
Heterosis for Nitrogen Use Efficiency of Maize Hybrids Enhanced over Decades in China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Location
2.2. Experimental Design
2.3. Sampling and Measurements
2.3.1. Plant Sampling
2.3.2. Nitrogen and Heterosis Indices
2.4. Statistical Analysis
3. Results
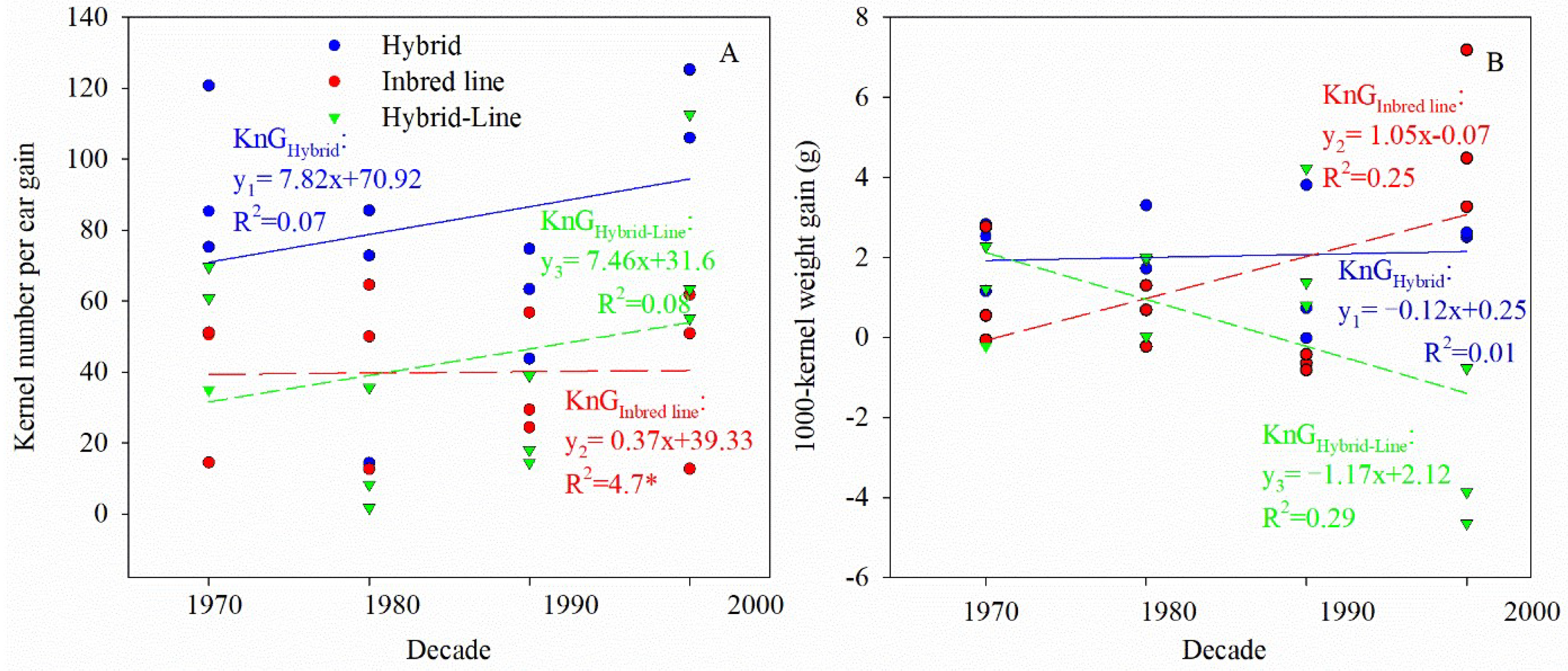
3.1. Heterotic Performance of Yield and Yield Components of Hybrids
3.2. The Heterosis of NUE and Its Related Parameters Indexes
3.3. Physiological Basis Underlying the Heterosis for NUE
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qin, X.; Fan, F.; Li, Y.; Xu, S.; Siddique, K.; Liao, Y.; Lübberstedt, T. Maize yield improvements in china: Past trends and future directions. Plant Breed. 2016, 135, 166–176. [Google Scholar] [CrossRef]
- He, H.; Hu, Q.; Li, R.; Pan, X.; He, Q. Regional gap in maize production, climate and resource utilization in China. Field Crops Res. 2020, 254, 107830. [Google Scholar] [CrossRef]
- Li, R.; Zhang, G.; Liu, G.; Wang, K.; Xie, R.; Hou, P.; Ming, B.; Wang, Z.; Li, S. Improving the yield potential in maize by constructing the ideal plant type and optimizing the maize canopy structure. Food Energy Secur. 2021, 10, e312. [Google Scholar] [CrossRef]
- Hou, P.; Liu, Y.; Liu, W.; Liu, G.; Xie, R.; Wang, K.; Ming, B.; Wang, Y.; Zhao, R.; Zhang, W. How to increase maize production without extra nitrogen input. Resour. Conserv. Recycl. 2020, 160, 104913. [Google Scholar] [CrossRef]
- Ning, P.; Fritschi, F.B.; Li, C. Temporal dynamics of post-silking nitrogen fluxes and their effects on grain yield in maize under low to high nitrogen inputs. Field Crops Res. 2017, 204, 249–259. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, B.; Yu, X.; Gao, J.; Sun, J.; Su, Z. Physiological basis of heterosis for nitrogen use efficiency of maize. Sci. Rep. 2019, 9, 18708. [Google Scholar] [CrossRef]
- Coque, M.; Gallais, A. Genetic variation among European maize varieties for nitrogen use efficiency under low and high nitrogen fertilization. Maydica 2007, 52, 383–397. [Google Scholar]
- Ciampitti, I.A.; Vyn, T.J. A comprehensive study of plant density consequences on nitrogen uptake dynamics of maize plants from vegetative to reproductive stages. Field Crops Res. 2011, 121, 2–18. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Giuzio, L.; Caro, A.D.; Flagella, Z. Relationships between nitrogen utilization and grain technological quality in durum wheat: I. nitrogen translocation and nitrogen use efficiency for protein. Agron. J. 2011, 103, 1487–1494. [Google Scholar] [CrossRef]
- Shu, G.; Cao, G.; Li, N.; Wang, A.; Wei, F.; Li, T.; Li, Y.; Xu, Y.; Wang, Y. Genetic variation and population structure in China summer maize germplasm. Sci. Rep. 2021, 11, 8012. [Google Scholar] [CrossRef]
- Kumar, B.; Rakshit, S.; Kumar, S.; Singh, B.K.; Lahkar, C.; Jha, A.K.; Kumar, K.; Kumar, P.; Choudhary, M.; Singh, S.B.; et al. Genetic Diversity, Population Structure and Linkage Disequilibrium Analyses in Tropical Maize Using Genotyping by Sequencing. Plants 2022, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Duvick, D.N.; Smith, J.S.C.; Cooper, M. Long-term selection in a commercial hybrid maize breeding program. Plant Breed. Rev. 2004, 24, 109–151. [Google Scholar]
- Duvick, D.N. Genetic progress in yield of United States maize (Zea mays L.). Maydica 2005, 50, 193–202. [Google Scholar]
- Curin, F.; Severini, A.D.; González, F.G.; Otegui, M.E. Water and radiation use efficiencies in maize: Breeding effects on single-cross Argentine hybrids released between 1980 and 2012. Field Crops Res. 2020, 246, 107683. [Google Scholar] [CrossRef]
- Li, Y.X.; Li, Y.; Ma, X.L.; Liu, C.; Liu, Z.Z. Contributions of parental inbreds and heterosis to morphology and yield of single-cross maize hybrids in China. Crop Sci. 2014, 54, 76–88. [Google Scholar] [CrossRef]
- Ma, D.L.; Xie, R.Z.; Liu, X.; Niu, X.K.; Hou, P.; Wang, K.R.; Li, S.K. Lodging-related stalk characteristics of maize varieties in China since the 1950s. Crop Sci. 2014, 54, 2805–2814. [Google Scholar] [CrossRef]
- Ma, D.L.; Xie, R.Z.; Niu, X.K.; Li, S.K.; Long, H.L.; Liu, Y.E. Changes in the morphological traits of maize genotypes in China between the 1950s and 2000s. Eur. J. Agron. 2014, 58, 1–10. [Google Scholar] [CrossRef]
- Kayad, A.; Sozzi, M.; Gatto, S.; Whelan, B.; Sartori, L.; Marinello, F. Ten years of corn yield dynamics at field scale under digital agriculture solutions: A case study from North Italy. Comput. Electron. Agric. 2021, 185, 106126. [Google Scholar] [CrossRef]
- Li, X.; Guo, L.; Zhou, B.; Tang, X.; Chen, C.; Zhang, L.; Zhang, S.-Y.; Li, C.-F.; Xiao, K.; Dong, W.-X. Characterization of low-N responses in maize (Zea mays L.) cultivars with contrasting nitrogen use efficiency in the North China Plain. J. Integr. Agric. 2019, 18, 2141–2152. [Google Scholar] [CrossRef]
- Ma, B.L.; Dwyer, L.M.; Tollenaar, M.; Smith, D.L. Stem infusion of nitrogen-15 to quantify nitrogen remobilization in maize. Commun. Soil Sci. Plant Anal. 1998, 29, 305–317. [Google Scholar] [CrossRef]
- Carlone, M.R.; Russell, W.A. Response to plant densities and nitrogen levels for four maize cultivars from different eras of breeding. Crop Sci. 1987, 27, 465–470. [Google Scholar] [CrossRef]
- Blum, A. Heterosis, stress, and the environment: A possible road map towards the general improvement of crop yield. J. Exp. Bot. 2013, 64, 4829–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duvick, D.N. Heterosis: Feeding people and protecting natural resources. In The Genetics and Exploitation of Heterosis in Crops; Coors, J.D., Pandey, S., Eds.; ASA, CSSA and SSSA: Madison, WI, USA, 1999; pp. 19–29. [Google Scholar]
- Tollenaar, M.; Deen, W.; Echarte, L.; Liu, W. Effect of crowding stress on dry matter accumulation and harvest index in maize. Agron. J. 2006, 98, 930–937. [Google Scholar] [CrossRef]
- Tollenaar, M.; Ahmadzadeh, A.; Lee, E.A. Physiological basis of heterosis for grain yield in maize. Crop Sci. 2004, 44, 2086–2094. [Google Scholar] [CrossRef]
- Hisse, I.R.; D’Andrea, K.E.; Otegui, M.E. Source-sink relations and kernel weight in maize inbred lines and hybrids: Responses to contrasting nitrogen supply levels. Field Crops Res. 2019, 230, 151–159. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, L.; Bian, S.; Liu, Z.; Wang, Y.; Lv, Y.; Cao, Y.; Yao, F.; Li, C.; Wei, W. Evolution of roots to improve water and nitrogen use efficiency in maize elite inbred lines released during different decades in China. Agric. Water Manag. 2019, 216, 44–59. [Google Scholar] [CrossRef]
- Bertin, P.; Gallais, A. Genetic variation for nitrogen use efficiency in a set of recombinant maize inbred lines. I. Agrophysiological results. Maydica 2000, 45, 53–66. [Google Scholar]
- Ning, P.; Liao, C.; Li, S.; Yu, P.; Zhang, Y.; Li, X.; Li, C. Maize cob plus husks mimics the grain sink to stimulate nutrient uptake by roots. Field Crops Res. 2012, 130, 38–45. [Google Scholar] [CrossRef]
- D’Andrea, K.E.; Otegui, M.E.; Cirilo, A.G.; Eyherabide, G.H. Genotypic variability in morphological and physiological traits among maize inbred lines: Nitrogen responses. Crop Sci. 2006, 46, 1266–1276. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, C.; Chen, X.; Li, Q.; Zhang, J.; Chen, F. Characterization of the plant traits contributed to high grain yield and high grain nitrogen concentration in maize. Field Crops Res. 2014, 159, 1–9. [Google Scholar] [CrossRef]
- Gaju, O.; Allard, V.; Martre, P.; Le Gouis, J.; Moreau, D.; Bogard, M.; Hubbart, S.; Foulkes, M.J. Nitrogen partitioning and remobilization in relation to leaf senescence, grain yield and grain nitrogen concentration in wheat cultivars. Field Crops Res. 2014, 155, 213–223. [Google Scholar] [CrossRef]
- Munaro, E.M.; D’Andrea, K.E.; Otegui, M.E.; Cirilo, A.G.; Eyhérabide, G.H. Heterotic response for grain yield and ecophysiological related traits to nitrogen availability in maize. Crop Sci. 2011, 51, 1172–1187. [Google Scholar] [CrossRef]
- Tollenaar, M.; Lee, E.A. Strategies for enhancing grain yield in maize. Plant Breed. Rev. 2011, 34, 37–82. [Google Scholar]
- Edlich-Muth, C.; Muraya, M.M.; Altmann, T.; Selbig, J. Phenomic prediction of maize hybrids. Biosystems 2016, 146, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.L. Variation Trend of Yield and Nitrogen Efficiency of Maize Varieties and Their Parents at Different Ages in China. Ph.D. Dissertation, Chinese Academy of Agricultural Sciences, Beijing, China, 2014. [Google Scholar]
- Chen, K.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Plant biomass and nitrogen partitioning changes between silking and maturity in newer versus older maize hybrids. Field Crops Res. 2015, 183, 315–328. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiency: A review. Field Crops Res. 2012, 133, 48–67. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Grain nitrogen source changes over time in maize: A review. Crop Sci. 2013, 53, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Haegele, J.W.; Cook, K.A.; Nichols, D.M.; Below, F.E. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades. Crop Sci. 2013, 53, 1256–1268. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.M.; Vyn, T.J. Maize plant resilience to N stress and post-silking N capacity changes over time: A review. Front. Plant Sci. 2016, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Yue, S.; Qiu, M.; Chen, X.; Cui, Z.; Chen, F. Using maize hybrids and in-season nitrogen management to improve grain yield and grain nitrogen concentrations. Field Crops Res. 2014, 166, 38–45. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrids | Decade | Parental Combination | Breeding/Provide Institution | Year of Hybrid Release |
|---|---|---|---|---|
| Jidan 101 | 1970s | Ji 63 × M 14 | Jilin Academy of Agricultural Sciences | 1974 |
| Zhongdan 2 | Mo 17 × Zi 330 | Chinese Academy of Agricultural Sciences | 1973 | |
| Danyu 13 | 1980s | Mo 17 × E 28 | Dandong Academy of Agricultural Sciences | 1979 |
| Nongda 60 | Shen 5003 × Zong 31 | China Agricultural University | 1985 | |
| Yedan 13 | 1990s | Ye 478 × Dan 340 | Laizhou Academy of Agricultural Sciences | 1989 |
| Nongda 108 | X 178 × HangC | China Agricultural University | 1991 | |
| Xianyu 335 | 2000s | PH6WC × PH4CV | Seed-Pioneer Tieling Co., Ltd. | 2000 |
| Zhengdan 958 | Zheng 58 × Chang 7–2 | Henan Academy of Agricultural Sciences | 1996 |
| Index | Grain Yield (t·ha−1) | Yield Gain (t·ha−1) | Grain Number per Ear | Grain Number Gain | 1000-Kernel Weight (g) | 1000-Kernel Weight Gain (g) | |||
|---|---|---|---|---|---|---|---|---|---|
| 150 N | 0 N | 150 N | 0 N | 150 N | 0 N | ||||
| Era of Hybrids | |||||||||
| 1970s | 11.5 d | 10.0 b | 1.5 d | 559.7 d | 499.1 b | 60.6 b | 341.2 b | 331.7 b | 9.5 b |
| 1980s | 11.9 c | 10.3 b | 1.6 c | 587.3 c | 514.4 a | 72.9 c | 333.3 c | 328.7 b | 4.6 c |
| 1990s | 12.6 b | 10.1 b | 2.5 b | 594.5 b | 482.6 c | 111.9 b | 343.1 b | 319.5 c | 23.6 a |
| 2000s | 14.3 a | 10.8 a | 3.5 a | 603.2 a | 426.5 d | 176.7 a | 369.9 a | 347.0 a | 22.9 a |
| Source of variance | |||||||||
| Era (E) | ** | ns | ** | ** | ns | ** | ns | ns | ns |
| N rate (N) | ** ns | - | ** ** | - | ** | - | |||
| E × N | ns ns | - | ns ns | - | ns ns | - | |||
| Heterotic Index | Source of Variation | ||
|---|---|---|---|
| Era of Hybrids (E) | N Rate (N) | (E × N) | |
| NUE | 5.93 * | - | - |
| NIE | 13.06 * | - | - |
| NRE | 9.12 * | - | - |
| Plant N | 61.45 ** | 22.29 ** | 1.75 |
| Grain N | 31.71 ** | 23.79 ** | 7.07 * |
| Rem N | 25.87 ** | 32.96 * | 3.07 |
| Rem Leaf N | 8.23 * | 2.11 | 6.20 * |
| Rem Stalk N | 9.56 * | 0.56 | 3.77 |
| VegN | 23.71 ** | 17.32 * | 1.36 |
| RepN | 59.87 ** | 32.96 ** | 44.3 * |
| NHI | 4.12 | 0.99 | 0.84 |
| BMRsilk | 11.46 * | 30.25 ** | 1.61 |
| BMRpm | 10.17 * | 31.09 ** | 0.58 |
| HI | 0.36 | 0.01 | 3.57 |
| Index | 1970 | 1980 | 1990 | 2000 | ||||
|---|---|---|---|---|---|---|---|---|
| 150 N | 0 N | 150 N | 0 N | 150 N | 0 N | 150 N | 0 N | |
| NUE | 8.7 ab | - | 7.3 b | - | 8.4 ab | - | 14.0 a | - |
| NIE | 10.2 a | - | 5.4 a | - | 11.5 a | - | 38.1 a | - |
| NRE | 0.2 a | - | 0.1 a | - | 0.1 a | - | −0.1 a | - |
| Plant N | 68.0 ab | 45.1 b | 43.3 bc | 25.5 b | 30.5 c | 20.5 b | 96.4 a | 114.3 a |
| Grain N | 45.5 b | 30.8 | 30.2 b | 14.0 | 34.2 b | 8.7 | 82.1 a | 84.6 a |
| Rem N | 33.7 | 16.2 | 75.1 | 12.9 | 51.9 | 11.1 | 77.4 | 16.8 |
| Rem Leaf N | 21.9 | 10.5 | 48.8 | 8.4 | 33.7 | 7.2 | 47.3 | 16.7 |
| Rem Stalk N | 11.8 | 5.7 | 26.3 | 4.5 | 18.2 | 3.9 | 25.5 | 9.0 |
| VegN | 56.2 | 30.5 | 88.1 | 24.5 | 48.3 | 22.9 | 91.7 | 46.5 |
| RepN | 11.8 | 14.6 ab | −44.8 | 1.1 b | −17.8 | −2.4b | 4.7 | 67.8 a |
| BMsilk | 3.1 | 2.5 ab | 3.0 | 1.4 b | 3.5 | 1.7 ab | 3.4 | 2.6 a |
| BMpm | 7.9 a | 5.7 | 6.9 ab | 4.5 | 4.4 b | 4.0 | 7.8 ab | 8.8 |
| Index | 1970 | 1980 | 1990 | 2000 | ||||
|---|---|---|---|---|---|---|---|---|
| 150 N (%) | 0 N (%) | 150 N (%) | 0 N (%) | 150 N (%) | 0 N (%) | 150 N (%) | 0 N (%) | |
| NUE | 177.4 | - | 223.8 | - | 186.2 | - | 312.9 | - |
| NIE | 76.6 b | - | 25.1 b | - | 51.3 b | - | 232.2 a | - |
| NRE | 64.6 a | - | 32.9 ab | - | 27.3 ab | - | −13.9 b | - |
| Plant N | 27.4 ab | 20.2 b | 23.7 b | 21.6 b | 15.5 c | 17.7 b | 32.9 a | 52.3 a |
| Grain N | 24.4 b | 23.7 b | 32.9 b | 27.3 b | 24.7 b | 15.0 b | 50.2 a | 64.7 a |
| Rem N | 79.9 | 37.2 | 122.8 | 56.9 | 94.0 | 28.1 | 193.6 | 56.4 |
| Rem Leaf N | 79.9 | 37.2 | 122.8 | 56.9 | 94.0 | 28.1 | 172.6 | 78.2 |
| Rem Stalk N | 79.9 | 37.2 | 122.8 | 56.9 | 94.0 | 28.1 | 172.6 | 78.2 |
| VegN | 33.0 | 23.8 ab | 45.6 | 29.8 a | 27.4 | 15.2 b | 53.3 | 32.4 a |
| RepN | 20.2 | 13.0 bc | −18.2 | 1.7c | −4.8 | 87.5 ab | 5.9 | 101.9 a |
| BMsilk | 38.8 | 33.5 ab | 35.7 | 16.4 c | 40.6 | 20.1 bc | 45.1 | 39.1 a |
| BMpm | 52.1 a | 38.0 b | 40.6 ab | 36.4 b | 24.0 b | 27.1 b | 45.2 a | 64.6 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Gao, J.; Li, Y.; Yu, S.; Wang, Z. Heterosis for Nitrogen Use Efficiency of Maize Hybrids Enhanced over Decades in China. Agriculture 2022, 12, 764. https://doi.org/10.3390/agriculture12060764
Li R, Gao J, Li Y, Yu S, Wang Z. Heterosis for Nitrogen Use Efficiency of Maize Hybrids Enhanced over Decades in China. Agriculture. 2022; 12(6):764. https://doi.org/10.3390/agriculture12060764
Chicago/Turabian StyleLi, Rongfa, Julin Gao, Yuanyuan Li, Shaobo Yu, and Zhigang Wang. 2022. "Heterosis for Nitrogen Use Efficiency of Maize Hybrids Enhanced over Decades in China" Agriculture 12, no. 6: 764. https://doi.org/10.3390/agriculture12060764