
Carbonated Irrigation Assessment of Grapevine Growth, Nutrient Absorption, and Sugar Accumulation in a Tempranillo (Vitis vinifera L.) Vineyard

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1 in Bare Soil Pots: Effects of Acidified Water Irrigation on Soil pH and Mineral Composition
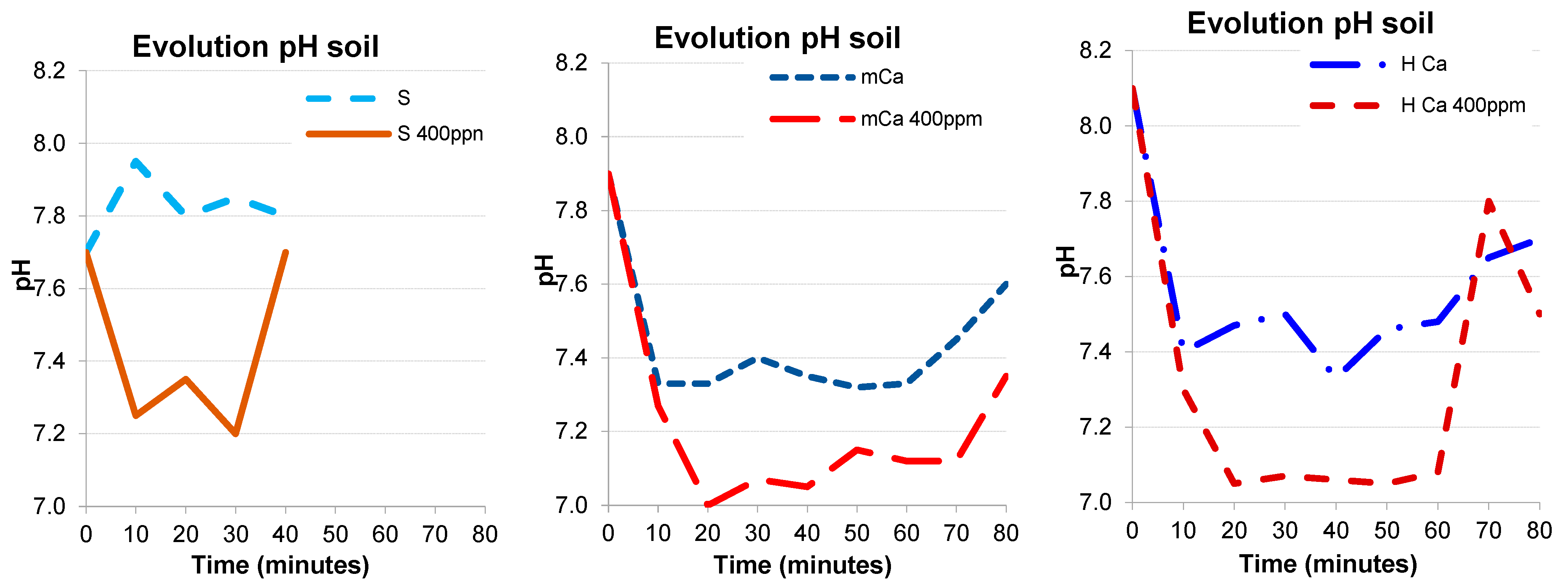
2.2. Experiment 2 in Planted cv. Tempranillo Pots: Effects of Acidified Water Irrigation with 3 Different Limestone Composition
2.3. Soil Mineral Composition
2.4. Plant Vegetative Weight
2.5. Leaf and Shoot Composition
2.6. Berry Composition
2.7. Data Analysis
3. Results
3.1. Experiment 1: Effects of Acidified Water Irrigation on Soil pH and Mineral Composition
3.2. Experiment 2: Effects of Irrigation with Acidified Water at 400 ppm CO2 on Soil and Grape Plants in Pots with 3 Different Soils
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brancadoro, L.; Vanoli, M.; Zoocchi, G. Adaptive responses to high concentration of HCO3 in Cabernet Sauvingnon. Acta Hort. 1996, 427, 75–81. [Google Scholar] [CrossRef]
- López-Rayo, S.; Sanchis-Pérez, I.; Ferreira, C.; Lucena, J. [S,S]-EDDS/Fe: A new chelate for the environmentally sustainable correction of iron chlorosis in calcareous soil. Sci. Total Environ. 2019, 647, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Dell’orto, M.; Brancadoro, L.; Scienza, A.; Zocchi, G. Use of biochemical parameters to select grapevine varieties resistant to iron-chlorosis. J. Plant Nutr. 2000, 23, 1767–1775. [Google Scholar] [CrossRef]
- Dell’orto, M.; De Nisi, P.; Pontiggia, A.; Zocchi, G. Fe deficiency responses in Parietaria diffusa: A calcicole plant. J. Plant Nutr. 2003, 26, 2057–2068. [Google Scholar] [CrossRef]
- Mengel, K. Iron availability in plant tissues-iron chlorosis on calcareous soils. Plant Soil 1994, 165, 275–283. [Google Scholar] [CrossRef]
- Nikolic, M.; Römhel, V. The Dynamics of Iron in the Leaf Apoplast. Significance for the Iron Nutrition of Plants. In The Apoplast of Higher Plants: Compartment of Storage, Transport, and Reactions. The Significance of the Apoplast for the Mineral Nutrition of Higher Plants; Sattelmacher, B., Horst, W.J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 353–371. [Google Scholar]
- Bavaresco, L.; Bertamini, M.; Iacono, F. Lime-induced chlorosis and physiological responses in grapevine (Vitis vinifera L. cv. Pinot blanc) leaves. Vitis 2006, 45, 45–46. [Google Scholar]
- Römheld, V. The chlorosis paradox: Fe inactivation as a secondary event in chlorotic leaves of grapevine. J. Plant Nutr. 2000, 23, 1629–1643. [Google Scholar] [CrossRef]
- Rombolà, A.D.; Tagliavini, M. Iron Nutrition of Fruit Tree Crops. In Nutrition in Plants and Rhizospheric Microorganisms; Abadía, J., Barton, L., Eds.; Springer: Berlin, Germany, 2006; pp. 61–83. [Google Scholar]
- Tangolar, S.G.; Ünlü, G.; Tangolar, S.; Daşgan, Y.; Yilmaz, N. Use of in vitro method to evaluate some grapevine varieties for tolerance and susceptibility to sodium bicarbonate-induced chlorosis. Vitr. Cell. Dev. Biol.-Plant 2008, 44, 233–237. [Google Scholar] [CrossRef]
- Cambrollé, J.; García, J.L.; Ocete, R.; Figueroa, M.E.; Cantos, M. Evaluating tolerance to calcareous soils in Vitis vinifera ssp. sylvestris. Plant Soil 2015, 396, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Alcántara, E.; Cordeiro, A.M.; Barranco, D. Selection of olive varieties for tolerance to iron chlorosis. J. Plant Physiol. 2003, 160, 1467–1472. [Google Scholar] [CrossRef] [Green Version]
- Ksouri, R.; Gharsalli, M.; Lachaal, M. Physiological responses of Tunisian grapevine varieties to bicarbonate-induced iron deficiency. J. Plant Physiol. 2005, 162, 335–341. [Google Scholar] [CrossRef]
- Covarrubias, J.I.; Rombolà, A.D. Physiological and biochemical responses of the iron chlorosis tolerant grapevine rootstock 140 Ruggeri to iron deficiency and bicarbonate. Plant Soil 2013, 370, 305–315. [Google Scholar] [CrossRef]
- Mukherjee, I.; Campbell, N.H.; Ash, J.S.; Connolly, E. Expression profiling of the Arabidopsis ferric chelate reductase (FRO) gene family reveals differential regulation by iron and car. Plant Sci. 2006, 223, 1178–1190. [Google Scholar] [CrossRef]
- Díaz, I.; del Campillo, M.C.; Cantos, M.; Torrent, J. Iron deficiency symptoms in grapevine as affected by the iron oxide and carbonate contents of model substrates. Plant Soil 2009, 322, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, J.I.; Pisi, A.; Rombolà, A.D. Evaluation of sustainable management techniques for preventing iron chlorosis in the grapevine. Aust. J. Grape Wine Res. 2014, 20, 149–159. [Google Scholar] [CrossRef]
- Jiménez, S.; Ollat, N.; Debordec, C.; Maucourtc, M.; Rellán-Álvareze, R.; Morenoa, M.A.; Gogorcena, Y. Metabolic response in roots of Prunus rootstocks submitted to iron chlorosis. J. Plant Physiol. 2011, 168, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Bavaresco, L.; van Zeller de Macedo Basto Gonçalves, M.A.; Civardi, S.; Gatti, M.; Ferrari, F. Effects of traditional and new methods on overcoming lime-induced chlorosis of grapevine (Vitis vinifera L. cv. Pinot blanc) leaves. Am. J. Enol. Vitic. 2010, 61, 186–190. [Google Scholar]
- Sánchez, R.; González, M.R.; Ilanova, M.; Rodríguez-Nogales, J.M.; Martín, P. Aroma composition of Tempranillo grapes as affected by iron deficiency chlorosis and vine water status. Sci. Agric. 2021, 78, e20190112. [Google Scholar] [CrossRef]
- Hunter, J.J.; Skrivan, R.; Ruffner, H.P. Diurnal and seasonal physiological changes in leaves of Vitis vinifera L.: CO2 assimilation rates, sugar levels and sucrolytic enzyme activity. Vitis 1994, 33, 184–195. [Google Scholar]
- Schulze, E.D. Whole-plant responses to drought. Aust. J. Plant Physiol. 1986, 13, 127–141. [Google Scholar] [CrossRef]
- Piagnani, C.; De Nisi, P.; Espen, L.; Zocchi, G. Adaptive responses to iron-deficiency in callus cultures of two cultivars of Vitis spp. J. Plant Physiol. 2003, 160, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Lucena, J.J. Synthetic Iron Chelates to correct Iron Deficiency in Plants. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Springer: Dordrecht, The Netherlands, 2006; pp. 103–128. [Google Scholar]
- López-Rayo, S.; Di Foggia, M.; Bombai, G.; Yunta, F.; Rodrigues Moreira, E.; Filippini, G.; Pisi, A.; Rombolà, A.D. Blood-derived compounds can efficiently prevent iron deficiency in the grapevine. Aust. J. Grape Wine Res. 2015, 21, 135–142. [Google Scholar] [CrossRef]
- Peet, M.M.; Willits, D.H. Greenhouse CO2 enrichment. Effects of increasing concentration or duration of enrichment on cucumber yield. J. Am. Hort. Sci. Soc. 1987, 112, 236–241. [Google Scholar]
- Ibrahimm, A.A. Response of plant to irrigation with CO2-enriched water. Acta Hortic. 1992, 323, 205–214. [Google Scholar] [CrossRef]
- Mauney, J.R.; Hendrix, D.L. Responses of glasshouse grown cotton to irrigation with carbon dioxide-saturated water. Crop Sci. 1988, 28, 835–838. [Google Scholar] [CrossRef] [Green Version]
- Mannini, P.; Gallina, D. Effetti dellírrigazione della fragola con acqua carbonicata. Riv. Irr. Dren. 1995, 42, 18–21. [Google Scholar]
- Arienzo, M.; Basile, G.; D’Andria, R.; Magliulo, V.; Zena, A. Irrigazione con acqua carbonicata e disponibilità di elementi nutritivi. Prova su coltura di fragola. Agrochimica 1995, 39, 61–72. [Google Scholar]
- Hartz, T.K.; Holt, D.B. Root-zone carbon dioxide enrichment in field does not improve tomato or cucumber yield. HortScience 1991, 26, 1423. [Google Scholar] [CrossRef] [Green Version]
- Lampreave, M. Fisiologia de L’absorció en L’ecosistema Vitícola Sòl-Planta-Atmosfera i Repercussions en la Qualitat del Raïm. Ph.D. Thesis, Department of Bioquímica i Biotecnologia, URV, Tarragona, Spain, 2002. [Google Scholar]
- Doorenbos, J.; Pruitt, W.O. Guidelines for Predicting Crop Water Requirements, 2nd ed.; lrrigation and Drainage, Paper 24; Food and Agriculture Organization: Rome, Italy, 1984; 144p. [Google Scholar]
- AOAC. International, Official Methods of Analysis; AOAC: Rockville, MD, USA, 1990. [Google Scholar]
- Olsen, R.S.; Cole, V.C.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; 4 Circular 939; USDA: Washinghton, DC, USA, 1954. [Google Scholar]
- Davies, D.H. Carotenoids. Chemistry and Biochemistry of Algal Pigments; Goodwin, T.W., Ed.; Academic Press: New York, NY, USA, 1976; Volume 2, pp. 38–165. [Google Scholar]
- Buysse, J.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Barham, D.; Trinder, P. An improved color reagent for the determination of blood glucose by the oxidase system. Analyst 1972, 97, 142–145. [Google Scholar] [CrossRef]
- OIV. Recueil des Méthodes Internationales D’analyse des Vins et des Moûts; Office International de la vigne et du vin: Paris, France, 1990. [Google Scholar]
- Alvino, A.; D’Andria, R.; Cerio, L.; Mori, M.; Sorrentino, G. Physiological response of Gladiolus to carbonated water application under two water regimes. Adv. Hort. Sci. 1997, 12, 145–152. [Google Scholar]
- Jalali, M.; Naderi, E. The impact of acid rain on phosphorus leaching from a sandy loam calcareous soil of western Iran. Environ. Earth Sci. 2012, 66, 311–317. [Google Scholar] [CrossRef]
- Hahn, G.; Marschner, H. Cation concentrations of short roots of Norway spruce as affected by acid irrigation and liming. Plant Soil 1998, 199, 23–27. [Google Scholar] [CrossRef]
- Covarrubias, J.I.; Retamales, C.; Donnini, S.; Rombolà, D.; Pastenes, C. Contrasting physiological responses to iron deficiency in Cabernet Sauvignon grapevines grafted on two rootstocks. Sci. Hortic. 2016, 199, 1–8. [Google Scholar] [CrossRef]
- Vannozzi, A.; Donnini, S.; Vigani, G.; Corso, M.; Valle, G.; Vitulo, N.; Bonghi, C.; Zocchi, G.; Lucchin, M. Transcriptional Characterization of a Widely-Used Grapevine Rootstock Genotype under Different Iron-Limited Conditions. Plant Sci. 2017, 7, 1994. [Google Scholar] [CrossRef] [Green Version]
- D’Andria, R.; Magliulo, V.; Maggio, A.; Basile, G.; Arienzo, M. Soil and plant nutrient modifications in response to irrigation with CO2 enriched water on tomato. Act. Hortic. 1993, 335, 557–562. [Google Scholar] [CrossRef]
- Aguilera, C.; Checa, A.B. Efectos de la fertirrigación carbónica en tomate. Frutic. Hortic. Floric. 2000, 227, 747–755. [Google Scholar]
- Baron, J.J.; Gorski, S.F. Response of eggplant to a root environment enriched with CO2. Hortscience 1986, 21, 496–498. [Google Scholar]
- Karimi, R.; Salimi, F. Iron-chlorosis tolerance screening of 12 commercial grapevine (Vitis vinifera L.) cultivars based on phytochemical indices. Sci. Hortic. 2021, 283, 110111. [Google Scholar] [CrossRef]
- Shahsavandi, F.; Eshghi, S.; Gharaghani, A.; Ghasemi-Fasaei, R.; Jafarinia, M. Effects of bicarbonate induced iron chlorosis on photosynthesis apparatus in grapevine. Sci. Hortic. 2020, 270, 109427. [Google Scholar] [CrossRef]
- Hunter, J.J.; Visser, J.H. The effect of partial defoliation, leaf position and developmental stage of the vine on leaf chlorophyll concentration in relation to the photosynthetic activity and light intensity in the canopy of Vitis vinifera L. cv. Cabernet Sauvignon. S. Afr. J. Enol. Vitic. 1989, 10, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Cartechini, A.; Palliotti, A. Effect of shanding on vine morphology and productivity and leaf gas exchange characteristics in grapevines in the fields. Am. J. Enol. Vitic. 1995, 46, 227–234. [Google Scholar]
- Gruber, B.; Kosegarten, H. Depressed growth of non-chlorotic vine grown in calcareous soil is an iron deficiency symptom prior to leaf chlorosis. J. Plant Nutr. Soil Sci. Z. Pflanzenernahr. Bodenkd. 2002, 165, 111–117. [Google Scholar] [CrossRef]
- Shahsavandi, F.; Eshghi, S. Effects of bicarbonate and Fe sources on vegetative growth and physiological traits of four grapevine cultivars. Commun. Soil Sci. Plant Anal. 2021, 52, 2401–2413. [Google Scholar] [CrossRef]
- Cinelli, F.; Fisichella, M.; Mulero, R. Morpho-physiological approaches to investigate lime-induced chlorosis in deciduous fruit tree species. J. Plant Nutr. 2003, 26, 2277–2294. [Google Scholar] [CrossRef]
- Pestana, M.; De Varennes, A.; Abadía, J.; Araújo, F. Differential tolerance to iron deficiency of citrus rootstock grown in nutrient solution. Sci. Hortic. 2005, 104, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Charlson, D.V.; Cianzio, S.R.; Shoemaker, R.C. Associating SSR markers with soybean resistance to iron deficiency chlorosis. J. Plant Nutr. 2003, 26, 2267–2276. [Google Scholar] [CrossRef]
- Hanson, A.D.; Hitz, W.D. Metabolic responses of mesophytes of plant water deficits. Annu. Rev. Plant Physiol. 1982, 33, 163–203. [Google Scholar] [CrossRef]
- Huber, S.C.; Rogers, H.M.; Mowry, F.L. Effects of water stress on photosynthesis and carbon partitioning in soybean (Glycine max L. Merr) plants grown in the field at different CO2 levels. Plant Physiol. 1984, 76, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.C.; Stern, W.R.; Evans, P. Water Relations and Osmotic Adjustment of leaves and Roots of Lupins in Response to Water Deficits. Crop Sci. 1978, 27, 977–983. [Google Scholar] [CrossRef]
- Sanchez, R.; Gonzalez, M.R.; Martin, P. Relationships between chlorophyll content of vine leaves, predawn leaf water potential at veraison, and chemical and sensory attributes of wine. J. Sci. Food Agric. 2020, 100, 5251–5259. [Google Scholar] [CrossRef]
- Morgan, J.M. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Hunter, J.J.; Ruffner, H.P. Assimilate transport in grapevines-effect of phloem disruption. Aust. J. Grape Wine Res. 2001, 7, 118–126. [Google Scholar] [CrossRef]
- Patakas, A. Changes in the solutes contributing to osmotic potential during leaf ontogeny in grapevine leave. Am. J. Enol. Vitic. 2000, 51, 223–225. [Google Scholar]
- Salisbury, F.; Ross, C. Fisiología de las Plantas. Bioquímica Vegetal; Thomson, P., Ed.; Paraninfo: Madrid, Spain, 2006; Volume 2, ISBN 978-84-283-2718. [Google Scholar]
- Stolwijk, J.A.J.; Thimann, K.V. On the uptake of carbon dioxide and bicarbonate by roots, and its influence on growth. Plant Physiol. 1957, 32, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Bergquist, N.O. Absorption of carbon dioxide by plant roots. Bot. Not. 1964, 117, 249–261. [Google Scholar]
- Arteca, R.N.; Poovaiah, B.W. Absorption of 14CO2 by potato roots and its subsequent translocation. J. Am. Soc. Hortic. Sci. 1982, 107, 398–401. [Google Scholar]
- Gonzalo, M.J.; Moreno, M.A.; Gogorcena, Y. Physiological responses and differential gene expression under iron deficiency conditions. J. Plant Physiol. 2011, 168, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Brancadoro, L.; Rabotti, G.; Scienza, A.; Zoocchi, G. Mechanisms of Fe-efficiency in roots of Vitis spp. in response to iron deficiency stress. Plant Soil 1995, 171, 229–234. [Google Scholar] [CrossRef]
- López-Millán, A.F.; Morales, F.; Gogorcena, Y.; Abadía, A.; Abadía, J. Metabolic responses in iron-deficient tomato plants. J. Plant Physiol. 2009, 166, 375–384. [Google Scholar] [CrossRef]
- Jiménez, S.; Pinochet, J.; Abadía, A.; Moreno, M.A.; Gogorcena, Y. Tolerance Response to Iron Chlorosis of Prunus Selections as Rootstocks. HortScience 2008, 43, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Abadía, J.; López-Millán, A.F.; Rombolà, A.; Abadía, A. Organic acids and Fe deficiency: A review. Plant Soil 2002, 241, 75–86. [Google Scholar] [CrossRef]
- Jiménez, S.; Gogorcena, Y.; Hévin, C.; Rombolà, A.D.; Ollat, N. Nitrogen nutrition influences some biochemical responses to iron deficiency in tolerant and sensitive genotypes of Vitis. Plant Soil 2007, 290, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Hahn, G.; Marschner, H. Effect of acid irrigation and liming on root growth of Norway spruce. Plant Soil 1998, 199, 11–22. [Google Scholar] [CrossRef]
- Marastoni, L.; Lucini, L.; Varanini, Z. Changes in physiological activities and root exudation profile of two grapevine rootstocks reveal common and specific strategies for Fe acquisition. Sci. Rep. 2020, 10, 18839. [Google Scholar] [CrossRef]
- Winkler, A.J.; Williams, W.O. Starch and sugars of Vitis vinifera. Plant Physiol. 1945, 20, 412–432. [Google Scholar] [CrossRef] [Green Version]
- Wees, D.; Stewart, K. The influence of bicarbonate enrichment and aeration on dissolved carbon dioxide and oxygen in NFT nutrient solutions used for lettuce production. Soil. Cult. 1987, 3, 50–62. [Google Scholar]
- Arteca, R.N.; Poovaiah, B.W. Changes in phosphoenolpyruvate carboxylase and ribulose-1,5-bisphospahte carboxylase in Solanum tuberosum L. as affected by root-zone application of CO2. HortScience 1982, 17, 396–398. [Google Scholar]
- Wen-Hsin, W.; Lu, J.; Jones, A.R.; Mortley, G.; Loretan, P.A.; Bonsi, C.K.; Hill, W. Proximate composition, amino acid profile, fatty acid composition, and mineral content of peanut seeds hydroponically grown at elevated CO2 levels. J. Agric. Food Chem. 1997, 45, 3863–3866. [Google Scholar]
- Nakayama, F.S.; Bucks, D.A. Using subsurface trickle system for carbon dioxide enrichment. In Proceedings of the 15th National Agricultural Plastics Congress, Tucson, AZ, USA, 13–17 April 1980; Jensen, M.H., Oebker, N.F., Eds.; National Agricultural Plastics Association: Manchester, MO, USA; pp. 13–18. [Google Scholar]


{kind=link}
| Irrigation Treatments | Water pH | Preparation of CO2 Solution | Calculated CO2 Gas Applied per Pot (CO2 g/pot) |
|---|---|---|---|
| Control CO2 at 0 ppm | 7.5 | No CO2 added (regular tap water) | 0 |
| High: CO2 at 800 ppm | 6.0 | CO2 gas dissolved under 1 atmosphere of pressure during 15 min in 35 L of tap water | 37.52 |
| Medium: CO2 at 400 ppm | 6.5 | 1 L of 800 ppm CO2 solution, added to 1.75 L tap water | 17.76 |
| Low: CO2 at 200 ppm | 7.0 | 250 mL of 800 ppm CO2 solution, added to 2.5 L tap water | 8.88 |
| Treatment/Type of Soil | pH | Limestone % | CaCO3 % |
|---|---|---|---|
| Neutral calcareous soil (Control) | 7.71 (±0.02) | 0.00 (±0.0) | 11.61 (±0.1) |
| Medium calcareous clayey soil | 7.93 (±0.03) | 10.41 (±0.1) | 32.63 (±0.2) |
| Highly calcareous clayey soil | 8.17 (±0.01) | 12.81 (±0.1) | 43.50 (±0.1) |
| Treatment Bare Soil Pots/CO2 Doses | pH | CaCO3 (%) | Ca (meq/100 g Soil) | Mg (meq/100 g Soil) | ||||
|---|---|---|---|---|---|---|---|---|
| Control CO2 at 0 ppm pH 7.5 | 8.08 ± 1.15 | ns | 24.0 ± 2.61 | ab | 88.7 ± 9.75 | b | 4.6 ± 0.23 | a |
| Low: CO2 at 200 ppm pH 7 | 8.1 ± 1.09 | ns | 24.9 ± 2.35 | a | 110.5 ± 8.37 | b | 3.5 ± 0.15 | a |
| Medium: CO2 at 400 ppm pH 6.5 | 8.03 ± 0.98 | ns | 24.9 ± 2.87 | a | 106 ± 11.51 | b | 1.4 ± 0.08 | b |
| High: CO2 at 800 ppm pH 6 | 7.94 ± 1.06 | ns | 20.9 ± 2.05 | b | 283.3 ± 9.44 | a | 4.8 ± 0.31 | a |
| Treatment Bare Soil Pots/CO2 Doses | p (ppm) | Fe (ppm) | Mn (ppm) | Zn (ppm) | ||||
| Control CO2 at 0 ppm pH 7.5 | 4.4 ± 0.36 | b | 1.2 ± 0.12 | b | 3.8 ± 1.01 | c | 3.0 ± 0.23 | b |
| Low: CO2 at 200 ppm pH 7 | 7.4 ± 0.89 | ab | 1.5 ± 0.25 | b | 5.2 ± 1.24 | b | 2.9 ± 0.15 | b |
| Medium: CO2 at 400 ppm pH 6.5 | 9.9 ± 1.74 | a | 4.1 ± 1.05 | a | 8.6 ± 0.78 | a | 3.9 ± 0.08 | a |
| High: CO2 at 800 ppm pH 6 | 6.6 ± 0.45 | b | 2.1 ± 0.39 | b | 4.4 ± 0.66 | bc | 2.2 ± 0.31 | b |
| Type of Soil | Irrigation Treatments | Fe (mg/kg Soil) | Zn (mg/kg Soil) | Mn (mg/kg Soil) | Cu (mg/kg Soil) | Carbonates (%) |
|---|---|---|---|---|---|---|
| Schist Neutral Soil | Control 0 ppm CO2 | 1.88 ± 0.75 | 1.90 ± 0.24 b | 4.67 ± 0.23 b | 0.34 ± 0.03 b | 28.42 ± 3.23 b |
| Schist Neutral Soil | 400 ppm CO2 | 1.62 ± 0.52 | 4.82 ± 0.0 a | 7.54 ± 3.63 a | 0.65 ± 0.02 a | 42.29 ± 4.63 a |
| Medium calc-clay | Control | 6.20 ± 0.1 a | 1.75 ± 0.06 b | 2.77 ± 0.27 | 0.87 ± 0.09 b | 33.58 ± 3.27 |
| Medium calc-clay | 400 ppm CO2 | 5.08 ± 0.19 b | 2.54 ± 0.17 a | 2.73 ± 0.23 | 1.33 ± 0.09 a | 37.83 ± 2.23 |
| Highly calcareous clay | Control | 4.08 ± 0.2 b | 2.38 ± 0.41 b | 2.36 ± 0.49 b | 1.17 ± 0.16 | 36.79 ± 3.49 |
| Highly calcareous clay | 400 ppm CO2 | 5.34 ± 0.83 a | 5.34 ± 0.36 a | 5.13 ± 0.19 a | 1.01 ± 0.15 | 33.29 ± 2.19 |
| FIRST YEAR | |||||
|---|---|---|---|---|---|
| Type of Soil | Irrigation Treatments | Chlorophyll (mg/g d.w.) | Fe (µg/kg d.w.) | Mn (mg/g d.w.) | Mg (mg/g d.w.) |
| Schist Neutral Soil | Control 0 ppm CO2 | 0.669 ± 0.03 b | 68.83 ± 8.46 | 68.94 ± 12.28 | 2.97 ± 0.25 |
| Schist Neutral Soil | 400 ppm CO2 | 1.074 ± 0.02 a | 83.01 ± 17.91 | 64.02 ± 7.11 | 1.66 ± 0.39 |
| Medium calc-clay | Control 0 ppm CO2 | 0.881 ± 0.04 | 99.60 ± 21.43 a | 64.19 ± 11.61 | 4.66 ± 0.68 a |
| Medium calc-clay | 400 ppm CO2 | 0.916 ± 0.05 | 65.77 ± 4.58 b | 55.44 ± 9.39 | 1.75 ± 0.50 b |
| Highly calcareous clay | Control 0 ppm CO2 | 0.732 ± 0.07 b | 95.67 ± 8.33 b | 29.39 ± 2.86 b | 5.03 ± 1.06 |
| Highly calcareous clay | 400 ppm CO2 | 1.013 ± 0.01 a | 193.36 ± 0.31 a | 54.75 ± 3.91 a | 3.66 ± 0.12 |
| SECOND YEAR | |||||
| Type of Soil | Irrigation Treatments | Chlorophyll (mg/g d.w.) | Fe (πg/kg d.w.) | Mn (mg/g d.w.) | Mg (mg/g d.w.) |
| Schist Neutral Soil | Control 0 ppm CO2 | 1.467 ± 0.03 b | 38.12 ± 4.36 a | 88.89 ± 2.37 a | 3.67 ± 0.27 a |
| Schist Neutral Soil | 400 ppm CO2 | 2.033 ± 0.08 a | 28.49 ± 3.16 b | 75.32 ± 4.95 b | 2.69 ± 0.12 b |
| Medium calc-clay | Control 0 ppm CO2 | 1.351 ± 0.11 | 31.61 ± 6.32 | 47.14 ± 4.19 a | 3.43 ± 0.17 |
| Medium calc-clay | 400 ppm CO2 | 1.574 ± 0.09 | 29.49 ± 4.48 | 35.29 ± 3.39 b | 3.44 ± 0.16 |
| Highly calcareous clay | Control 0 ppm CO2 | 1.254 ± 0.17 b | 22.94 ± 0.92 b | 48.12 ± 1.68 b | 3.81 ± 0.09 b |
| Highly calcareous clay | 400 ppm CO2 | 2.007 ± 0.10 a | 26.60 ± 1.35 a | 61.41 ± 1.12 a | 4.64 ± 0.13 a |
| THIRD YEAR | |||||
| Type of Soil | Irrigation Treatments | Chlorophyll (mg/g d.w.) | |||
| Schist Neutral Soil | Control 0 ppm CO2 | 2.49 ± 0.3 b | |||
| Schist Neutral Soil | 400 ppm CO2 | 3.46 ± 0.8 a | |||
| Medium calc-clay | Control 0 ppm CO2 | 2.29 ± 0.11 | |||
| Medium calc-clay | 400 ppm CO2 | 2.68 ± 0.19 | |||
| Highly calcareous clay | Control 0 ppm CO2 | 2.13 ± 0.17 b | |||
| Highly calcareous clay | 400 ppm CO2 | 3.41 ± 0.10 a | |||
| Type of Soil | Irrigation Treatments | Sucrose (mg/g d.w.) | Glucose (mg/g d.w.) | Fructose (mg/g d.w.) | Starch (mg/g d.w.) |
|---|---|---|---|---|---|
| Schist Neutral Soil | Control 0 ppm CO2 | 146.3 ± 1.93 b | 31.9 ± 8.81 b | 27.2 ± 6.9 b | 23.22 ± 4.84 a |
| Schist Neutral Soil | 400 ppm CO2 | 266.5 ± 26.2 a | 42.8 ± 6.32 a | 38.5 ± 5.4 a | 13.01 ± 2.53 b |
| Medium calc-clay | Control 0 ppm CO2 | 207.9 ± 31.8 b | 19.9 ± 4.73 b | 22.3 ± 3.8 b | 6.66 ± 1.09 b |
| Medium calc-clay | 400 ppm CO2 | 342.9 ± 45.1 a | 54.2 ± 3.61 a | 39.2 ± 0.5 a | 10.05 ± 1.75 a |
| Highly calcareous clay | Control 0 ppm CO2 | 194.4 ± 24.9 a | 41.7 ± 3.44 b | 14.3 ± 2.2 b | 3.52 ± 0.73 b |
| Highly calcareous clay | 400 ppm CO2 | 49.3 ± 17.6 b | 65.6 ± 10.9 a | 41.4 ± 5.3 a | 10.82 ± 1.69 a |
| Type of Soil | Irrigation Treatments | Malic (mg/g d.w.) V | Malic (mg/g d.w.) M | Malic (mg/g d.w.) H | Tartaric (mg/g d.w.) V | Tartaric (mg/g d.w.) M | Tartaric (mg/g d.w.) H |
|---|---|---|---|---|---|---|---|
| Schist Neutral Soil | Control 0 ppm CO2 | 18.35 ± 1.1 | 19.13 ± 1.2 | 17.2 ± 0.7 b | 141.7 ± 13.1 | 163.4 ± 14.6 b | 151.1 ± 6.91 b |
| Schist Neutral Soil | 400 ppm CO2 | 19.74 ± 1.9 | 18.06 ± 1.0 | 22.7 ± 0.4 a | 204.8 ± 14.5 | 336.3 ± 64.0 a | 174.5 ± 15.4 a |
| Medium calc-clay | Control 0 ppm CO2 | 20.99 ± 3.8 | 20.73 ± 1.4 | 17.5 ± 0.9 b | 197.0 ± 39.7 | 366.3 ± 39.6 | 154.8 ± 4.1 b |
| Medium calc-clay | 400 ppm CO2 | 15.54 ± 1.5 | 19.94 ± 0.6 | 24.8 ± 1.1 a | 262.4 ± 40.7 | 364.6 ± 31.9 | 195.3 ± 7.8 a |
| Highly calcareous clay | Control 0 ppm CO2 | 20.36 ± 0.8 | 21.89 ± 1.9 | 12.6 ± 1.9 b | 209.9 ± 25.1 | 317.5 ± 68.1 a | 109.7 ± 10.5 b |
| Highly calcareous clay | 400 ppm CO2 | 19.98 ± 1.9 | 21.11 ± 1.9 | 26.4 ± 3.4 a | 201.7 ± 14.4 | 214.5 ± 50.1 b | 205.9 ± 26.2 a |
| Type of Soil | Irrigation Treatments | Shoot Starch (% d.w.) 1st Year | Root Starch (% d.w.) 1st Year | Root Citric Acid (mg/g d.w.) 2nd Year | Root Tartaric Acid (mg/g d.w.) 2nd Year |
|---|---|---|---|---|---|
| Schist Neutral Soil | Control 0 ppm CO2 | 7.8 ± 0.9 | 12.7 ± 0.6 b | 3.0 ± 0.5 b | 380.2 ± 74.1 b |
| Schist Neutral Soil | 400 ppm CO2 | 8.7 ± 0.7 | 29.8 ± 5.9 a | 8.6 ± 0.7 a | 505.8 ± 56.3 a |
| Medium calc-clay | Control 0 ppm CO2 | 5.7 ± 0.8 b | 14.5 ± 0.9 b | 4.2 ± 1.4 b | 579.9 ± 52.6 a |
| Medium calc-clay | 400 ppm CO2 | 10.9 ± 0.7 a | 23.1 ± 4.3 a | 10.3 ± 0.6 a | 254.3 ± 19.6 b |
| Highly calcareous clay | Control 0 ppm CO2 | 5.2 ± 0.5 b | 12.7 ± 1.3 b | 5.7 ± 0.8 | 1506.1 ± 485.9 a |
| Highly calcareous clay | 400 ppm CO2 | 9.9 ± 0.6 b | 32.9 ± 2.1 a | 5.9 ± 0.7 | 326.9 ± 19.27 b |
| FIRST YEAR | |||||
|---|---|---|---|---|---|
| Type of Soil | Irrigation Treatments | Dried Root Weight (g) | Dried Shoot Weight (g) | Shoot Length (cm) | |
| Schist Neutral Soil | Control 0 ppm CO2 | 82.61 ± 3.8 | 27.91 ± 3.2 b | 97.12 ± 7.3 | |
| Schist Neutral Soil | 400 ppm CO2 | 82.39 ± 9.4 | 39.89 ± 2.1 a | 101.64 ± 8.6 | |
| Medium calc-clay | Control 0 ppm CO2 | 58.25 ± 15.6 | 17.29 ± 3.8 | 62.21 ± 2.9 | |
| Medium calc-clay | 400 ppm CO2 | 54.33 ± 5.2 | 16.96 ± 2.1 | 72.29 ± 1.13 | |
| Highly calc-clay | Control 0 ppm CO2 | 21.53 ± 2.4 a | 12.70 ± 0.7 a | 19.71 ± 3.5 | |
| Highly calc-clay | 400 ppm CO2 | 7.67 ± 2.9 b | 1.84 ± 0.3 b | 19.51 ± 5.8 | |
| SECOND YEAR | |||||
| Type of Soil | Irrigation Treatments | Dried Root Weight (g) | Dried Shoot Weight (g) | Shoot Length (cm) | Leaf Surface (cm2) |
| Schist Neutral Soil | Control 0 ppm CO2 | 95.49 ± 9.5 | 62.54 ± 7.08 | 99.2 ± 8.4 | 3274.3 ± 467.9 |
| Schist Neutral Soil | 400 ppm CO2 | 92.03 ± 7.15 | 67.40 ± 4.20 | 96.4 ± 4.5 | 3490.3 ± 426.3 |
| Medium calc-clay | Control 0 ppm CO2 | 102.75 ± 6.45 b | 51.11 ± 5.82 | 82.7 ± 5.4 b | 3279.5 ± 387.0 b |
| Medium calc-clay | 400 ppm CO2 | 129.29 ± 18.0 a | 52.99 ± 8.67 | 94.2 ± 4.8 a | 4069.5 ± 621.0 a |
| Highly calc-clay | Control 0 ppm CO2 | 83.94 ± 5.32 b | 14.52 ± 2.54 a | 68.4 ± 9.4 a | 2024.3 ± 95.9 |
| Highly calc-clay | 400 ppm CO2 | 102.08 ± 13.9 a | 10.90 ± 2.44 b | 45.3 ± 4.2 b | 1512.3 ± 335.2 |
| Type of Soil | Irrigation Treatments | Berry Kg/Vine | pH | Brix | TA (g/L) | Tartaric Acid (g/L) | Malic Acid (g/L) |
|---|---|---|---|---|---|---|---|
| Schist Neutral Soil | Control 0 ppm CO2 | 17.2 ± 0.7 b | 3.91 ± 0.007 | 9.8 ± 0.2 a | 4.12 ± 0.07 | 3.85 ± 0.13 b | 2.08 ± 0.08 |
| Schist Neutral Soil | 400 ppm CO2 | 22.7 ± 0.4 a | 3.95 ± 0.009 | 9.5 ± 0.2 b | 3.82 ± 0.07 | 3.94 ± 0.11 a | 1.78 ± 0.02 |
| Medium calc-clay | Control 0 ppm CO2 | 17.5 ± 0.9 b | 3.78 ± 0.006 | 10.5 ± 0.3 b | 4.80 ± 0.01 a | 4.45 ± 0.27 | 2.21 ± 0.15 |
| Medium calc-clay | 400 ppm CO2 | 24.8 ± 1.1 a | 3.75 ± 0.01 | 10.8 ± 0.1 a | 4.35 ± 0.01 b | 3.85 ± 0.02 | 2.24 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lampreave, M.; Mateos, A.; Valls, J.; Nadal, M.; Sánchez-Ortiz, A. Carbonated Irrigation Assessment of Grapevine Growth, Nutrient Absorption, and Sugar Accumulation in a Tempranillo (Vitis vinifera L.) Vineyard. Agriculture 2022, 12, 792. https://doi.org/10.3390/agriculture12060792
Lampreave M, Mateos A, Valls J, Nadal M, Sánchez-Ortiz A. Carbonated Irrigation Assessment of Grapevine Growth, Nutrient Absorption, and Sugar Accumulation in a Tempranillo (Vitis vinifera L.) Vineyard. Agriculture. 2022; 12(6):792. https://doi.org/10.3390/agriculture12060792
Chicago/Turabian StyleLampreave, Miriam, Assumpta Mateos, Josep Valls, Montserrat Nadal, and Antoni Sánchez-Ortiz. 2022. "Carbonated Irrigation Assessment of Grapevine Growth, Nutrient Absorption, and Sugar Accumulation in a Tempranillo (Vitis vinifera L.) Vineyard" Agriculture 12, no. 6: 792. https://doi.org/10.3390/agriculture12060792