
Genetic Diversity of Fish in Aquaculture and of Common Carp (Cyprinus carpio) in Traditional Rice–Fish Coculture

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
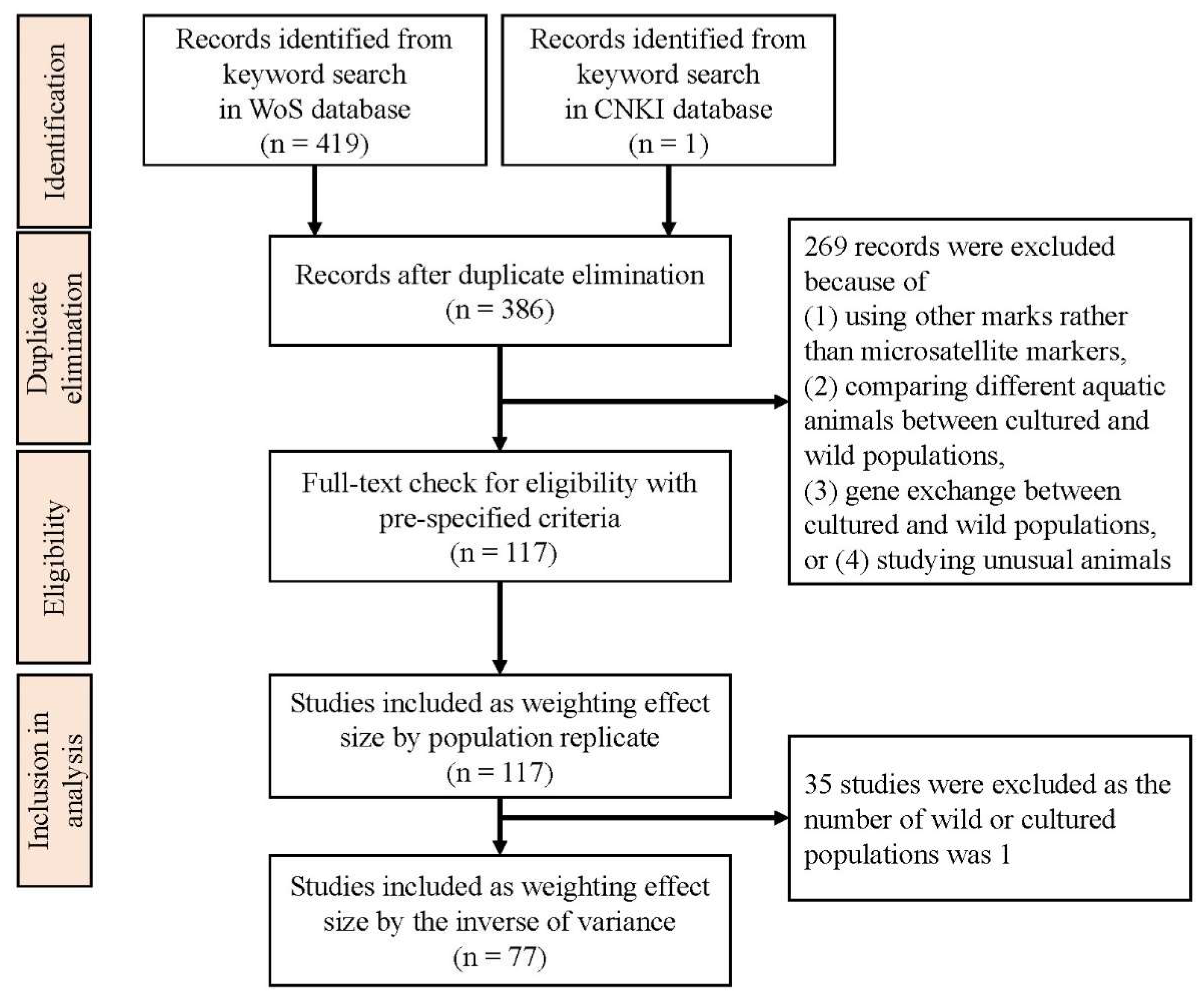
2.1. Meta-Analysis
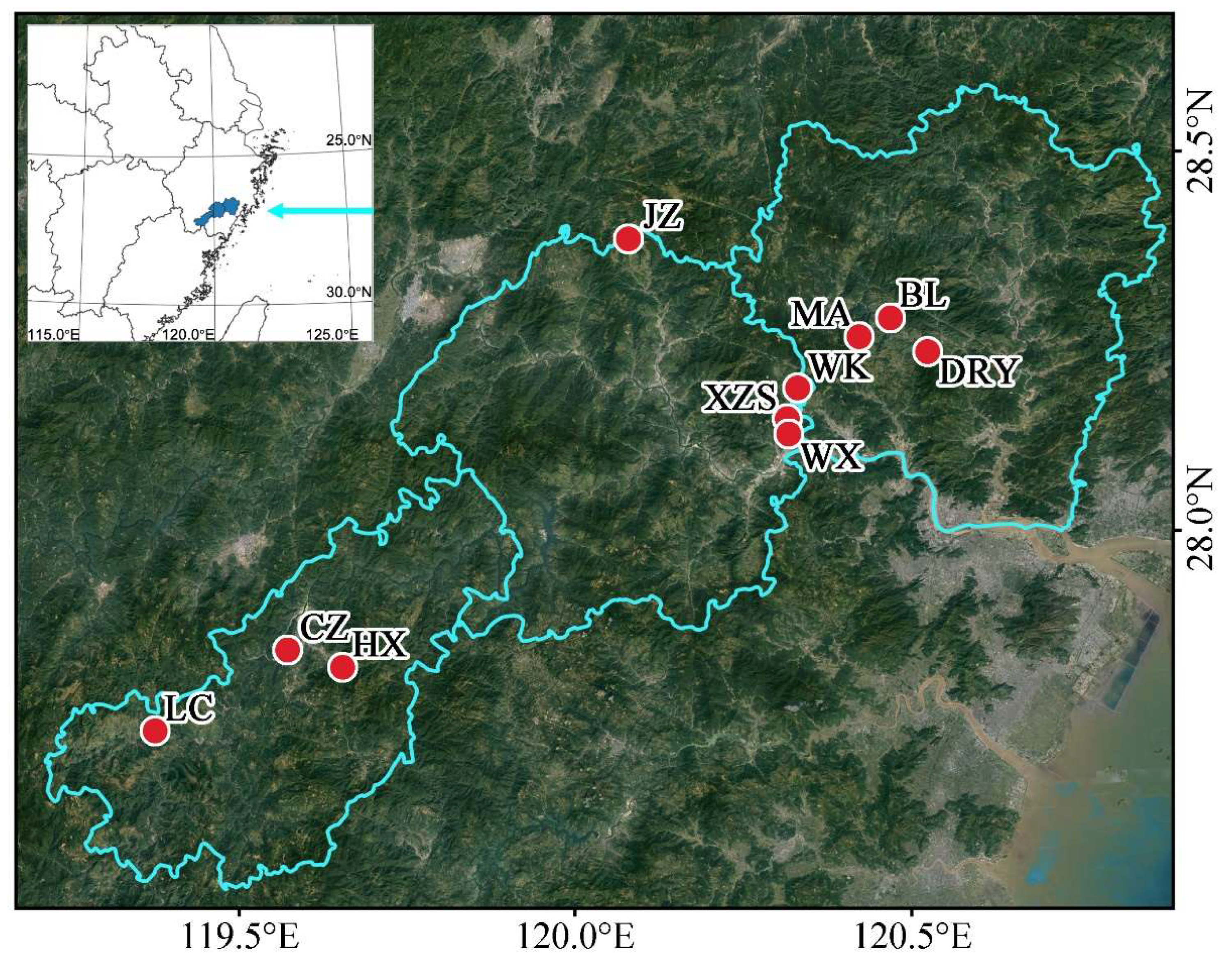
2.2. Sample Collection and DNA Extraction
2.3. Microsatellite Analysis
2.4. Genetic Data Analysis
2.4.1. Genetic Diversity
2.4.2. Genetic Variation
3. Results
3.1. Meta-Analysis
3.2. Genetic Diversity within Carp Populations in Rice–Fish Coculture
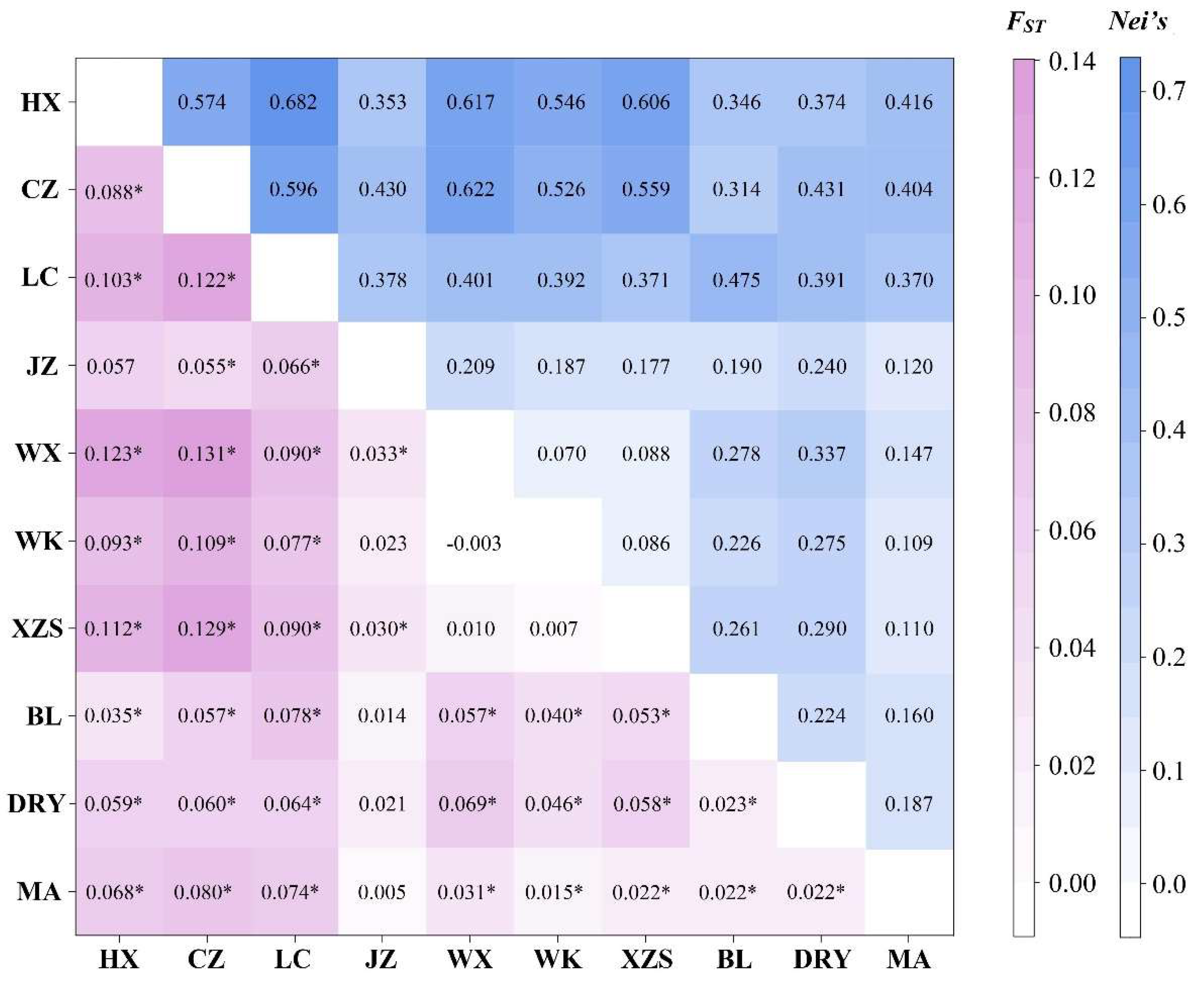
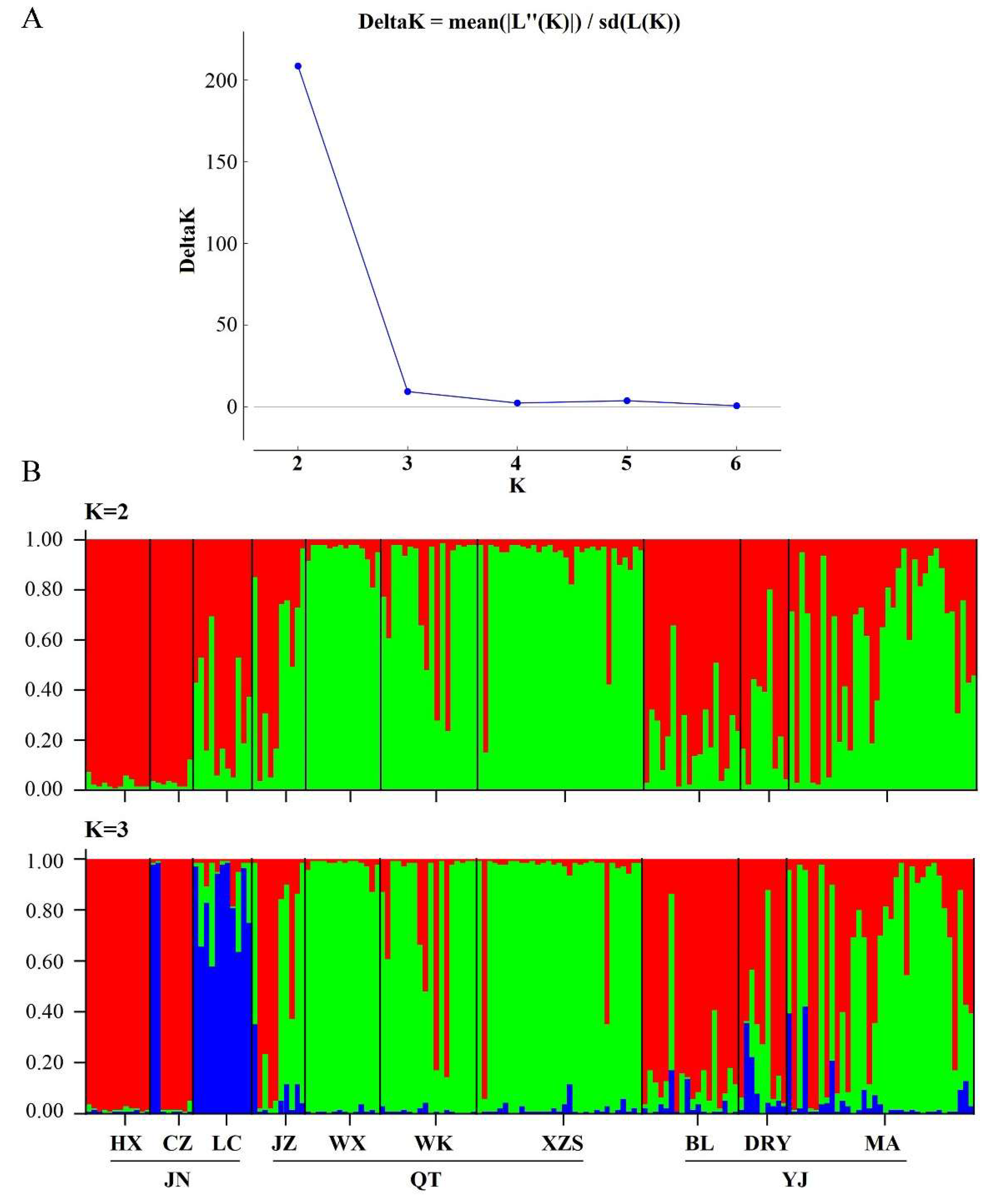
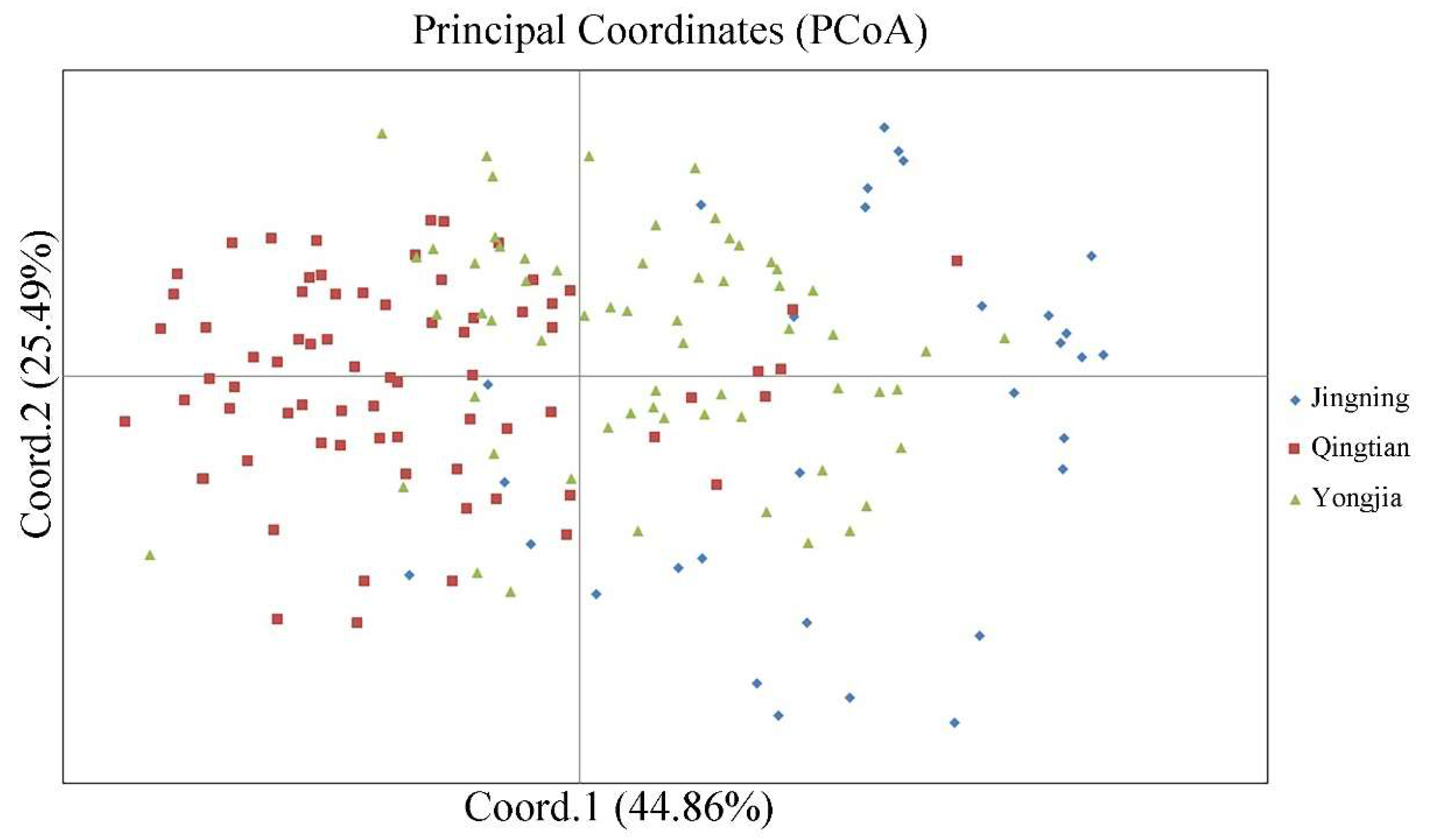
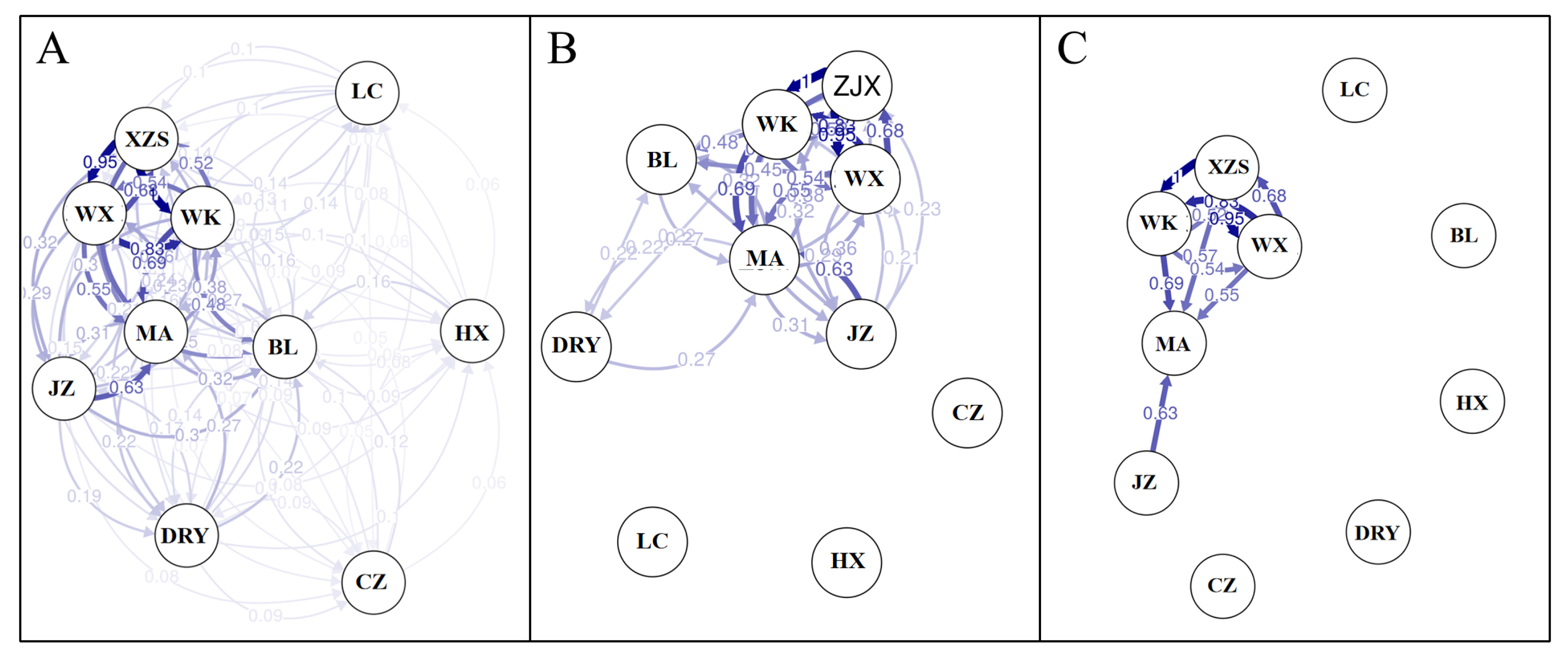
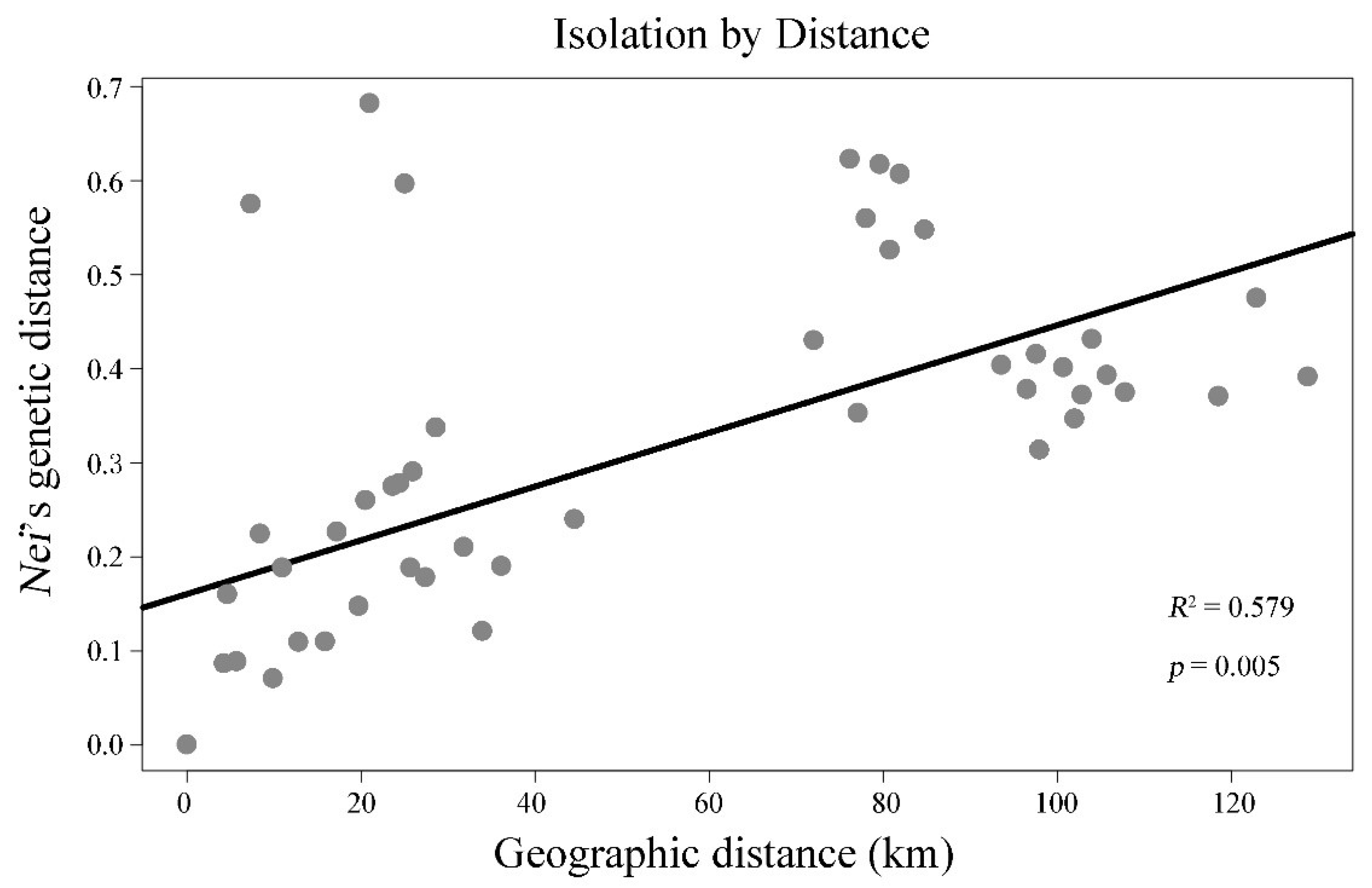
3.3. Genetic Differentiation among Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pingali, P.L. Green Revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; D’Amato, A.; Pallante, G. Benefit sharing mechanisms for agricultural genetic diversity use and on-farm conservation. Econ. Politica 2020, 37, 337–355. [Google Scholar] [CrossRef]
- Castilla, F.; Montilla-Bascón, G.; Bekele, W.A.; Howarth, C.J.; Prats, E. Population genomics of mediterranean oat (A. sativa) reveals high genetic diversity and three loci for heading date. Theor. Appl. Genet. 2021, 134, 2063–2077. [Google Scholar]
- van de Wouw, M.; Kik, C.; Hintum, T.; van Treuren, R.; Visser, B. Genetic erosion in crops: Concept, research results and challenges. Plant Genet. Resour. 2010, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Boettcher, P.J.; Hoffmann, I. Protecting indigenous livestock diversity. Science 2011, 334, 1058. [Google Scholar] [CrossRef]
- Aerts, R.; Berecha, G.; Honnay, O. Protecting coffee from intensification. Science 2015, 347, 139. [Google Scholar] [CrossRef] [Green Version]
- Rajpurohit, D.; Jhang, T. In situ and ex situ conservation of plant genetic resources and traditional knowledge. In Plant Genetic Resources and Traditional Knowledge for Food Security; Springer: Singapore, 2015; pp. 137–162. [Google Scholar]
- Priyanka, V.; Kumar, R.; Dhaliwal, I.; Kaushik, P. Germplasm conservation: Instrumental in agricultural biodiversity—A review. Sustainability 2021, 13, 6743. [Google Scholar] [CrossRef]
- Jarvis, D.I.; Brown, A.H.D.; Cuong, P.H.; Collado-Panduro, L.; Latournerie-Moreno, L.; Gyawali, S.; Tanto, T.; Sawadogo, M.; Mar, I.; Sadiki, M.; et al. A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc. Natl. Acad. Sci. USA 2008, 105, 8160. [Google Scholar] [CrossRef] [Green Version]
- Achtak, H.; Ater, M.; Oukabli, A.; Santoni, S.; Kjellberg, F.; Khadari, B. Traditional agroecosystems as conservatories and incubators of cultivated plant varietal diversity: The case of fig (Ficus carica L.) in Morocco. BMC Plant Biol. 2010, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Garlock, T.; Asche, F.; Anderson, J.; Bjørndal, T.; Kumar, G.; Lorenzen, K.; Tveteras, R. A global blue revolution: Aquaculture growth across regions, species, and countries. Rev. Fish Sci. Aquac. 2020, 28, 107–116. [Google Scholar] [CrossRef]
- Ahmad, A.; Abdullah, S.R.S.; Hasan, H.A.; Othman, A.R.; Ismail, N.I. Aquaculture industry: Supply and demand, best practices, effluent and its current issues and treatment technology. J. Environ. Manag. 2021, 287, 112271. [Google Scholar] [CrossRef] [PubMed]
- Napier, J.A.; Haslam, R.P.; Olsen, R.E.; Tocher, D.R.; Betancor, M.B. Agriculture can help aquaculture become greener. Nat. Food 2020, 1, 680–683. [Google Scholar] [CrossRef]
- Wang, C.; Li, Z.; Wang, T.; Xu, X.; Zhang, X.; Li, D. Intelligent fish farm—The future of aquaculture. Aquacult. Int. 2021, 29, 2681–2711. [Google Scholar] [CrossRef] [PubMed]
- Ju, R.T.; Li, X.; Jiang, J.J.; Wu, J.; Liu, J.; Strong, D.R.; Li, B. Emerging risks of non-native species escapes from aquaculture: Call for policy improvements in China and other developing countries. J. Appl. Ecol. 2020, 57, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.P.; Wong, L.L.; Razali, S.A.; Afiqah-Aleng, N.; Mohd Nor, S.A.; Sung, Y.Y.; de Peer, Y.V.; Sorgeloos, P.; Danish-Daniel, M. Applications of next-generation sequencing technologies and computational tools in molecular evolution and aquatic animals conservation studies: A short review. Evol. Bioinform. 2019, 15, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, J.D.P.; Gomes, P.F.F.; Hamoy, I.G.; Santos, S.E.B.D.; Schneider, H.; Sampaio, I. Loss of genetic variability in the captive stocks of tambaqui, Colossoma macropomum (Cuvier, 1818), at breeding centres in Brazil, and their divergence from wild populations. Aquac. Res. 2018, 49, 1914–1925. [Google Scholar] [CrossRef]
- Lind, C.E.; Evans, B.S.; Knauer, J.; Taylor, J.J.U.; Jerry, D.R. Decreased genetic diversity and a reduced effective population size in cultured silver-lipped pearl oysters (Pinctada maxima). Aquaculture 2009, 286, 12–19. [Google Scholar] [CrossRef]
- Loukovitis, D.; Ioannidi, B.; Chatziplis, D.; Kotoulas, G.; Magoulas, A.; Tsigenopoulos, C.S. Loss of genetic variation in Greek hatchery populations of the European sea bass (Dicentrarchus labrax L.) as revealed by microsatellite DNA analysis. Mediterr. Mar. Sci. 2015, 16, 197–200. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wu, X.; Tang, J.J.; Zhang, J.E.; Luo, S.M.; Chen, X. Conservation of traditional rice varieties in a Globally Important Agricultural Heritage System (GIAHS): Rice-fish coculture. Agric. Sci. China. 2011, 10, 101–105. Available online: http://www.fao.org/giahs/giahs-home/en/ (accessed on 10 March 2022). [CrossRef]
- Ren, W.; Hu, L.; Guo, L.; Zhang, J.; Tang, L.; Zhang, E.; Zhang, J.; Luo, S.; Tang, J.; Chen, X. Preservation of the genetic diversity of a local common carp in the agricultural heritage rice–fish system. Proc. Natl. Acad. Sci. USA 2018, 115, E546–E554. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Hu, L.; Tang, J.; Wu, X.; Li, N.; Yuan, Y.; Yang, H.; Zhang, J.; Luo, S.; Chen, X. Ecological mechanisms underlying the sustainability of the agricultural heritage rice–fish coculture system. Proc. Natl. Acad. Sci. USA 2011, 108, E1381–E1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Li, Q.; Ling, Q.F.; Wang, L.; Wang, D.P.; Tong, G.J. Study on the morphological differences of four common carp populations. J. Anhui Agric. Sci. 2011, 39, 15404–15405. [Google Scholar]
- Crooijmans, R.P.M.A.; Bierbooms, V.A.F.; Komen, J.; VanderPoel, J.J.; Groenen, M.A.M. Microsatellite markers in common carp (Cyprinus carpio L.). Anim. Genet. 1997, 28, 129–134. [Google Scholar] [CrossRef]
- David, L.; Rajasekaran, P.; Fang, J.; Hillel, J.; Lavi, U. Polymorphism in ornamental and common carp strains (Cyprinus carpio L.) as revealed by AFLP analysis and a new set of microsatellite markers. Mol. Genet. Genom. 2001, 266, 353–362. [Google Scholar] [CrossRef]
- Wei, D.W.; Lou, Y.D.; Sun, X.Y.; Shen, J.B. Screening of microsatellite markers in common carp. Zool. Res. 2001, 223, 238–241. [Google Scholar]
- Wang, D.; Liao, X.L.; Cheng, L.; Yu, X.M.; Tong, J.G. Development of novel EST-SSR markers in common carp by data mining from public EST sequences. Aquaculture 2007, 271, 558–574. [Google Scholar] [CrossRef]
- Yue, G.H.; Ho, M.Y.; Orban, L.; Komen, J. Microsatellites within genes and ESTs of common carp and their applicability in silver crucian carp. Aquaculture 2004, 234, 85–98. [Google Scholar] [CrossRef]
- Oosterhout, C.V.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 65: Genetic analysis in excel population genetic software for teaching and research an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Luikart, G.; Allendorf, F.; Cornuet, J.; Sherwin, W. Distortion of allele frequency distributions provides a test for recent population bottlenecks. J. Hered. 1998, 89, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Piry, S.; Luikart, G.; Cornuet, J.M. Bottleneck: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Luikart, G.; Cornuet, J.M. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv. Biol. 1998, 12, 228–237. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 35: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. Available online: http://taylor0.biology.ucla.edu/structureHarvester/ (accessed on 27 March 2022). [CrossRef]
- Sundqvist, L.; Keenan, K.; Zackrisson, M.; Prodöhl, P.; Kleinhans, D. Directional genetic differentiation and relative migration. Ecol. Evol. 2016, 6, 3461–3475. Available online: https://popgen.shinyapps.io/divMigrate-online/ (accessed on 29 March 2022). [CrossRef] [Green Version]
- Oksanen, J.; Kindt, R.; O’Hara, B. Vegan: R functions for vegetation ecologists. Date Access 2005, 15, 2014. [Google Scholar]
- Abdelrahman, H.; ElHady, M.; Alcivar-Warren, A.; Allen, S.; Al-Tobasei, R.; Bao, L.; Beck, B.; Blackburn, H.; Bosworth, B.; Buchanan, J.; et al. Aquaculture genomics, genetics and breeding in the United States: Current status, challenges, and priorities for future research. BMC Genom. 2012, 18, 1–23. [Google Scholar]
- Reading, B.J.; McGinty, A.S.; Clark, R.W.; Hopper, M.S.; Woods III, L.C.; Baltzegar, D.A. Genomic enablement of temperate bass aquaculture (family Moronidae). In Breeding and Culture of Perch and Bass; Science China Press (Chinese Academy of Sciences): Beijing, China, 2018. [Google Scholar]
- Andersen, L.K.; Clark, R.W.; Hopper, M.S.; Hodson, R.G.; Schilling, J.; Daniels, H.V.; Woods, L.C., III; Kovach, A.I.; Berlinsky, D.L.; Kenter, L.W.; et al. Methods of domestic striped bass (Morone saxatilis) spawning that do not require the use of any hormone induction. Aquaculture 2021, 533, 736025. [Google Scholar] [CrossRef]
- Gibson, J.; Gamage, S.; Hanotte, O.; Iñiguez, L.; Maillard, J.; Rischkowsky, B.; Semambo, D.; Toll, J.; Gibson, J. Options and Strategies for the Conservation of Farm Animal Genetic Resources: Report of an International Workshop, AGROPOLIS, Montpellier, France, 7–10 November 2005; CGIAR System-Wide Genetic Resources Programme (SGRP)/Bioversity International: Rome, Italy, 2006; Volume 5. [Google Scholar]
- Harvey, B. Blue Genes: Sharing and Conserving the World’s Aquatic Biodiversity; Routledge: London, UK, 2013. [Google Scholar]
- Norris, A.T.; Bradley, D.G.; Cunningham, E.P. Microsatellite genetic variation between and within farmed and wild Atlantic salmon (Salmo salar) populations. Aquaculture 1999, 180, 247–264. [Google Scholar] [CrossRef]
- Machado-Schiaffino, G.; Dopico, E.; Garcia-Vazquez, E. Genetic variation losses in Atlantic salmon stocks created for supportive breeding. Aquaculture 2007, 264, 59–65. [Google Scholar] [CrossRef]
- Fazzi-Gomes, P.F.; Aguiar, J.D.P.; Marques, D.; Fonseca Cabral, G.; Moreira, F.C.; Rodrigues, M.D.N.; Silva, C.S.; Hamoy, I.; Santos, S. Novel microsatellite markers used for determining genetic diversity and tracing of wild and farmed populations of the Amazonian giant fish Arapaima gigas. Genes 2021, 12, 1324. [Google Scholar] [CrossRef]
- Danzmann, R.G.; Ferguson, M.M.; Allendorf, F.W. Genetic variability and components of fitness in hatchery strains of rainbow trout. J. Fish Biol. 1989, 35, 313–319. [Google Scholar] [CrossRef]
- Koehn, R.K.; Diehl, W.J.; Scott, T.M. The different contribution by individual enzymes of glycolysis and protein catabolism to the relationship between heterozygosity and growth rate in the coot clam Milinia lateralis. Genetics 1988, 118, 121–130. [Google Scholar] [CrossRef]
- Tiira, K.; Laurila, A.; Peuhkuri, N.; Piironen, J.; Ranta, E.; Primmer, C.R. Aggressiveness is associated with genetic diversity in landlocked salmon (Salmo salar). Mol. Ecol. 2003, 12, 2399–2407. [Google Scholar] [CrossRef]
- Ponzoni, R.W.; Nguyen, N.H.; Khaw, H.L.; Ninh, N.H. Accounting for genotype by environment interaction in economic appraisal of genetic improvement programs in common carp Cyprinus carpio. Aquaculture 2008, 285, 47–55. [Google Scholar] [CrossRef]
- Ninh, N.H.; Ponzoni, R.W.; Nguyen, N.H.; Woolliams, J.A.; Taggart, J.B.; McAndrew, B.J.; Penman, D.J. A comparison of communal and separate rearing of families in selective breeding of common carp (Cyprinus carpio): Responses to selection. Aquaculture 2013, 408, 152–159. [Google Scholar] [CrossRef]
- Gjerde, B. Design of breeding programs. In Selection and Breeding Programs in Aquaculture; Gjedrem, T., Ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 173–195. [Google Scholar]
- Loughnan, S.R.; Smith-Keune, C.; Jerry, D.R.; Beheregaray, L.B.; Robinson, N.A. Genetic diversity and relatedness estimates for captive barramundi (Lates calcarifer, Bloch) broodstock informs efforts to form a base population for selective breeding. Aquac. Res. 2016, 47, 3570–3584. [Google Scholar] [CrossRef]
- Abebe, A.T.; Kolawole, A.O.; Unachukwu, N.; Chigeza, G.; Gedil, M. Assessment of diversity in tropical soybean (Glycine max (L.) Merr.) varieties and elite breeding lines using single nucleotide polymorphism markers. Plant Genet. Resour. 2021, 19, 20–28. [Google Scholar] [CrossRef]
- Ren, W.Z. The Genetic Diversity in Traditional Rice-Fish System. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2016. [Google Scholar]
- Rahman, M.L.; Shahjahan, M.; Ahmed, N. Tilapia farming in Bangladesh: Adaptation to climate change. Sustainability 2021, 13, 7657. [Google Scholar] [CrossRef]
- Ji, D.; Su, X.; Yao, J.; Zhang, W.; Wang, R.; Zhang, S. Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China. Animals 2022, 12, 1377. [Google Scholar] [CrossRef] [PubMed]
- Pusadee, T.; Jamjod, S.; Chiang, Y.C.; Rerkasem, B.; Schaal, B.A. Genetic structure and isolation by distance in a landrace of Thai rice. Proc. Natl. Acad. Sci. USA 2009, 106, 13880–13885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, N.; Garine, E.; Khasah, C.; Dounias, E.; Hossaert-McKey, M.; McKey, D. Farmers’ practices, metapopulation dynamics, and conservation of agricultural biodiversity on-farm: A case study of sorghum among the Duupa in sub-sahelian Cameroon. Biol. Conserv. 2005, 121, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, C.; D’Eeckenbrugge, G.C. Social organization of crop genetic diversity. The G × E × S Interaction Model. Diversity 2011, 4, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Labeyrie, V.; Thomas, M.; Muthamia, Z.K.; Leclerc, C. Seed exchange networks, ethnicity, and sorghum diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Delêtre, M.; McKey, D.B.; Hodkinson, T.R. Marriage exchanges, seed exchanges, and the dynamics of manioc diversity. Proc. Natl. Acad. Sci. USA 2011, 108, 18249–18254. [Google Scholar] [CrossRef] [Green Version]
- Bashir, M.A.; Liu, J.; Geng, Y.; Wang, H.; Pan, J.; Zhang, D.; Rehim, A.; Aon, M.; Liu, H. Co-culture of rice and aquatic animals: An integrated system to achieve production and environmental sustainability. J. Clean. Prod. 2020, 249, 119310. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Village | Abbreviation | Sample Size | Geographic Locations |
|---|---|---|---|---|
| Jingning | Hexi | HX | 12 | 119.69° E 27.93° N |
| Chengzhao | CZ | 8 | 119.61° E 27.96° N | |
| Luci | LC | 11 | 119.40° E 27.87° N | |
| Qingtian | Jizhai | JZ | 10 | 120.18° E 28.46° N |
| Wenxi | WX | 14 | 120.39° E 28.18° N | |
| Wukeng | WK | 18 | 120.41° E 28.24° N | |
| Xiaozhoushan | XZS | 31 | 120.39° E 28.20° N | |
| Yongjia | Bilian | BL | 18 | 120.56° E 28.32° N |
| Daruoyan | DRY | 9 | 120.61° E 28.27° N | |
| Minao | MA | 35 | 120.51° E 28.30° N |
| Class | Studies | Weight | Genetic Diversity Indices | Effect Size | CI-l | CI-u | Decrease (%) |
|---|---|---|---|---|---|---|---|
| Salmon | 21 | 1/Var | Na | −0.30 | −0.18 | −0.43 | 26 |
| Flounder | 3 | 1/Var | Na | −0.49 | −0.24 | −0.65 | 38 |
| Perch | 10 | 1/Var | Na | −0.14 | 0.01 | −0.32 | 13 |
| Arthropod | 3 | 1/Var | Na | −0.05 | 0.01 | −0.62 | 5 |
| Mollusk | 16 | 1/Var | Na | −0.22 | −0.07 | −0.69 | 20 |
| Carp | 14 | 1/Var | Na | −0.13 | −0.03 | −0.23 | 12 |
| Herring | 2 | 1/Var | Na | −0.43 | −0.13 | −0.45 | 35 |
| Catfish | 3 | 1/Var | Na | −0.07 | 0.03 | −0.28 | 7 |
| Arapaima | 2 | 1/Var | Na | −0.07 | −0.04 | −0.24 | 6 |
| Overall | 74 | 1/Var | Na | −0.23 | −0.16 | −0.32 | 20 |
| Salmon | 21 | 1/Var | He | −0.11 | −0.05 | −0.23 | 11 |
| Flounder | 3 | 1/Var | He | −0.03 | −0.01 | −0.12 | 3 |
| Perch | 9 | 1/Var | He | −0.04 | −0.03 | −0.07 | 4 |
| Arthropod | 3 | 1/Var | He | −0.01 | 0.00 | −0.10 | 1 |
| Mollusk | 16 | 1/Var | He | −0.05 | −0.02 | −0.13 | 5 |
| Carp | 14 | 1/Var | He | −0.05 | −0.02 | −0.09 | 5 |
| Herring | 2 | 1/Var | He | −0.07 | −0.02 | −0.12 | 7 |
| Catfish | 3 | 1/Var | He | 0.00 | 0.05 | −0.06 | 0 |
| Arapaima | 2 | 1/Var | He | −0.09 | −0.09 | −0.19 | 9 |
| Overall | 73 | 1/Var | He | −0.08 | −0.04 | −0.13 | 8 |
| Salmon | 25 | Np | Na | −0.13 | 0.06 | −0.27 | 12 |
| Flounder | 7 | Np | Na | −0.24 | 0.12 | −0.50 | 21 |
| Perch | 20 | Np | Na | −0.27 | −0.09 | −0.46 | 24 |
| Arthropod | 3 | Np | Na | −0.23 | 0.01 | −0.62 | 21 |
| Mollusk | 28 | Np | Na | −0.31 | −0.11 | −0.62 | 27 |
| Carp | 16 | Np | Na | −0.28 | −0.14 | −0.55 | 24 |
| Echinoderm | 2 | Np | Na | −0.36 | 0.09 | −0.52 | 31 |
| Herring | 3 | Np | Na | −0.16 | −0.13 | −0.25 | 15 |
| Catfish | 4 | Np | Na | −0.36 | 0.24 | −0.81 | 30 |
| Puffer | 3 | Np | Na | −0.25 | −0.07 | −0.54 | 22 |
| Overall | 111 | Np | Na | −0.24 | −0.15 | −0.33 | 21 |
| Salmon | 25 | Np | He | −0.02 | 0.07 | −0.09 | 2 |
| Flounder | 7 | Np | He | −0.08 | −0.05 | −0.10 | 7 |
| Perch | 19 | Np | He | −0.06 | −0.01 | −0.11 | 6 |
| Arthropod | 3 | Np | He | −0.03 | 0.00 | −0.07 | 3 |
| Mollusk | 28 | Np | He | −0.06 | −0.01 | −0.12 | 6 |
| Carp | 16 | Np | He | −0.11 | −0.05 | −0.14 | 10 |
| Echinoderm | 2 | Np | He | 0.00 | 0.01 | −0.03 | 0 |
| Herring | 2 | Np | He | −0.01 | 0.00 | −0.02 | 1 |
| Catfish | 4 | Np | He | −0.04 | 0.04 | −0.09 | 4 |
| Puffer | 3 | Np | He | −0.05 | −0.01 | −0.12 | 4 |
| Overall | 109 | Np | He | −0.05 | −0.03 | −0.07 | 5 |
| Pop. | Na | Ne | I | Ho | He | Fis |
|---|---|---|---|---|---|---|
| HX | 5.80 | 4.35 | 1.53 | 0.75 | 0.74 | 0.00 |
| CZ | 7.00 | 4.70 | 1.63 | 0.76 | 0.73 | −0.02 |
| LC | 5.85 | 3.86 | 1.44 | 0.68 | 0.69 | 0.01 |
| JZ | 7.25 | 4.52 | 1.64 | 0.68 | 0.74 | 0.08 |
| WX | 5.85 | 3.92 | 1.42 | 0.68 | 0.68 | 0.00 |
| WK | 7.50 | 4.42 | 1.59 | 0.71 | 0.72 | 0.03 |
| XZS | 7.75 | 4.07 | 1.54 | 0.70 | 0.71 | 0.01 |
| BL | 9.65 | 5.70 | 1.85 | 0.76 | 0.77 | 0.03 |
| DRY | 6.90 | 4.81 | 1.62 | 0.70 | 0.73 | 0.07 |
| MA | 10.40 | 5.29 | 1.83 | 0.72 | 0.76 | 0.06 |
| Mean | 7.40 | 4.57 | 1.61 | 0.71 | 0.73 | 0.03 |
| Pop. | Effective Population Size Estimate | 95% Confidence Intervals | Bottleneck Test | |
|---|---|---|---|---|
| Lower Bound | Upper Bound | TPM (p-Value) | ||
| HX | 9.9 | 8.0 | 12.5 | 0.63575 |
| CZ | 8.6 | 5.7 | 13.6 | 0.06155 |
| LC | 61.5 | 27.6 | Inf | 0.83501 |
| JZ | Inf | 148.9 | Inf | 0.99585 |
| WX | 124.8 | 46.4 | Inf | 0.47816 |
| WK | 61.8 | 41.4 | 114.2 | 0.98802 |
| XZS | 54.2 | 44.0 | 69.3 | 0.99928 |
| BL | 209.6 | 98.4 | Inf | 0.99884 |
| DRY | Inf | 58.3 | Inf | 0.87726 |
| MN | 38.0 | 34.1 | 42.6 | 0.99940 |
| Total | 63.4 | 60.2 | 66.9 | 0.99985 |
| Source of Variation | d.f. | Sum of Square | Variance Component | % of Variation | p Value |
|---|---|---|---|---|---|
| Among populations | 9 | 173.541 | 0.37651 Va | 5 | 0.001 |
| Within populations | 165 | 2311.173 | 7.17756 Vb | 95 | 0.001 |
| Total | 174 | 2484.714 | 7.55407 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Ren, W.; Zhang, S.; Zhao, L.; Tang, J.; Hu, L.; Chen, X. Genetic Diversity of Fish in Aquaculture and of Common Carp (Cyprinus carpio) in Traditional Rice–Fish Coculture. Agriculture 2022, 12, 997. https://doi.org/10.3390/agriculture12070997
Ye Y, Ren W, Zhang S, Zhao L, Tang J, Hu L, Chen X. Genetic Diversity of Fish in Aquaculture and of Common Carp (Cyprinus carpio) in Traditional Rice–Fish Coculture. Agriculture. 2022; 12(7):997. https://doi.org/10.3390/agriculture12070997
Chicago/Turabian StyleYe, Yingying, Weizheng Ren, Shixiang Zhang, Lufeng Zhao, Jianjun Tang, Liangliang Hu, and Xin Chen. 2022. "Genetic Diversity of Fish in Aquaculture and of Common Carp (Cyprinus carpio) in Traditional Rice–Fish Coculture" Agriculture 12, no. 7: 997. https://doi.org/10.3390/agriculture12070997