
RNAi Suppression of Hormone Receptor HR3 Blocks Larval Molting and Metamorphosis in the Cigarette Beetle, Lasioderma serricorne

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects Culture
2.2. Molecular Cloning and Sequence Analysis
2.3. Spatiotemporal Expression Analysis
2.4. Expression Profiles of LsHR3 in Response to 20E
2.5. Functional Analysis of LsHR3 by RNAi
2.6. Knockdown of LsHR3 on Chitin Metabolism and Cuticle Formation
2.7. Statistical Analysis
3. Results
3.1. Identification and Characterization of LsHR3
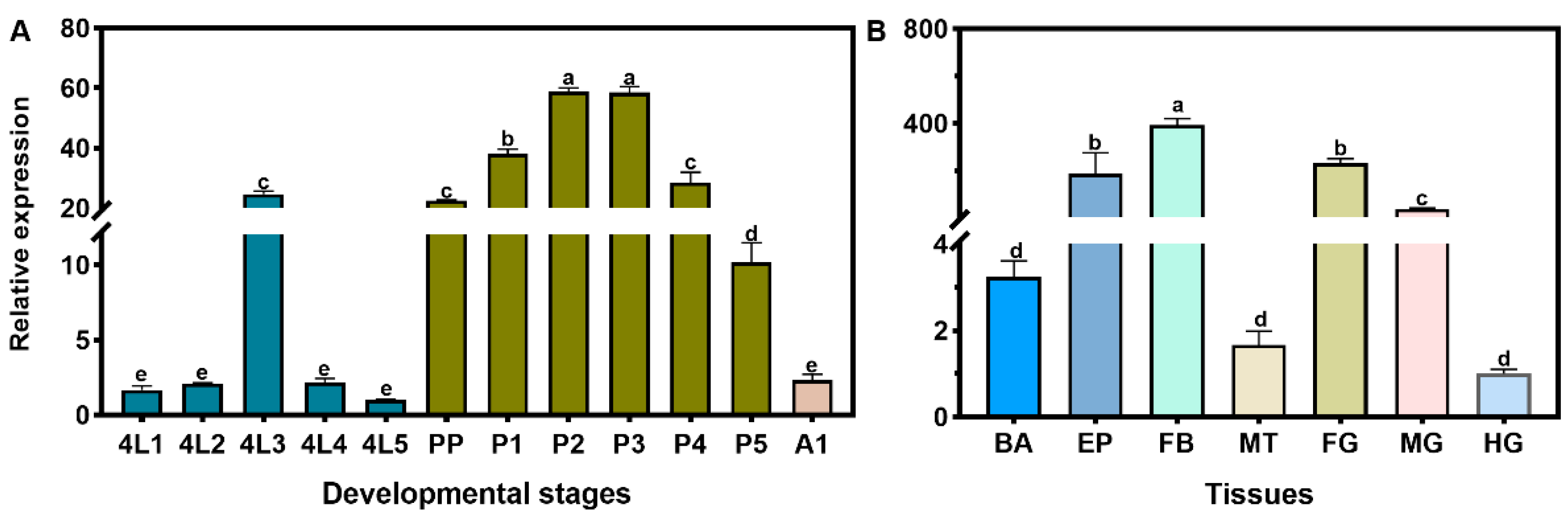
3.2. Spatiotemporal Expression Analysis of LsHR3
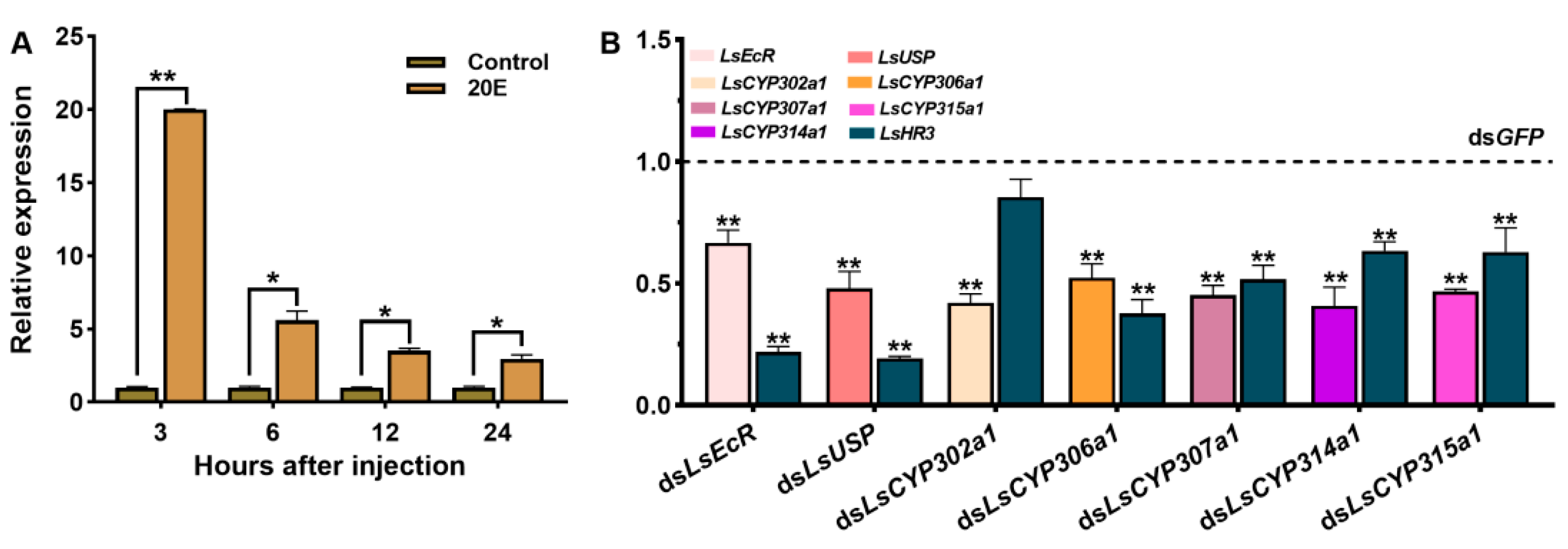
3.3. The Response of LsHR3 to 20E
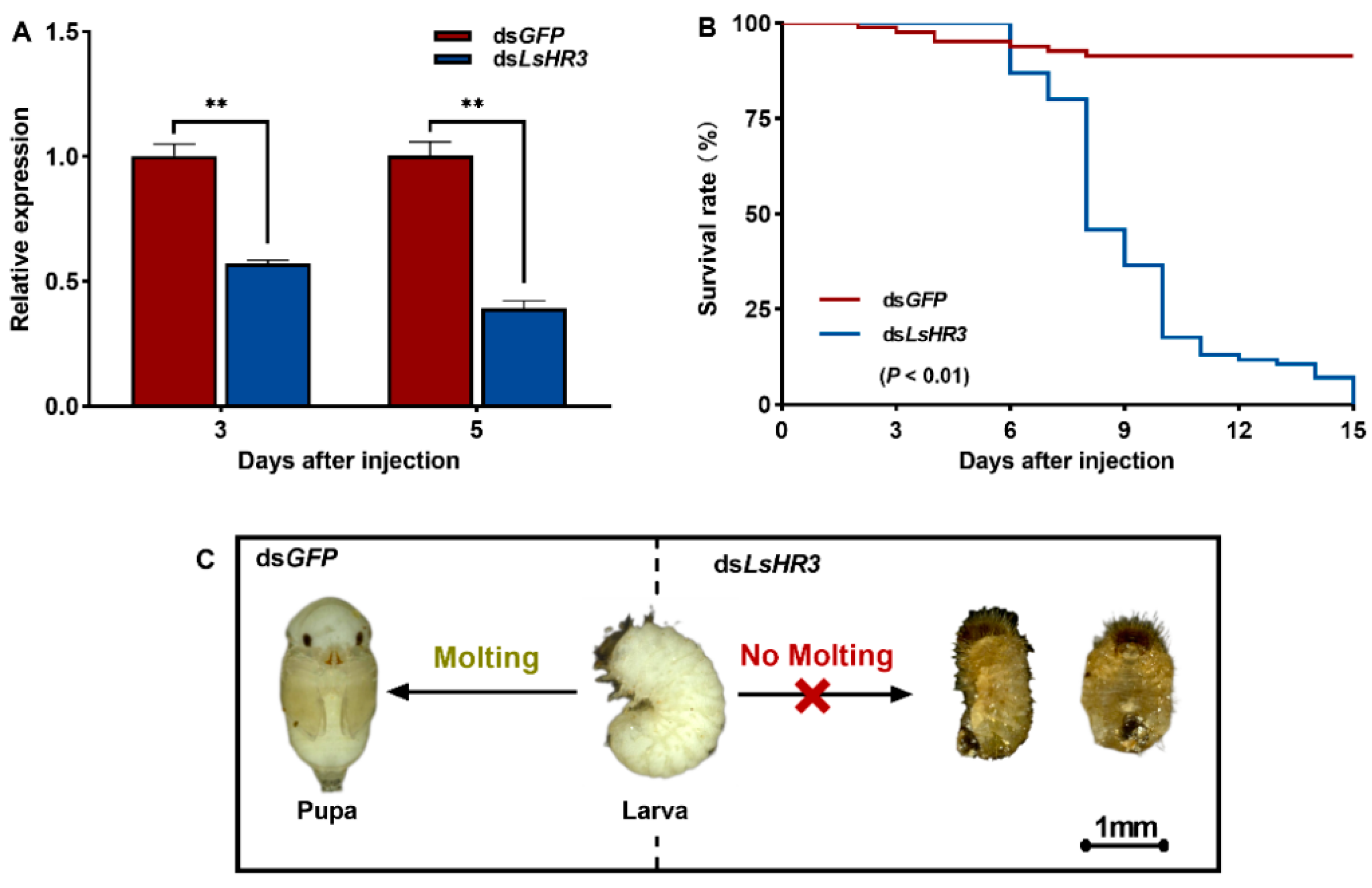
3.4. Knockdown of LsHR3 Disrupts the Larval–Pupal Molting
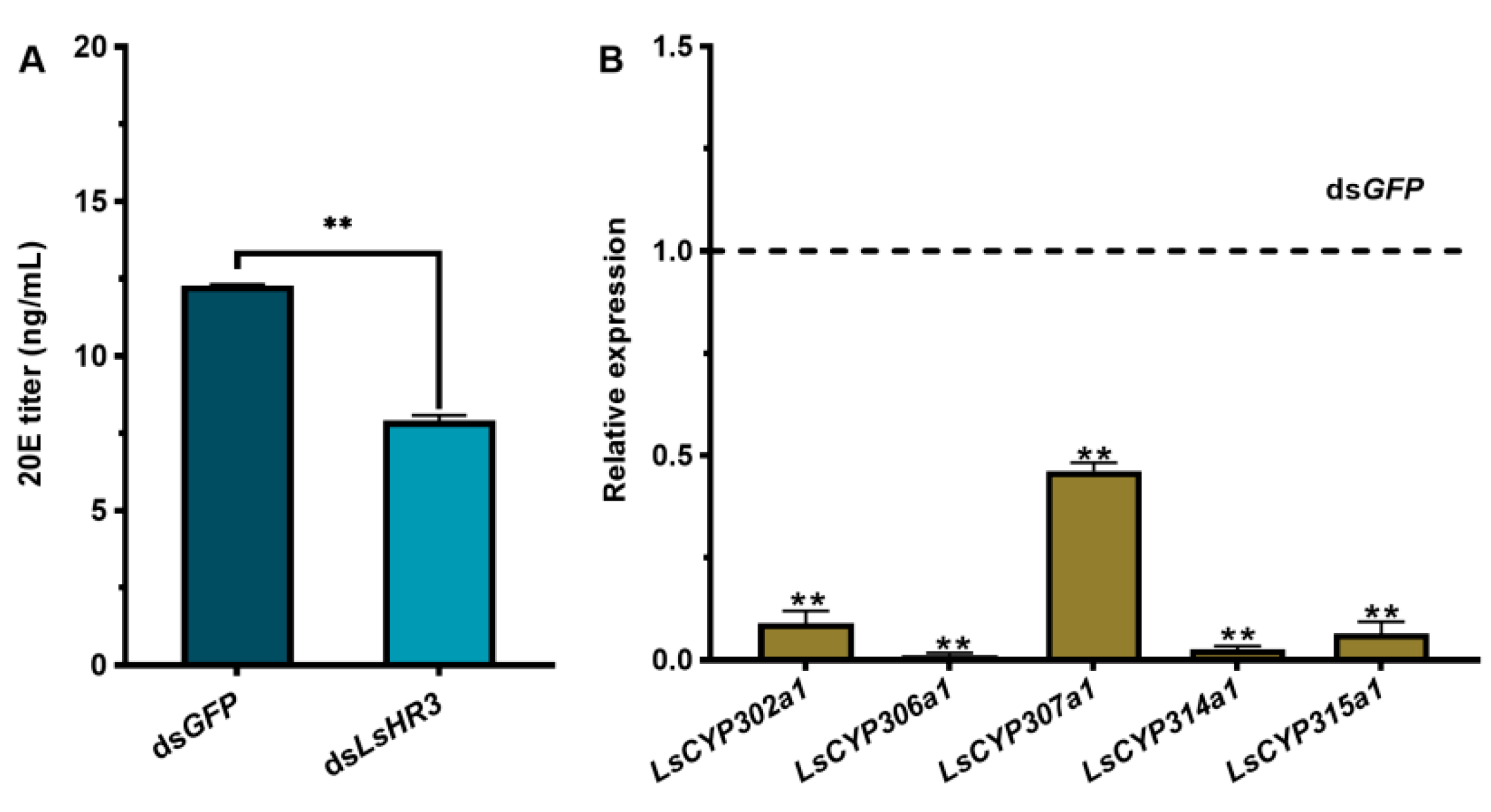
3.5. Knockdown of LsHR3 Disturbs 20E Synthesis
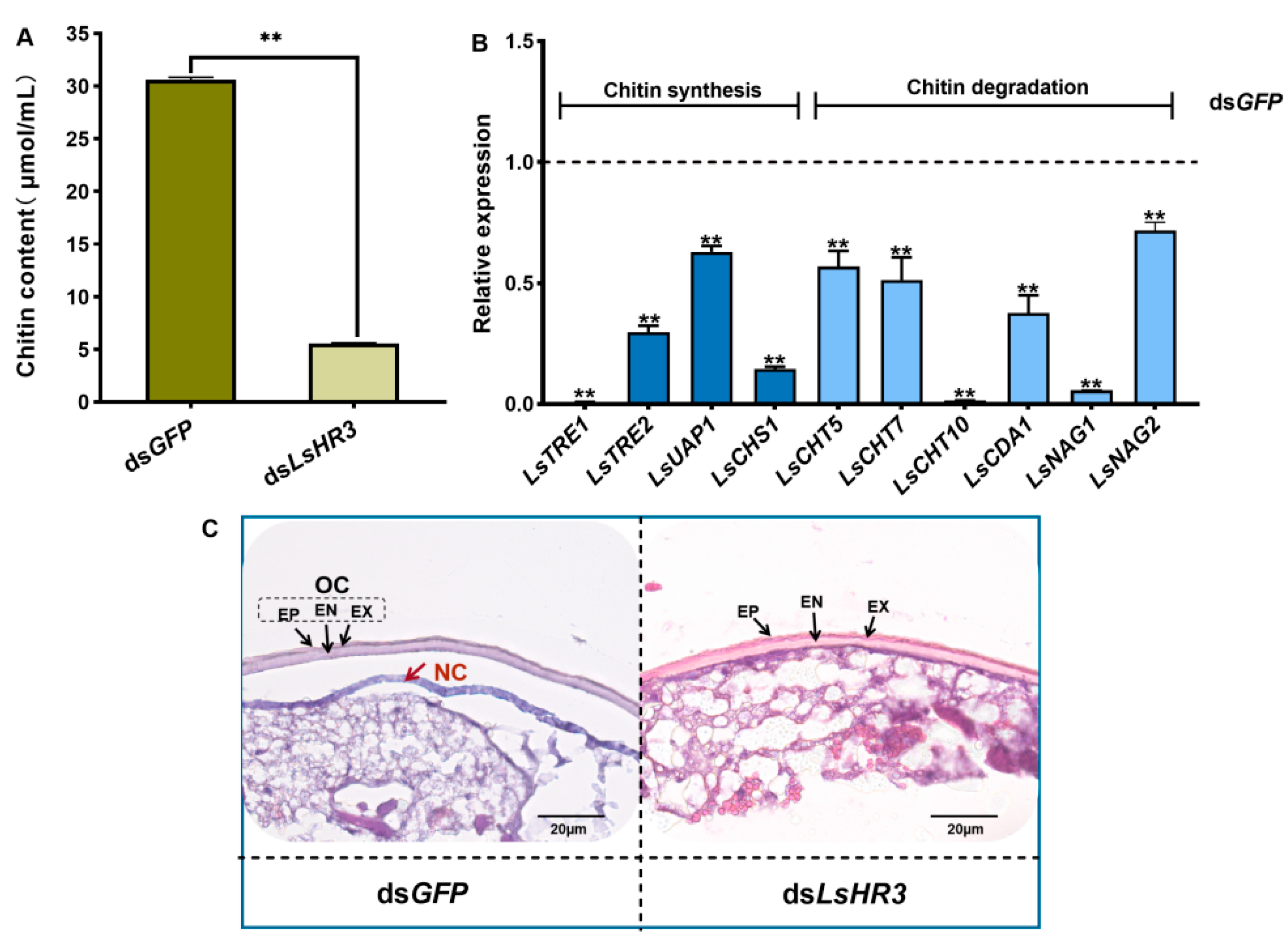
3.6. Knockdown of LsHR3 Inhibits the Chitin Metabolism and Cuticle Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Retief, E.; Nicholas, A. The cigarette beetle Lasioderma serricorne (F.) (Coleoptera: Anobii-dae): A serious herbarium pest. Bothalia 1988, 18, 97–99. [Google Scholar] [CrossRef]
- Sağlam, Ö.; Edde, P.A.; Phillips, T.W. Resistance of Lasioderma serricorne (Coleoptera: Anobiidae) to fumigation with phosphine. J. Econ. Entomol. 2015, 108, 2489–2495. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Chen, X.; Lin, T.; Cheng, X.S. Evaluation of beta-cyfluthrin resistance of cigarette beetle (Coleoptera: Anobiidae) from cigarette manufacturing factories of China and underlying metabolic mechanisms responsible for resistance. J. Econ. Entomol. 2021, 114, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, N.; Takahashi, R.; Matsuda, H.; Mikawa, Y.; Suzuki, T.; Suzuki, T.; Sonoda, S. Sodium channel mutations (T929I and F1534S) found in pyrethroid-resistant strains of the cigarette beetle, Lasioderma serricorne (Coleoptera: Anobiidae). J. Pestic. Sci. 2021, 46, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Kennerdell, J.R.; Carthew, R.W. Use of dsRNA-mediated genetic interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway. Cell 1998, 95, 1017–1026. [Google Scholar] [CrossRef]
- Jain, R.G.; Robinson, K.E.; Fletcher, S.J.; Mitter, N. RNAi-based functional genomics in Hemiptera. Insects 2020, 11, 557. [Google Scholar] [CrossRef]
- Lü, J.; Yang, C.X.; Liu, Z.Q.; Velez, A.M.; Guo, M.J.; Chen, S.M.; Qiu, B.L.; Zhang, Y.J.; Zhou, X.J.; Pan, H.P. Dietary RNAi toxicity assay suggests alpha and gamma subunits of HvCOPI as novel molecular targets for Henosepilachna vigintioctopunctata, an emerging coccinellid pest. J. Pest. Sci. 2021, 94, 1473–1486. [Google Scholar] [CrossRef]
- Sharma, R.; Christiaens, O.; Taning, C.N.; Smagghe, G. RNAi-mediated mortality in southern green stinkbug Nezara viridula by oral delivery of dsRNA. Pest Manag. Sci. 2021, 77, 77–84. [Google Scholar] [CrossRef]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef]
- Andersen, S.O.; Hojrup, P.; Roepstorff, P. Insect cuticular proteins. Insect Biochem. Mol. Biol. 1995, 25, 153–176. [Google Scholar] [CrossRef]
- Zitnan, D.; Kim, Y.J.; Zitnanová, I.; Roller, L.; Adams, M.E. Complex steroid-peptide-receptor cascade controls insect ecdysis. Gen. Comp. Endocr. 2007, 153, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.J.; Billas, I.M.; Bonneton, F.; Graham, L.D.; Lawrence, M.C. Ecdysone receptors: From the Ashburner model to structural biology. Annu. Rev. Entomol. 2013, 58, 251–271. [Google Scholar] [CrossRef]
- Riddiford, L.M.; Hiruma, K.; Zhou, X.; Nelson, C.A. Insights into the molecular basis of the hormonal control of molting and metamorphosis from Manduca sexta and Drosophila melanogaster. Insect Biochem. Mol. Biol. 2003, 33, 1327–1338. [Google Scholar] [CrossRef]
- Cruz, J.; Martín, D.; Bellés, X. Redundant ecdysis regulatory functions of three nuclear receptor HR3 isoforms in the direct-developing insect Blattella germanica. Mech. Develop. 2007, 124, 180–189. [Google Scholar] [CrossRef]
- Guo, W.C.; Liu, X.P.; Fu, K.Y.; Shi, J.F.; Lü, F.G.; Li, G.Q. Functions of nuclear receptor HR3 during larval-pupal molting in Leptinotarsa decemlineata (Say) revealed by in vivo RNA interference. Insect Biochem. Mol. Biol. 2015, 63, 23–33. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.Y.; Han, X.; Niu, J.Z.; Wang, J.J. RNAi of the nuclear receptor HR3 suggests a role in the molting process of the spider mite Panonychus citri. Exp. Appl. Acarol. 2020, 81, 75–83. [Google Scholar] [CrossRef]
- Fei, X.W.; Zhang, Y.; Ding, L.L.; Li, Y.J.; Deng, X.D. Controlling the development of the dengue vector Aedes aegypti using HR3 RNAi transgenic Chlamydomonas. PLoS ONE 2020, 15, e0240223. [Google Scholar] [CrossRef]
- Edde, P.A. Biology, ecology, and control of Lasioderma serricorne (F.) (Coleoptera: Anobiidae): A Review. J. Econ. Entomol. 2019, 112, 1011–1031. [Google Scholar] [CrossRef]
- Yang, W.J.; Xu, K.K.; Cao, Y.; Meng, Y.L.; Liu, Y.; Li, C. Identification and expression analysis of four small heat shock protein genes in cigarette beetle, Lasioderma serricorne (Fabricius). Insects 2019, 10, 139. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Yang, W.J.; Xu, K.K.; Yan, Y.; Li, C.; Jin, D.C. Role of Chitin deacetylase 1 in the molting and metamorphosis of the cigarette beetle Lasioderma serricorne. Int. J. Mol. Sci. 2020, 21, 2449. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.K.; Shen, X.M.; Ding, T.B.; Niu, J.Z.; Zhong, R.; Liao, C.Y.; Feng, Y.C.; Dou, W.; Wang, J.J. Functional analysis of a chitinase gene during the larval-nymph transition in Panonychus citri by RNA interference. Exp. Appl. Acarol. 2016, 70, 1–15. [Google Scholar] [CrossRef]
- Song, Y.; Villeneuve, D.L.; Toyota, K.; Iguchi, T.; Tollefsen, K.E. Ecdysone receptor agonism leading to lethal molting disruption in Arthropods: Review and adverse outcome pathway development. Environ. Sci. Technol. 2017, 51, 4142–4157. [Google Scholar] [CrossRef]
- Xiao, H.; Jeang, K.T. Glutamine-rich domains activate transcription in yeast Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 22873–22876. [Google Scholar] [CrossRef]
- Tan, A.; Palli, S.R. Identification and characterization of nuclear receptors from the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 430–439. [Google Scholar] [CrossRef]
- Sapin, G.D.; Tomoda, K.; Tanaka, S.; Shinoda, T.; Miura, K.; Minakuchi, C. Involvement of the transcription factor E75 in adult cuticular formation in the red flour beetle Tribolium castaneum. Insect Biochem. Mol. Biol. 2020, 126, 103450. [Google Scholar] [CrossRef]
- Shen, C.H.; Xu, Q.Y.; Mu, L.L.; Fu, K.Y.; Guo, W.C.; Li, G.Q. Involvement of Leptinotarsa hormone receptor 38 in the larval-pupal transition. Gene 2020, 751, 144779. [Google Scholar] [CrossRef]
- Zhao, X.M.; Qin, Z.Y.; Liu, W.M.; Liu, X.J.; Moussian, B.; Ma, E.B.; Li, S.; Zhang, J.Z. Nuclear receptor HR3 controls locust molt by regulating chitin synthesis and degradation genes of Locusta migratoria. Insect Biochem. Mol. Biol. 2018, 92, 1–11. [Google Scholar] [CrossRef]
- King-Jones, K.; Thummel, C.S. Nuclear receptors-a perspective from Drosophila. Nat. Rev. Genet. 2005, 6, 311–323. [Google Scholar] [CrossRef]
- Flynt, A.S. Insecticidal RNA interference, thinking beyond long dsRNA. Pest Manag. Sci. 2021, 77, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.M.; Chen, W.; Li, C.Z.; Ou, S.Y.; He, L. RNAi targeting ecdysone receptor blocks the larva to adult development of Tetranychus cinnabarinus. Pestic. Biochem. Physiol. 2019, 159, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, X.Y.; Smagghe, G.; Niu, J.Z.; Wang, J.J. Molting process revealed by the detailed expression profiles of RXR1/RXR2 and mining the associated genes in a spider mite, Panonychus citri. Insect Sci. 2022, 29, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.Y.; Meng, Q.W.; Deng, P.; Guo, W.C.; Li, G.Q. Leptinotarsa hormone receptor 4 (HR4) tunes ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis. Insect Biochem. Mol. Biol. 2018, 94, 50–60. [Google Scholar] [CrossRef]
- Street, S.M.; Eytcheson, S.A.; LeBlanc, G.A. The role of nuclear receptor E75 in regulating the molt cycle of Daphnia magna and consequences of its disruption. PLoS ONE 2019, 14, e0221642. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Takaki, K. Transcriptional regulation of cuticular genes during insect metamorphosis. Front. Biosci. 2020, 25, 106–117. [Google Scholar]
- Yogindran, S.; Rajam, M.V. Host-derived artificial miRNA-mediated silencing of ecdysone receptor gene provides enhanced resistance to Helicoverpa armigera in tomato. Genomics 2021, 113, 736–747. [Google Scholar] [CrossRef]
- Xu, Q.Y.; Deng, P.; Zhang, Q.; Li, A.; Fu, K.Y.; Guo, W.C.; Li, G.Q. Ecdysone receptor isoforms play distinct roles in larval-pupal-adult transition in Leptinotarsa decemlineata. Insect Sci. 2020, 27, 487–499. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.-X.; He, R.-T.; Yan, S.-Y.; Yang, W.-J. RNAi Suppression of Hormone Receptor HR3 Blocks Larval Molting and Metamorphosis in the Cigarette Beetle, Lasioderma serricorne. Agriculture 2022, 12, 1257. https://doi.org/10.3390/agriculture12081257
Ma L-X, He R-T, Yan S-Y, Yang W-J. RNAi Suppression of Hormone Receptor HR3 Blocks Larval Molting and Metamorphosis in the Cigarette Beetle, Lasioderma serricorne. Agriculture. 2022; 12(8):1257. https://doi.org/10.3390/agriculture12081257
Chicago/Turabian StyleMa, Li-Xin, Rong-Tao He, Shu-Yan Yan, and Wen-Jia Yang. 2022. "RNAi Suppression of Hormone Receptor HR3 Blocks Larval Molting and Metamorphosis in the Cigarette Beetle, Lasioderma serricorne" Agriculture 12, no. 8: 1257. https://doi.org/10.3390/agriculture12081257