |
| Spd | 316.91 ± 17.07 a | 267.65 ± 29.34 ab | 272.14 ± 26.96 b | 278.46 ± 21.95 c | 247.86 ± 17.03 c | ||
| PEG | 299.47 ± 25.48 a | 303.14 ± 27.38 a | 350.88 ± 29.89 a | 397.80 ± 15.03 a | 412.42 ± 15.04 a | ||
| Spd + PEG | 321.51 ± 13.76 a | 283.15 ± 30.24 ab | 307.36 ± 20.94 ab | 340.00 ± 26.92 b | 345.44 ± 22.95 b | ||
| CK | Fenghe 1 | NH4+ contents (μg g−1 FW) | 246.68 ± 16.08 a | 218.82 ± 21.08 c | 197.60 ± 16.21 c | 191.90 ± 16.21 c | 203.30 ± 20.57 c |
| Spd | 215.33 ± 15.79 a | 192.53 ± 12.97 c | 187.47 ± 20.65 c | 207.73 ± 22.77 c | 211.22 ± 20.93 c | ||
| PEG | 243.11 ± 25.92 a | 317.62 ± 13.35 a | 363.22 ± 22.98 a | 428.45 ± 26.48 a | 472.47 ± 22.93 a | ||
| Spd + PEG | 228.00 ± 21.52 a | 264.73 ± 14.91 b | 307.80 ± 20.11 b | 340.10 ± 23.98 b | 359.10 ± 20.55 b |
| Treatment | Varieties | NR Activity (μg g−1 FW h−1) | ||||
|---|---|---|---|---|---|---|
| 0d | 1d | 2d | 3d | 4d | ||
| CK | Xianyu 335 | 12.50 ± 0.91 a | 14.03 ± 1.07 a | 13.19 ± 1.15 a | 13.52 ± 0.89 a | 14.92 ± 1.11 a |
| Spd | 13.47 ± 0.74 a | 14.86 ± 0.80 a | 14.27 ± 0.99 a | 13.80 ± 1.25 a | 14.09 ± 0.62 a | |
| PEG | 12.72 ± 1.09 a | 11.63 ± 1.08 b | 8.00 ± 0.84 c | 6.02 ± 0.61 c | 9.37 ± 0.45 c | |
| Spd + PEG | 13.68 ± 0.59 a | 13.04 ± 1.15 ab | 10.41 ± 1.02 b | 8.56 ± 0.60 b | 12.09 ± 1.13 b | |
| CK | Fenghe 1 | 10.40 ± 0.69 a | 11.48 ± 0.86 a | 9.88 ± 0.89 a | 9.61 ± 0.70 a | 10.80 ± 0.82 a |
| Spd | 10.62 ± 0.96 a | 10.84 ± 0.46 a | 11.11 ± 0.78 a | 10.37 ± 0.58 a | 11.44 ± 1.08 a | |
| PEG | 9.65 ± 0.48 a | 7.23 ± 0.37 c | 4.69 ± 0.52 c | 3.46 ± 0.31 c | 3.95 ± 0.84 c | |
| Spd + PEG | 10.42 ± 0.66 a | 9.30 ± 0.87 b | 7.63 ± 0.69 b | 5.36 ± 0.49 b | 5.70 ± 0.69 b | |
| Treatment | Varieties | Parameters | 0d | 1d | 2d | 3d | 4d |
|---|---|---|---|---|---|---|---|
| CK | Xianyu 335 | GS activity (μmol GHA g−1 FW h−1) | 26.64 ± 1.75 a | 28.79 ± 1.54 a | 29.55 ± 2.33 a | 26.50 ± 2.18 a | 25.89 ± 1.78 a |
| Spd | 24.82 ± 1.76 a | 30.66 ± 2.27 a | 30.75 ± 2.24 a | 25.31 ± 2.23 a | 26.77 ± 2.08 a | ||
| PEG | 24.33 ± 2.17 a | 25.04 ± 2.00 b | 20.30 ± 1.59 c | 16.29 ± 1.15 c | 15.20 ± 1.30 c | ||
| Spd + PEG | 27.48 ± 1.49 a | 29.16 ± 1.91 a | 24.93 ± 1.80 b | 20.76 ± 1.87 b | 21.59 ± 1.18 b | ||
| CK | Fenghe 1 | GS activity (μmol GHA g−1 FW h−1) | 22.50 ± 1.20 a | 24.23 ± 1.13 a | 23.11 ± 1.90 a | 25.44 ± 2.41 a | 22.72 ± 2.03 a |
| Spd | 22.65 ± 1.09 a | 25.11 ± 2.19 a | 25.40 ± 1.92 a | 23.66 ± 1.13 a | 23.42 ± 2.13 a | ||
| PEG | 20.79 ± 1.84 a | 15.53 ± 1.82 c | 11.26 ± 1.19 c | 10.38 ± 1.55 c | 7.64 ± 0.93 c | ||
| Spd + PEG | 22.43 ± 1.02 a | 20.29 ± 1.42 b | 16.59 ± 1.37 b | 15.83 ± 2.36 b | 12.93 ± 1.63 b | ||
| CK | Xianyu 335 | GOGAT activity (μmol NADH g−1 FW h−1) | 0.65 ± 0.05 a | 0.58 ± 0.04 a | 0.59 ± 0.05 ab | 0.52 ± 0.05 a | 0.56 ± 0.03 a |
| Spd | 0.71 ± 0.06 a | 0.64 ± 0.04 a | 0.63 ± 0.04 a | 0.56 ± 0.03 a | 0.59 ± 0.05 a | ||
| PEG | 0.62 ± 0.06 a | 0.45 ± 0.03 b | 0.41 ± 0.03 c | 0.28 ± 0.02 c | 0.36 ± 0.03 c | ||
| Spd + PEG | 0.68 ± 0.05 a | 0.59 ± 0.04 a | 0.54 ± 0.04 b | 0.36 ± 0.03 b | 0.46 ± 0.05 b | ||
| CK | Fenghe 1 | GOGAT activity (μmol NADH g−1 FW h−1) | 0.52 ± 0.03 a | 0.59 ± 0.05 a | 0.57 ± 0.04 a | 0.58 ± 0.02 a | 0.57 ± 0.03 a |
| Spd | 0.50 ± 0.05 a | 0.57 ± 0.03 a | 0.58 ± 0.03 a | 0.60 ± 0.05 a | 0.61 ± 0.02 a | ||
| PEG | 0.56 ± 0.04 a | 0.37 ± 0.02 c | 0.30 ± 0.03 c | 0.22 ± 0.03 c | 0.25 ± 0.02 c | ||
| Spd + PEG | 0.54 ± 0.04 a | 0.47 ± 0.03 b | 0.45 ± 0.02 b | 0.35 ± 0.03 b | 0.37 ± 0.04 b | ||
| CK | Xianyu 335 | GDH activity (μmol NAD g−1 FW h−1) | 1.19 ± 0.07 a | 1.22 ± 0.08 b | 1.09 ± 0.09 b | 1.22 ± 0.08 c | 1.12 ± 0.09 c |
| Spd | 1.31 ± 0.10 a | 1.31 ± 0.07 ab | 1.18 ± 0.06 ab | 1.26 ± 0.07 bc | 1.18 ± 0.05 c | ||
| PEG | 1.25 ± 0.10 a | 1.37 ± 0.06 a | 1.29 ± 0.07 a | 1.39 ± 0.06 b | 1.42 ± 0.06 b | ||
| Spd + PEG | 1.16 ± 0.10 a | 1.44 ± 0.09 a | 1.31 ± 0.07 a | 1.54 ± 0.07 a | 1.61 ± 0.07 a | ||
| CK | Fenghe 1 | GDH activity (μmol NAD g−1 FW h−1) | 1.06 ± 0.06 a | 0.97 ± 0.08 a | 1.08 ± 0.07 b | 1.08 ± 0.07 a | 1.07 ± 0.07 a |
| Spd | 1.00 ± 0.07 a | 1.00 ± 0.06 a | 1.09 ± 0.07 b | 1.18 ± 0.08 a | 1.13 ± 0.04 a | ||
| PEG | 0.93 ± 0.06 a | 0.77 ± 0.03 b | 0.66 ± 0.06 c | 0.48 ± 0.05 c | 0.37 ± 0.04 c | ||
| Spd + PEG | 1.06 ± 0.07 a | 0.89 ± 0.07 a | 0.85 ± 0.07 a | 0.78 ± 0.07 b | 0.69 ± 0.07 b |
| Treatment | Varieties | Parameters | 0d | 1d | 2d | 3d | 4d |
|---|---|---|---|---|---|---|---|
| CK | Xianyu 335 | GOT activity (μmol mg−1 30 min−1) | 0.27 ± 0.02 a | 0.26 ± 0.02 a | 0.23 ± 0.02 a | 0.26 ± 0.02 a | 0.27 ± 0.01 a |
| Spd | 0.27 ± 0.02 a | 0.28 ± 0.02 a | 0.25 ± 0.02 a | 0.28 ± 0.02 a | 0.29 ± 0.02 a | ||
| PEG | 0.29 ± 0.02 a | 0.22 ± 0.01 b | 0.18 ± 0.01 b | 0.17 ± 0.01 c | 0.15 ± 0.02 c | ||
| Spd + PEG | 0.27 ± 0.03 a | 0.24 ± 0.01 ab | 0.22 ± 0.02 a | 0.21 ± 0.02 b | 0.21 ± 0.01 b | ||
| CK | Fenghe 1 | GOT activity (μmol mg−1 30 min−1) | 0.26 ± 0.02 a | 0.28 ± 0.03 a | 0.29 ± 0.02 a | 0.26 ± 0.02 a | 0.24 ± 0.02 a |
| Spd | 0.27 ± 0.03 a | 0.29 ± 0.03 a | 0.31 ± 0.03 a | 0.26 ± 0.03 a | 0.27 ± 0.03 a | ||
| PEG | 0.29 ± 0.03 a | 0.20 ± 0.02 c | 0.16 ± 0.02 c | 0.14 ± 0.02 c | 0.10 ± 0.01 c | ||
| Spd + PEG | 0.26 ± 0.02 a | 0.25 ± 0.02 b | 0.22 ± 0.03 b | 0.20 ± 0.02 b | 0.17 ± 0.02 b | ||
| CK | Xianyu 335 | GPT activity (μmol mg−1 30 min−1) | 0.37 ± 0.04 a | 0.31 ± 0.03 ab | 0.36 ± 0.03 a | 0.31 ± 0.04 a | 0.30 ± 0.03 a |
| Spd | 0.39 ± 0.03 a | 0.34 ± 0.03 a | 0.34 ± 0.03 a | 0.30 ± 0.02 a | 0.31 ± 0.03 a | ||
| PEG | 0.36 ± 0.02 a | 0.25 ± 0.04 b | 0.23 ± 0.03 b | 0.21 ± 0.02 c | 0.18 ± 0.02 c | ||
| Spd + PEG | 0.40 ± 0.03 a | 0.31 ± 0.03 ab | 0.29 ± 0.03 a | 0.25 ± 0.03 b | 0.23 ± 0.03 b | ||
| CK | Fenghe 1 | GPT activity (μmol mg−1 30 min−1) | 0.30 ± 0.02 a | 0.31 ± 0.02 a | 0.33 ± 0.03 a | 0.37 ± 0.04 a | 0.33 ± 0.03 a |
| Spd | 0.34 ± 0.03 a | 0.33 ± 0.02 a | 0.33 ± 0.02 a | 0.35 ± 0.04 a | 0.36 ± 0.03 a | ||
| PEG | 0.35 ± 0.04 a | 0.26 ± 0.02 b | 0.20 ± 0.01 c | 0.16 ± 0.02 c | 0.14 ± 0.03 c | ||
| Spd + PEG | 0.33 ± 0.03 a | 0.29 ± 0.03 ab | 0.25 ± 0.03 b | 0.22 ± 0.03 b | 0.20 ± 0.02 b |
| Treatment | RGLAm (%) | Vm (%) | Vmax (%) | Tmax (d) | LAD (m2 d−1) |
|---|---|---|---|---|---|
| CK | 43.05 | 1.10 | 1.87 | 45.91 | 21.70 |
| DS | 18.30 | 1.67 | 2.95 | 32.04 | 14.74 |
| DS + 0.05 mM Spd | 22.67 | 1.41 | 2.41 | 37.52 | 17.96 |
| DS + 0.1 mM Spd | 28.04 | 1.36 | 2.34 | 38.83 | 17.71 |
| DS + 0.2 mM Spd | 25.08 | 1.50 | 2.60 | 34.70 | 15.69 |
| Treatment | Kernels (No Ear−1) | 100-Kernel Weight (g) | Ear Number (No m−2) | Yield (kg ha−1) |
|---|---|---|---|---|
| CK | 486.69 ± 10.85 a | 33.72 ± 0.49 a | 7.2 ± 0.5 a | 11,816.81 ± 200.56 a |
| DS | 444.71 ± 11.43 c | 29.07 ± 1.06 c | 6.8 ± 0.2 a | 8790.63 ± 124.58 c |
| DS + 0.05 mM Spd | 452.21 ± 14.56 bc | 30.23 ± 0.75 bc | 6.8 ± 0.4 a | 9295.23 ± 178.43 bc |
| DS + 0.1 mM Spd | 461.47 ± 9.17 b | 31.09 ± 1.04 b | 7.0 ± 0.2 a | 9841.26 ± 126.60 b |
| DS + 0.2 mM Spd | 457.49 ± 6.79 bc | 30.62 ± 0.85 bc | 6.8 ± 0.4 a | 9525.89 ± 132.24 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Li, L.; Meng, Y.; Liu, H.; Li, J.; Yu, Y.; Qian, C.; Wei, S.; Gu, W. Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions. Agriculture 2022, 12, 1270. https://doi.org/10.3390/agriculture12081270
Dong L, Li L, Meng Y, Liu H, Li J, Yu Y, Qian C, Wei S, Gu W. Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions. Agriculture. 2022; 12(8):1270. https://doi.org/10.3390/agriculture12081270
Chicago/Turabian StyleDong, Ling, Lijie Li, Yao Meng, Hongliang Liu, Jing Li, Yang Yu, Chunrong Qian, Shi Wei, and Wanrong Gu. 2022. "Exogenous Spermidine Optimizes Nitrogen Metabolism and Improves Maize Yield under Drought Stress Conditions" Agriculture 12, no. 8: 1270. https://doi.org/10.3390/agriculture12081270