
Evaluation of Cadmium Effects on Six Solanum melongena L. Cultivars from the Mediterranean Basin

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Germination Assays at Different Cd Concentrations
2.3. Hydroponic Culture of Plantlets at Different Cd Concentrations
2.4. Ionome Analysis, Bioaccumulation Coefficients and the Translocation Index
2.5. Photosynthetic Pigments Analysis
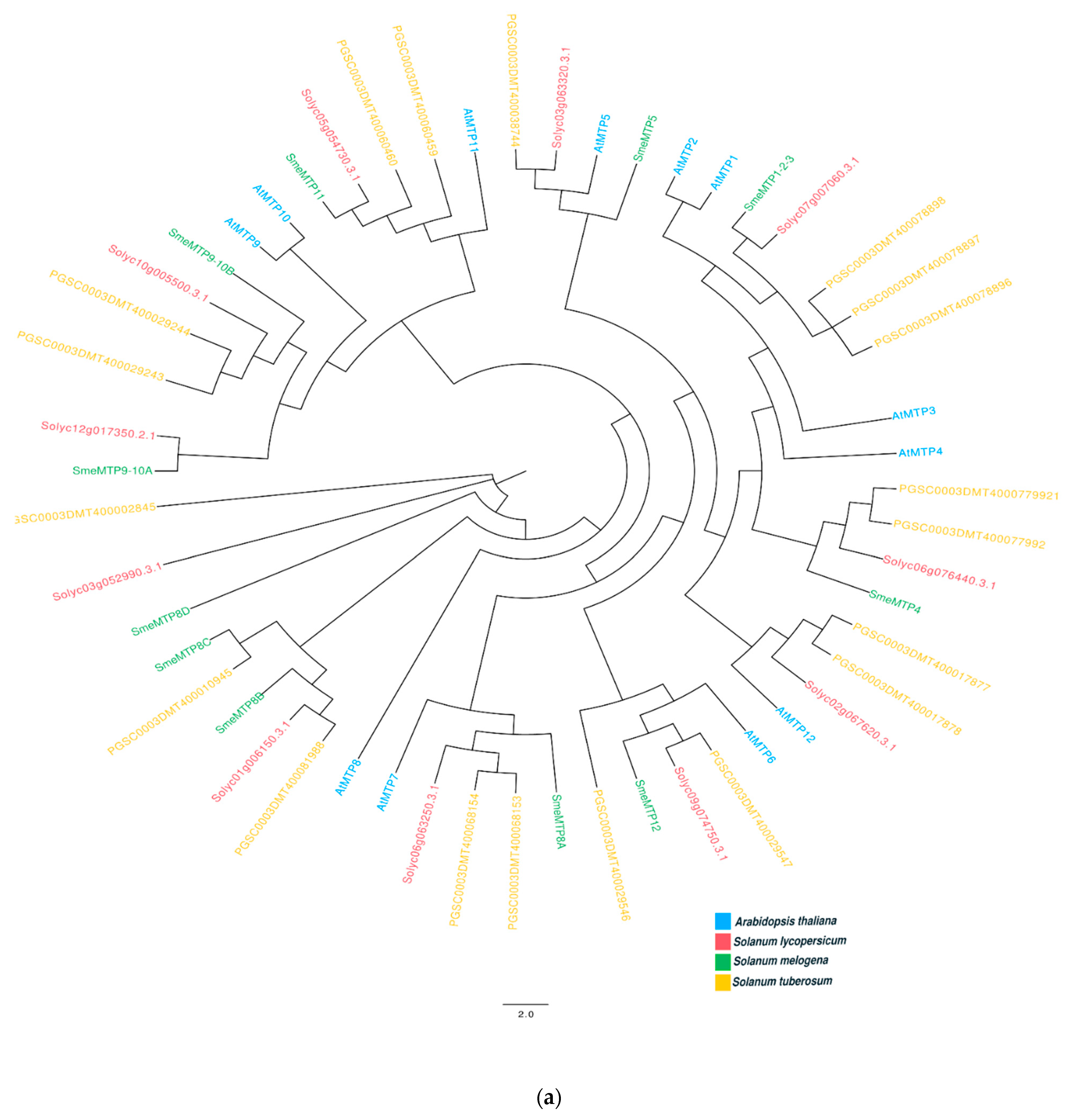
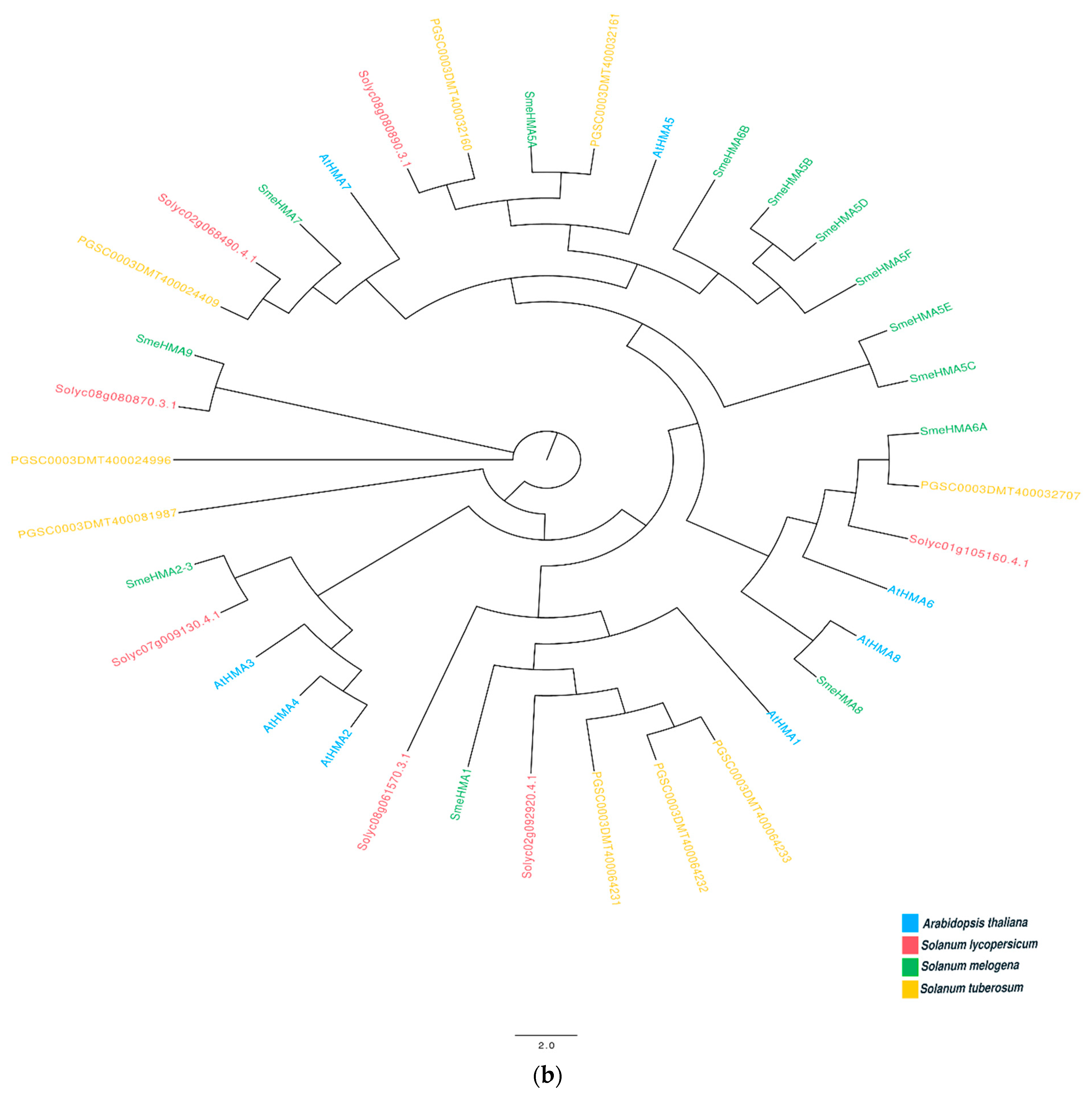
2.6. In Silico Identification of Genes Belonging to MTP and HMA Family
2.7. Quantitative PCR (qPCR) Analysis of Gene Expression
2.8. Statistical Analysis
3. Results
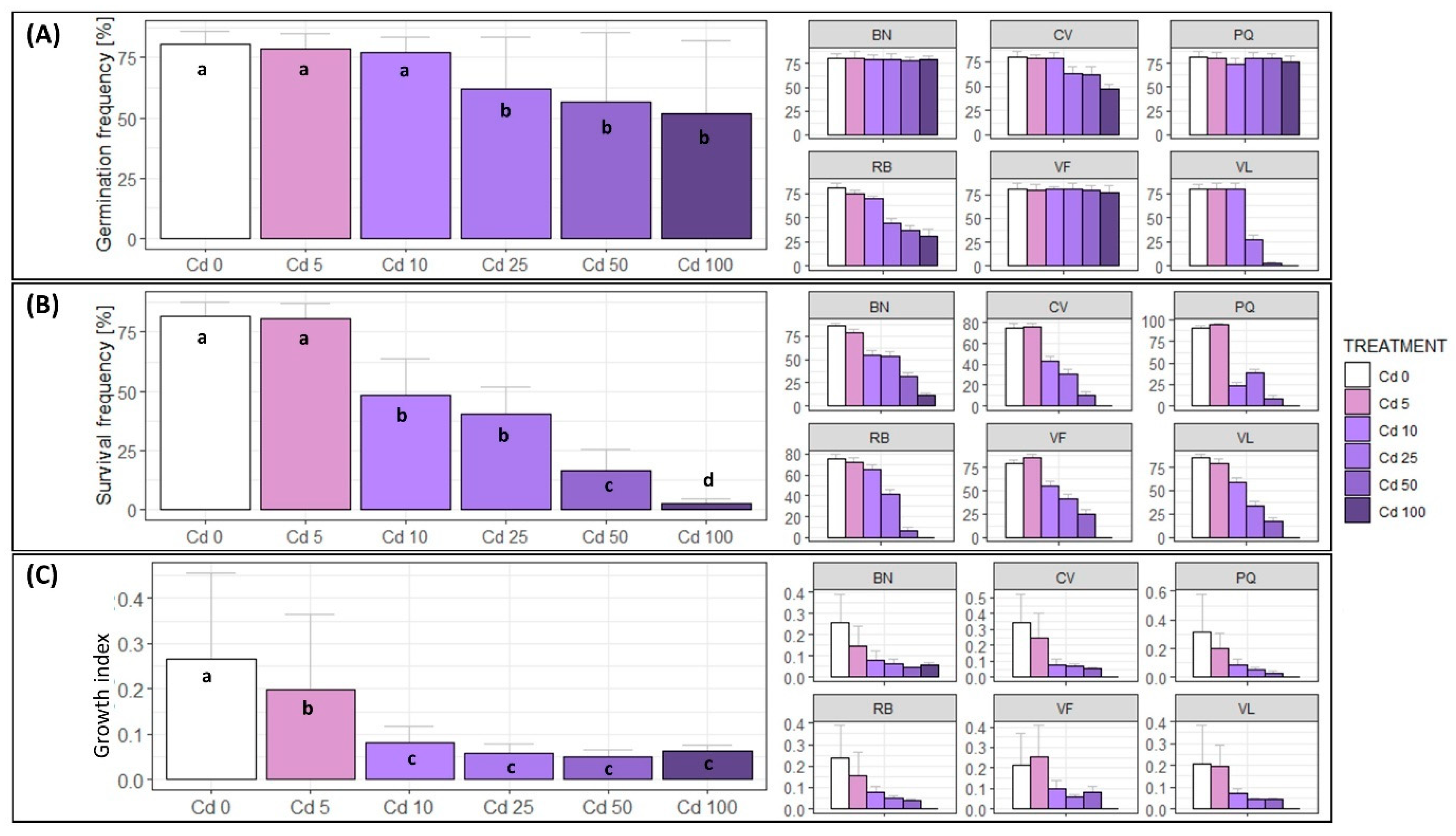
3.1. Germination Assays at Different Cd Concentrations
3.2. Hydroponic Culture of Plantlets at Different Cd Concentrations
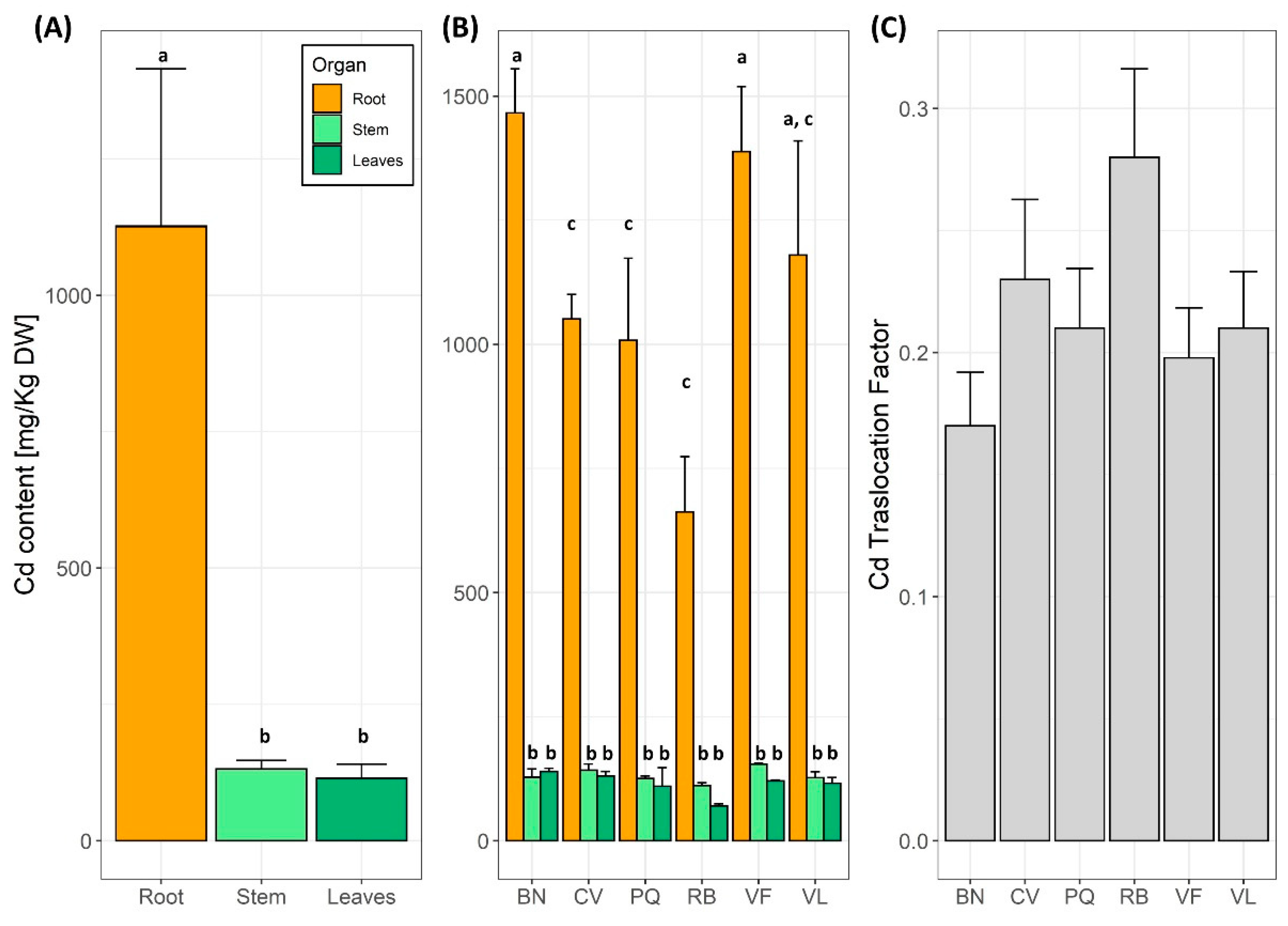
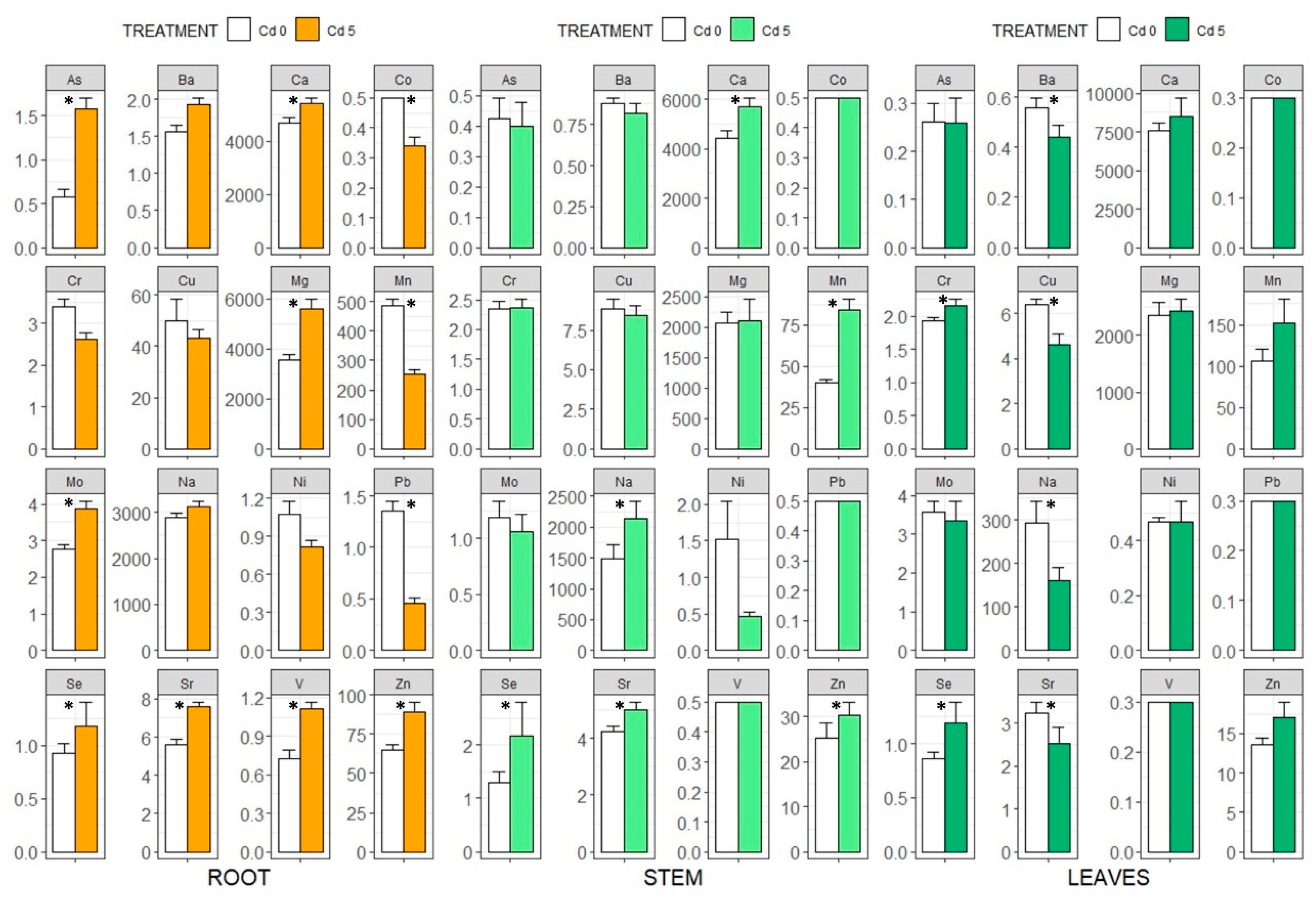
3.3. Ionome Analysis, Bioaccumulation Coefficients and the Translocation Index
3.4. Analysis of Photosynthetic Pigments
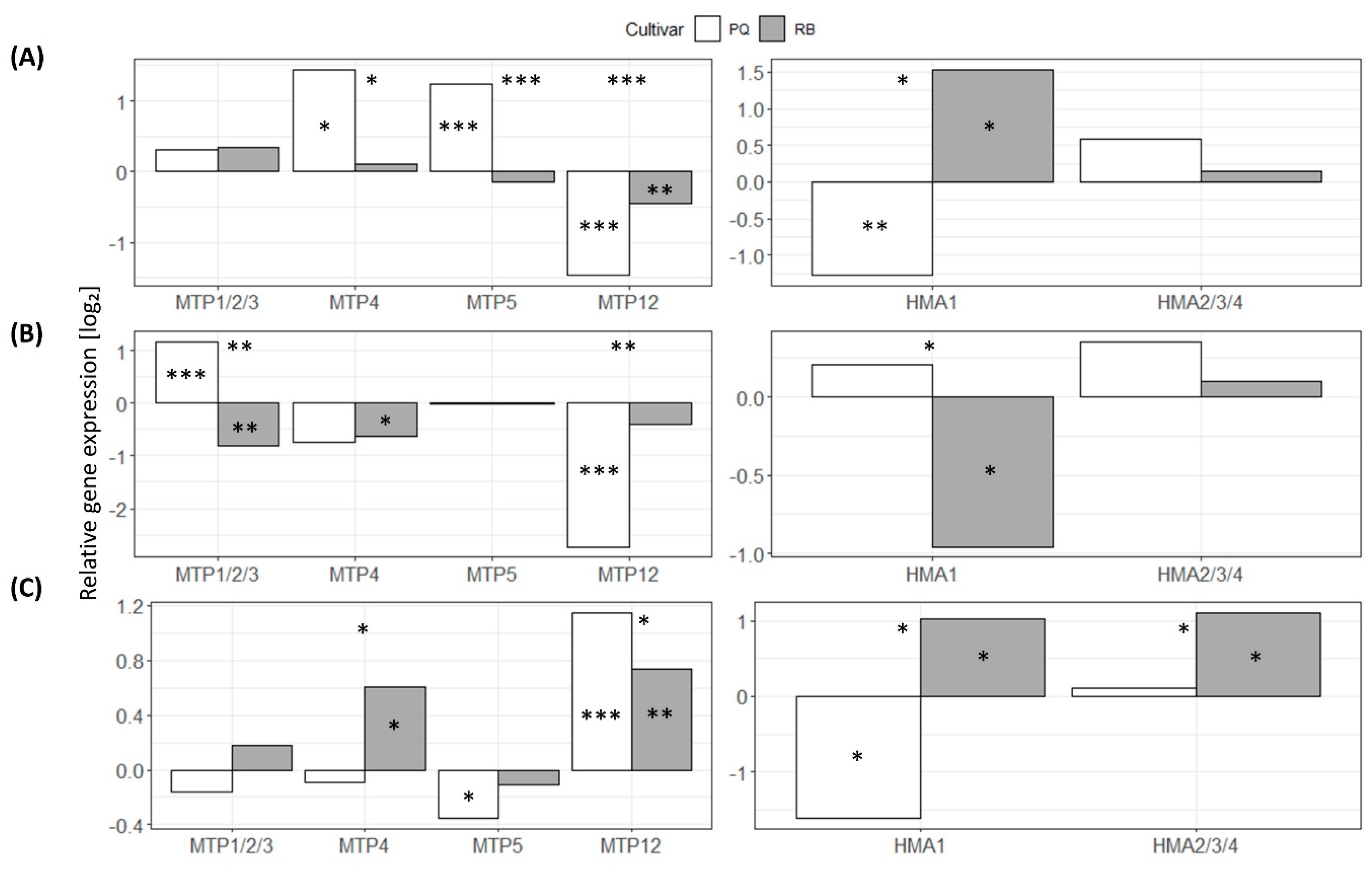
3.5. Quantitative Analysis of Gene Expression of HMA and MTP Genes
4. Discussion
4.1. Effects of Cd on Eggplant Germination
4.2. Effects of Cd on Eggplant Survival and Growth Rates
4.3. Effects of Cd on Eggplant Accumulation and Translocation
4.4. Effects of Cd on Eggplant on Photosynthetic Pigments Content
4.5. Effects of Cd on Eggplant Ionic Balance
4.6. Effects of Cd on Eggplant HMP and MTP Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Phylogenetic Analysis on MTP and HMA Genes


References
- Srivastava, V.; Sarkar, A.; Singh, S.; Singh, P.; de Araujo, A.S.; Singh, R.P. Agroecological responses of heavy metal pollution with special emphasis on soil health and plant performances. Front. Environ. Sci. 2017, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M.C. Cadmium bioavailability, uptake, toxicity and detoxification in soil-plant system. Rev. Environ. Contam. Toxicol. 2017, 241, 73–137. [Google Scholar]
- Edelstein, M.; Ben-Hur, M. Heavy metals and metalloids: Sources, risks and strategies to reduce their accumulation in horticultural crops. Sci. Hortic. 2018, 234, 431–444. [Google Scholar] [CrossRef]
- Sohail, M.I.; Arif, M.; Rauf, A.; Rizwan, M.; Ali, S.; Saqib, M.; Zia-Ur-Rehman, M. Organic manures for cadmium tolerance and remediation. In Cadmium Tolerance in Plants: Agronomic, Molecular, Signaling, and Omic Approaches; Hasanuzzaman, M., Narasimha, M., Prasad, V., Nahar, K., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 19–67. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; Smolders, E.; Zhao, F.J.; Grant, C.; Montalvo, D. Chapter One—Managing cadmium in agricultural systems. Adv. Agron 2021, 166, 1–129. [Google Scholar] [CrossRef]
- Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Cadmium toxicity in plants. Braz. J. Plant Physiol 2005, 17, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Méndez, A.A.; Gallego, S.M. Oxidative post translational modifications of proteins related to cell cycle are involved in cadmium toxicity in wheat seedlings. Plant Sci. 2012, 196, 1–7. [Google Scholar] [CrossRef]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and plant development: An agony from seed to seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Vliet, L.; Peterson, C.; Hale, B. Cd accumulation in roots and shoots of durum wheat: The roles of transpiration rate and apoplastic bypass. J. Exp. Bot. 2007, 58, 2939–2947. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Plant, J.A.; Voulvoulis, N.; Oates, C.J.; Ihlenfeld, C. Cadmium levels in Europe: Implications for human health. Environ. Geochem. Health 2010, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Aarts, M.G.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jin, L.; Wang, X. Cadmium absorption and transportation pathways in plants. Int. J. Phytoremediation 2017, 19, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Chen, J.; Zou, W.; Meng, L.; Fan, X.; Xu, G.; Ye, G. Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice. Int. J. Mol. Sci. 2019, 20, 3417. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Gao, Y.; Tang, Y.; Wang, D.; Chen, X.; Yao, Y.; Guo, Y. Genome-Wide Identification, Comprehensive Gene Feature, Evolution, and Expression Analysis of Plant Metal Tolerance Proteins in Tobacco Under Heavy Metal Toxicity. Front. Genet. 2019, 10, 345. [Google Scholar] [CrossRef]
- Costa, G.; Morel, J.L. Water relations, gas exchange and amino acid content in Cd-treated lettuce. Plant Physiol. Biochem. 1994, 32, 561–570. [Google Scholar]
- Das, P.; Samantaray, S.; Rout, G.R. Studies on cadmium toxicity in plants: A review. Environ. Pollut. 1997, 98, 29–36. [Google Scholar] [CrossRef]
- Zhang, X.D.; Meng, J.G.; Zhao, K.X.; Chen, X.; Yang, Z.M. Annotation and characterization of Cd-responsive metal transporter genes in rapeseed (Brassica napus). Biometals 2018, 31, 107–121. [Google Scholar] [CrossRef]
- Sanità Di Toppi, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Sun, J.Y.; Shen, Z.G. Effects of Cd stress on photosynthetic characteristics and nutrient uptake of cabbages with different Cd-tolerance. Chin. J. Appl. Ecol. 2007, 18, 2605–2610. [Google Scholar]
- Zhang, J.; Sun, W.; Li, Z.; Liang, Y.; Song, A. Cadmium fate and tolerance in rice cultivars. Agron. Sustain. Dev. 2009, 29, 483–490. [Google Scholar] [CrossRef]
- Piotto, F.A.; Carvalho, M.E.A.; Souza, L.A.; Rabêlo, F.H.S.; Franco, M.R.; Batagin-Piotto, K.D.; Azevedo, R.A. Estimating tomato tolerance to heavy metal toxicity: Cadmium as study case. Environ. Sci. Pollut. Res. 2018, 25, 27535–27544. [Google Scholar] [CrossRef]
- Zhou, Q.; Guo, J.J.; He, C.T.; Shen, C.; Huang, Y.Y.; Chen, J.X.; Guo, J.H.; Yuan, J.G.; Yang, Z.Y. Comparative transcriptome analysis between low- and high-cadmium-accumulating genotypes of pakchoi (Brassica chinensis L.) in response to cadmium stress. Environ. Sci. Technol. 2016, 50, 6485–6494. [Google Scholar] [CrossRef]
- Guo, J.J.; Tan, X.; Fu, H.L.; Chen, J.X.; Lin, X.X.; Ma, Y.; Yang, Z.Y. Selection for Cd pollution-safe cultivars of Chinese kale (Brassica alboglabra L. H. Bailey) and biochemical mechanisms of the cultivar-dependent Cd accumulation involving in Cd subcellular distribution. J. Agric. Food. Chem. 2018, 66, 1923–1934. [Google Scholar] [CrossRef]
- Lester, R.N.; Hasan, S.M.Z. Origin and domestication of the brinjal eggplant, Solanum melongena, from S. incanum, in Africa and Asia. In Solanaceae III: Taxonomy, Chemistry, Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; The Royal Botanic Gardens, Kew: London, UK, 1991; pp. 369–388. [Google Scholar]
- Meyer, R.S.; Karol, K.G.; Little, D.P.; Nee, M.H.; Litt, A. Phylogeographic relationships among Asian eggplants and new perspectives on eggplant domestication. Mol. Phylogenetics Evol. 2012, 63, 685–701. [Google Scholar] [CrossRef]
- Cericola, F.; Portis, E.; Toppino, L.; Barchi, L.; Acciarri, N.; Ciriaci, T.; Sala, T.; Rotino, G.L.; Lanteri, S. The population structure and diversity of eggplant from Asia and the Mediterranean Basin. PLoS ONE 2013, 8, e73702. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Fu, H.L.; Liao, Q.; Huang, B.F.; Huang, Y.Y.; Xin, J.L. Selection for low-cadmium cultivars and cadmium subcellular distribution comparison between two selected cultivars of eggplant (Solanum melongena L.). Environ. Sci. Pollut. Res. 2021, 28, 57739–57750. [Google Scholar] [CrossRef]
- Dai, H.; Wei, S.; Twardowska, I.; Zhang, Q. In search of the exclusion/low-accumulation mechanisms: Cadmium uptake and accumulation from soil by cultivated (Solanum melongena L.) and wild eggplants (Solanum torvum L.). J. Clean. Prod. 2021, 323, 129141. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.; Zhang, G. Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Hakmaoui, A.; Ater, M.; Boka, K.; Baron, M. Copper and cadmium tolerance, uptake and effect on chloroplast ultrastructure. Studies on Salix purpurea and Phragmites australis. Z. Für Nat. C 2007, 62, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.; Illiano, A.; Carpentieri, A.; Spinelli, M.; Melchiorre, C.; Fontanarosa, C.; di Serio, M.; Amoresano, A. Quantification of Polyphenols and Metals in Chinese Tea Infusions by Mass Spectrometry. Foods 2020, 9, 835. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Hayat, K.; Zhang, D.; Zhou, P. Small structures with big impact: Multi-walled carbon nanotubes enhanced remediation efficiency in hyperaccumulator Solanum nigrum L. under cadmium and arsenic stress. Chemosphere 2021, 276, 130130. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Buschmann, C. Chlorophylls and carotenoids: Measurements and characterization by UV-Vis spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Hirakawa, H.; Shirasawa, K.; Miyatake, K.; Nunome, T.; Negoro, S.; Ohyama, A.; Yamaguchi, H.; Sato, S.; Isobe, S.; Tabata, S.; et al. Draft Genome Sequence of Eggplant (Solanum melongena L.): The Representative Solanum Species Indigenous to the Old World. DNA Res. 2014, 21, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Slater, G.S.C.; Birney, E. Automated generation of heuristics for biological sequence comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, S.A.; Krämer, U. The zinc homeostasis network of land plants. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1553–1567. [Google Scholar] [CrossRef]
- Dar, F.A.; Pirzadah, T.B.; Malik, B. Accumulation of Heavy Metals in Medicinal and Aromatic Plants. In Plant Micronutrients; Aftab, T., Hakeem, K.R., Eds.; Springer: Cham, Switzerland, 2020; pp. 113–127. [Google Scholar] [CrossRef]
- Gantasala, N.P.; Papolu, P.K.; Thakur, P.K.; Kamaraju, D.; Sreevathsa, R.; Rao, U. Selection and validation of reference genes for quantitative gene expression studies by real-time PCR in eggplant (Solanum melongena L). BMC Res. Notes 2013, 6, 312. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.V.; Ivanov, V.B. Physiological Aspects of Cadmium and Lead Toxic Effects on Higher Plants. Russ. J. Plant Physiol. 2001, 48, 523–544. [Google Scholar] [CrossRef]
- Kuriakose, S.V.; Prasad, M.N.V. Cadmium stress affects seed germination and seedling growth in Sorghum bicolor (L.) Moench by changing the activities of hydrolyzing enzymes. Plant Growth Regul. 2008, 54, 143–156. [Google Scholar] [CrossRef]
- Siddhu, G.; Sirohi, D.S.; Kashyap, K.; Khan, I.A.; Khan, M.A. Toxicity of cadmium on the growth and yield of Solanum melongena L. J. Environ. Biol. 2008, 29, 853–857. [Google Scholar] [PubMed]
- Qin, Q.; Li, X.; Zhuang, J.; Weng, L.; Liu, W.; Tai, P. Long-distance transport of cadmium from roots to leaves of Solanum melongena. Ecotoxicol. 2015, 24, 2224–2232. [Google Scholar] [CrossRef]
- Dube, B.K.; Sinha, P.; Shukla, K.; Chatterjee, C.; Pandey, V.K.; Rai, A.D. Involvement of excess cadmium on oxidative stress and other physiological parameters of eggplant. J. Plant Nutr. 2009, 32, 996–1004. [Google Scholar] [CrossRef]
- Neelima, P.; Reddy, K.J. Differential effect of cadmium and mercury on growth and metabolism of Solanum melongena L. seedlings. J. Environ. Biol. 2003, 24, 453–460. [Google Scholar]
- Ovečka, M.; Takáč, T. Managing heavy metal toxicity stress in plants: Biological and biotechnological tools. Biotechnol. Adv. 2014, 32, 73–86. [Google Scholar] [CrossRef]
- Youssef, M.A.; AbdEl-Gawad, A.M. Accumulation and Translocation of Heavy Metals in Eggplant (Solanum melongena L.) Grown in a Contaminated Soil. J. Environ. Chem. Eng. 2018, 3, 9–18. [Google Scholar]
- Michałowski, M.; Gołas, J. The contents of selected heavy metals of willow organs as an indicator of their use in sewage sludge utilization. Workb. Basic Probl. Agric. Sci. 2011, 477, 411–419. [Google Scholar]
- Lux, A.; Šottníková, A.; Opatrná, J.; Greger, M. Differences in structure of adventitious roots in Salix clones with contrasting characteristics of cadmium accumulation and sensitivity. Physiol. Plant. 2004, 120, 537–545. [Google Scholar] [CrossRef]
- Kang, W.; Bao, J.; Zheng, J.; Hu, H.; Du, J. Distribution and chemical forms of copper in the root cells of castor seedlings and their tolerance to copper phytotoxicity in hydroponic culture. Environ. Sci. Pollut. Res. Int. 2015, 22, 7726–7734. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Mechanisms to cope with arsenic or cadmium excess in plants. Curr. Plant Biol. 2009, 12, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E.; Prince, R.C.; Pickering, I.J.; Raskin, I. Mechanisms of cadmium mobility and accumulation in Indian mustard. Plant Physiol. 1995, 109, 1427–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.; Li, J.; Pan, X.; Jiang, H.; Sun, L.; Fu, Z. Photoperiod and temperature influence cadmium’s effects on photosynthesis-related gene transcription in Chlorella vulgaris. Ecotoxicol. Environ. Saf. 2010, 73, 1202–1206. [Google Scholar] [CrossRef] [PubMed]
- Lagriffoul, A.; Mocquot, B.; Vangronsveld, J.; Mench, M. Cadmium toxicity effects on growth, mineral and chlorophyll contents, and activities of stress related enzymes in Young maize plants (Zea mays L.). Plant Soil 1998, 200, 241–250. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Grajek, H.; Rydzyński, D.; Piotrowicz-Cieślak, A.; Herman, A.; Maciejczyk, M.; Wieczorek, Z. Cadmium ion-chlorophyll interaction–Examination of spectral properties and structure of the cadmium-chlorophyll complex and their relevance to photosynthesis inhibition. Chemosphere 2020, 261, 127434. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Gzyl, J.; Rucińska-Sobkowiak, R.; Arasimowicz-Jelonek, M.; Deckert, J. The new insights into cadmium sensing. Front. Plant Sci. 2014, 5, 245. [Google Scholar] [CrossRef]
- Hopkins, W.G.; Huüner, N.P. Introduction to Plant Physiology, 4th ed.; Wiley: New York, NY, USA, 1995. [Google Scholar]
- Hatata, M.M.; Abdel, A.E. Oxidative stress and antioxidant defense mechanisms in response to cadmium treatments. Am.-Eurasian J. Agric. Environ. Sci. 2008, 4, 655–669. [Google Scholar]
- Qin, S.; Liu, H.; Nie, Z.; Rengel, Z.; Gao, W.; Li, C.; Zhao, P. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere 2020, 30, 168–180. [Google Scholar] [CrossRef]
- Baxter, I.R.; Vitek, O.; Lahner, B.; Muthukumar, B.; Borghi, M.; Morrissey, J.; Guerinot, M.L.; Salt, D.E. The leaf ionome as a multivariable system to detect a plant’s physiological status. Proc. Natl. Acad. Sci. USA 2008, 105, 12081–12086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, I.R.; Ziegler, G.; Lahner, B.; Mickelbart, M.V.; Foley, R.; Danku, J.; Armstrong, P.; Salt, D.E.; Hoekenga, O.A. Single-kernel ionomic profiles are highly heritable indicators of genetic and environmental influences on elemental accumulation in maize grain (Zea mays). PLoS ONE 2014, 9, e87628. [Google Scholar] [CrossRef] [Green Version]
- Buescher, E.; Achberger, T.; Amusan, I.; Giannini, A.; Ochsenfeld, C.; Rus, A.; Lahner, B.; Hoekenga, O.; Yakubova, E.; Harper, J.F.; et al. Natural genetic variation in selected populations of Arabidopsis thaliana is associated with ionomic differences. PLoS ONE 2010, 5, e11081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danku, J.M.C.; Lahner, B.; Yakubova, E.; Salt, D.E. Large-scale plant ionomics. In Plant Mineral Nutrients. Methods in Molecular Biology (Methods and Protocols); Maathuis, F., Ed.; Humana Press: Totowa, NJ, USA, 2013; Volume 953, pp. 255–276. [Google Scholar] [CrossRef]
- Zhang, M.; Pinson, S.R.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corso, M.; de la Torre, V.S.G. Biomolecular approaches to understanding metal tolerance and hyperaccumulation in plants. Metallomics 2020, 12, 840–859. [Google Scholar] [CrossRef]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef]
- Siemianowski, O.; Barabasz, A.; Kendziorek, M.; Ruszczyńska, A.; Bulska, E.; Williams, L.E.; Antosiewicz, D.A. HMA4 expression in tobacco reduces Cd accumulation due to the induction of the apoplastic barrier. J. Exp. Bot. 2014, 65, 1125–1139. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yang, F.; Liu, J.; Xie, W.; Zhang, L.; Chen, Z.; Peng, Z.; Yongbin, O.; Yao, Y. Genome-wide identification of metal tolerance protein genes in Populus trichocarpa and their roles in response to various heavy metal stresses. Int. J. Mol. Sci. 2020, 21, 1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhiguo, E.; Tingting, L.I.; Chen, C.H.E.N.; Lei, W.A.N.G. Genome-wide survey and expression analysis of P1B-ATPases in rice, maize and sorghum. Rice Sci. 2018, 25, 208–217. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment | Tolerance Index [%] | sd |
|---|---|---|---|
| BN | Cd 5 | 90.63 | 4.32 |
| Cd 10 | 71.25 | 3.35 | |
| Cd 25 | 60.9 | 5.54 | |
| Cd 50 | 58.35 | 2.38 | |
| CV | Cd 5 | 88.23 | 6.98 |
| Cd 10 | 51.76 | 5.85 | |
| Cd 25 | 50.89 | 3.99 | |
| Cd 50 | 48.76 | 4.01 | |
| PQ | Cd 5 | 60.08 | 6.87 |
| Cd 10 | 35.71 | 5.72 | |
| Cd 25 | 20.15 | 4.88 | |
| Cd 50 | 4.89 | 1.32 | |
| RB | Cd 5 | 100 | 4.84 |
| Cd 10 | 55 | 3.83 | |
| Cd 25 | 21 | 2.15 | |
| Cd 50 | 18.74 | 3.44 | |
| VF | Cd 5 | 73.38 | 7.02 |
| Cd 10 | 48.38 | 3.66 | |
| Cd 25 | 39.12 | 2.83 | |
| Cd 50 | 35.48 | 2.46 | |
| VL | Cd 5 | 60.07 | 7.10 |
| Cd 10 | 28.12 | 6.77 | |
| Cd 25 | 29.31 | 5.44 | |
| Cd 50 | 27.96 | 4.32 |
| Cultivar | Bioaccumulation Index | sd |
|---|---|---|
| BN | 2.61 | 0.16 |
| CV | 1.87 | 0.09 |
| PQ | 1.79 | 0.29 |
| RB | 1.18 | 0.20 |
| VF | 2.47 | 0.23 |
| VL | 2.10 | 0.41 |
| Treatment | Chl a (µg g−1 Fw) | Sd | Chl b (µg g−1 Fw) | Sd | Chl tot (µg g−1 Fw) | Sd | Carotenoid (µg g−1 Fw) | Sd |
|---|---|---|---|---|---|---|---|---|
| Cd 0 | 660.18 a | 182.64 | 241.20 a | 60.37 | 901.38 a | 242.58 | 622.54 a | 184.32 |
| Cd 5 | 548.94 b | 213.51 | 201.75 a | 85.00 | 750.69 a | 292.80 | 643.88 a | 201.88 |
| Cd 10 | 282.80 c | 61.18 | 175.12 b | 65.26 | 457.93 b | 125.53 | 721.58 a | 229.03 |
| Cd 25 | 183.22 c | 17.63 | 147.47 b | 52.60 | 330.69 b | 70.04 | 830.39 b | 279.56 |
| Cd 50 | 89.08 c | 48.20 | 166.74 b | 177.37 | 255.82 b | 187.56 | 1006.52 b | 277.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippone, E.; Tranchida-Lombardo, V.; Vitiello, A.; Ruiu, F.; Di Salvatore, M.; Galise, T.R.; Laccetti, L.; Amoresano, A.; D’Ambrosio, G.; Frusciante, L.; et al. Evaluation of Cadmium Effects on Six Solanum melongena L. Cultivars from the Mediterranean Basin. Agriculture 2022, 12, 1059. https://doi.org/10.3390/agriculture12071059
Filippone E, Tranchida-Lombardo V, Vitiello A, Ruiu F, Di Salvatore M, Galise TR, Laccetti L, Amoresano A, D’Ambrosio G, Frusciante L, et al. Evaluation of Cadmium Effects on Six Solanum melongena L. Cultivars from the Mediterranean Basin. Agriculture. 2022; 12(7):1059. https://doi.org/10.3390/agriculture12071059
Chicago/Turabian StyleFilippone, Edgardo, Valentina Tranchida-Lombardo, Alessia Vitiello, Fabrizio Ruiu, Mina Di Salvatore, Teresa Rosa Galise, Lucrezia Laccetti, Angela Amoresano, Gennaro D’Ambrosio, Luigi Frusciante, and et al. 2022. "Evaluation of Cadmium Effects on Six Solanum melongena L. Cultivars from the Mediterranean Basin" Agriculture 12, no. 7: 1059. https://doi.org/10.3390/agriculture12071059