
Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Locations and Experimental Design
2.2. Measurements
2.3. Statistical Analysis
2.3.1. Linear Mixed Model
2.3.2. AMMI Model Analysis
2.3.3. BLUP Technique
2.3.4. Combining of AMMI Analysis and BLUP Techniques
3. Results
3.1. The Prediction Accuracy of BLUP and AMMI Model
3.2. Variance Components of Yield Components
3.3. Three-Way Analysis of Variance
3.4. AMMI Analysis of Variance
3.5. WAASB Scores of Evaluated Genotypes
3.6. Integrate AMMI Model and BLUP Technology to Understand the GEI
3.7. Correlation and Cluster Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ci, X.; Li, M.; Liang, X.; Xie, Z.; Zhang, D.; Li, X.; Lu, Z.; Ru, G.; Bai, L.; Xie, C.; et al. Genetic contribution to advanced yield for maize hybrids released from 1970 to 2000 in China. Crop Sci. 2011, 51, 13–20. [Google Scholar] [CrossRef]
- Xu, W.; Liu, C.; Wang, K.; Xie, R.; Ming, B.; Wang, Y.; Zhang, G.; Liu, G.; Zhao, R.; Fan, P.; et al. Adjusting maize plant density to different climatic conditions across a large longitudinal distance in China. Field Crop Res. 2017, 212, 126–134. [Google Scholar] [CrossRef]
- Liu, G.; Yang, H.; Xie, R.; Yang, Y.; Liu, W.; Guo, X.; Xue, J.; Ming, B.; Wang, K.; Hou, P.; et al. Genetic gains in maize yield and related traits for high-yielding cultivars released during 1980s to 2010s in China. Field Crop Res. 2021, 270, 108223. [Google Scholar] [CrossRef]
- FAO—Food and Agriculture Organization of the United Nations. FAO Statistical Year Book; FAO: Rome, Italy, 2020. [Google Scholar]
- Padi, F.K. Genotype × environment interaction and yield stability in a cowpea-based cropping system. Euphytica 2007, 158, 11–25. [Google Scholar] [CrossRef]
- Krause, M.D.; Dias, K.O.D.; dos Santos, J.P.R.; de Oliveira, A.A.; Guimaraes, L.J.M.; Pastina, M.M.; Margarido, G.R.A.; Garcia, A.A.F. Boosting predictive ability of tropical maize hybrids via genotype-by-environment interaction under multivariate GBLUP models. Crop Sci. 2020, 60, 3049–3056. [Google Scholar] [CrossRef]
- Cooper, M.; Technow, F.; Messina, C.; Gho, C.; Totir, L.R. Use of crop growth models with whole-genome prediction: Application to a maize multienvironment trial. Crop Sci. 2016, 56, 2141–2156. [Google Scholar] [CrossRef] [Green Version]
- Bocianowski, J.; Warzecha, T.; Nowosad, K.; Bathelt, R. Genotype by environment interaction using AMMI model and estimation of additive and epistasis gene effects for 1000-kernel weight in spring barley (Hordeum vulgare L.). J. Appl. Genet. 2019, 60, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.S. Genotype-environment interaction and stability analyses: An update. In Quantitative Genetics, Genomics and Plant Breeding, 2nd ed.; CAB International: Wallingford, UK, 2020. [Google Scholar]
- Yan, W.K.; Hunt, L.A.; Sheng, Q.L.; Szlavnics, Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Dehghani, H.; Sabaghpour, S.H.; Ebadi, A. Study of genotype × environment interaction for chickpea yield in Iran. Agron. J. 2010, 102, 1–8. [Google Scholar] [CrossRef]
- Smith, A.; Ganesalingam, A.; Lisle, C.; Kadkol, G.; Cullis, B. Use of contemporary groups in the construction of multi-environment trial datasets for selection in plant breeding programs. Front. Plant Sci. 2021, 11, 623586. [Google Scholar] [CrossRef]
- Vaezi, B.; Pour-Aboughadareh, A.; Mohammadi, R.; Armion, M.; Mehraban, A.; Hossein-Pour, T.; Dorri, M. GGE biplot and AMMI analysis of barley yield performance in Iran. Cereal Res. Commun. 2017, 45, 500–511. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.S.; Gorman, D.P. Genotype × environment interaction in maize. Agron. J. 1989, 81, 662–664. [Google Scholar] [CrossRef]
- Babić, V.; Babić, M.; Ivanović, M.; Kraljević-Balalić, M.; Dimitrijević, M. Understanding and utilization of genotype-by-environment interaction in maize breeding. Genetika 2010, 42, 79–90. [Google Scholar] [CrossRef]
- Yousaf, M.I.; Akhtar, N.; Mumtaz, A.; Shehzad, A.; Mehboob, A. Yield stability studies in indigenous and exotic maize hybrids under genotype by environment interaction. Pak. J. Bot. 2021, 53, 1–8. [Google Scholar] [CrossRef]
- Paderewski, J.; Gauch, H.G.; Madry, W.; Gacek, E. AMMI analysis of four-way genotype × location management × year data from a wheat trial in Poland. Crop Sci. 2016, 56, 2157–2164. [Google Scholar] [CrossRef]
- Gauch, H.G. Model selection and validation for yield trials with interaction. Biometrics 1988, 44, 705–715. [Google Scholar] [CrossRef]
- Alemu, G.; Geleta, N.; Dabi, A.; Delessa, A.; Duga, R. Stability models for selecting adaptable and stable bread wheat (Tritium aestivum L.) varieties for grain yield in Ethiopia. J. Agric. Sci. Eng. 2021, 7, 14–22. [Google Scholar]
- Gauch, H.; Moran, D.R. AMMISOFT for AMMI analysis with best practices. bioRxiv 2019, 538454. [Google Scholar] [CrossRef]
- Gauch, H.G. A simple protocol for AMMI analysis of yield trials. Crop Sci. 2013, 53, 1860–1869. [Google Scholar] [CrossRef]
- Ali, M.; Elsadek, A.; Salem, E.M. Stability Parameters and AMMI Analysis of Quinoa (Chenopodium quinoa Willd.). Egypt. J. Agron. 2018, 40, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Agahi, K.; Ahmadi, J.; Oghan, H.A.; Fotokian, M.H.; Orang, S.F. Analysis of genotype × environment interaction for seed yield in spring oilseed rape using the AMMI model. Crop Breed. Appl. Biot. 2020, 20, e26502012. [Google Scholar] [CrossRef]
- Gurmu, F.; Shimelis, H.; Laing, M.; Mashilo, J. Genotype-by-environment interaction analysis of nutritional composition in newly-developed sweetpotato clones. J. Food Compos. Anal. 2020, 88, 103426. [Google Scholar] [CrossRef]
- Rodrigues, P.C.; Monteiro, A.; Lourenc, V.M. A robust AMMI model for the analysis of genotype-by-environment data. Bioinformatics 2016, 32, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivoto, T.; Lúcio, A.D.; da Silva, J.A.; Marchioro, V.S.; de Souza, V.Q.; Jost, E. Mean performance and stability in multi-environment trials I: Combining features of AMMI and BLUP techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- Karimizadeh, R.; Pezeshkpour, P.; Barzali, M.; Mehraban, A.; Sharifi, P. Evaluation the mean performance and stability of lentil genotypes by combining features of AMMI and BLUP techniques. J. Crop Breed. 2020, 12, 160–170. [Google Scholar]
- Singamsetti, A.; Shahi, J.P.; Zaidi, P.H.; Seetharam, K.; Vinayan, M.T.; Kumar, M.; Madankar, K. Genotype × environment interaction and selection of maize (Zea mays L.) hybrids across moisture regimes. Field Crop Res. 2021, 270, 108224. [Google Scholar] [CrossRef]
- Huang, X.; Jang, S.; Kim, B.; Piao, Z.; Redona, E.; Koh, H.J. Evaluating Genotype × Environment Interactions of Yield Traits and Adaptability in Rice Cultivars Grown under Temperate, Subtropical and Tropical Environments. Agriculture 2021, 11, 558. [Google Scholar] [CrossRef]
- Hilmarsson, H.S.; Rio, S.; Sánchez, J.I.Y. Genotype by Environment Interaction Analysis of Agronomic Spring Barley Traits in Iceland Using AMMI, Factorial Regression Model and Linear Mixed Model. Agronomy 2021, 11, 499. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Ahakpaz, F.; Neyestani, E.; Hesami, A.; Mohammadi, B.; Mahmoudi, K.N.; Abedi-Asl, G.; Noshabadi, M.R.J.; Noshabadi, M.R.J.; Ahakpaz, F.; Alipour, H. Genotype-by-environment interaction analysis for grain yield of barley genotypes under dryland conditions and the role of monthly rainfall. Agr. Water Manag. 2021, 245, 106665. [Google Scholar] [CrossRef]
- Hadasch, S.; Forkman, J.; Piepho, H.P. Cross-Validation in AMMI and GGE Models: A Comparison of Methods. Crop Sci. 2017, 57, 264–274. [Google Scholar] [CrossRef]
- Piepho, H.P. Best linear unbiased prediction (BLUP) for regional yield trials: A comparison to additive main effects and multiplicative interaction (AMMI) analysis. Theor. Appl. Genet. 1994, 89, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.; Timsina, D. Analysis of yield and yield attributing traits of maize genotypes in Chitwan, Nepal. World J. Agric. Res. 2015, 3, 153–162. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M.; Obeng-Antwi, K.; Osuman, A.S.; Ado, S.G.; Coulibay, N.; Yallou, C.G.; Abdulai, M.; Boakyewaa, G.A.; Didjeira, A. Performance of extra-early maize cultivars based on GGE biplot and AMMI analysis. J. Agric. Sci. 2012, 150, 473–483. [Google Scholar] [CrossRef]
- Bernardo Júnior, L.A.Y.; de Silva, C.P.; de Oliveira, L.A.; Nuvunga, J.J.; Pires, L.P.M.; Von Pinho, R.G.; Balestre, M. AMMI Bayesian models to study stability and adaptability in maize. Agron. J. 2018, 110, 1765–1776. [Google Scholar] [CrossRef]
- Alwala, S.; Kwolek, T.; McPherson, M.; Pellow, J.; Meyer, D. A comprehensive comparison between Eberhart and Russell joint regression and GGE biplot analyses to identify stable and high yielding maize hybrids. Field Crop Res. 2010, 119, 225–230. [Google Scholar] [CrossRef]
- Yue, H.; Jiang, X.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Bu, J. A study on genotype-by-environment interactions for the multiple traits of maize hybrids in China. Agron. J. 2021, 113, 4889–4899. [Google Scholar] [CrossRef]
- Piepho, H.P.; Möhring, J.; Melchinger, A.E.; Büchse, A. BLUP for phenotypic selection in plant breeding and variety testing. Euphytica 2008, 161, 209–228. [Google Scholar] [CrossRef]
- Gauch, H.G.; Piepho, H.P.; Annicchiarico, P. Statistical analysis of yield trials by AMMI and GGE: Further considerations. Crop Sci. 2008, 48, 866–889. [Google Scholar] [CrossRef]
- Krishnamurthy, S.L.; Sharma, P.C.; Sharma, D.K.; Singh, Y.P.; Singh, R.K. Additive main effects and multiplicative interaction analyses of yield performance in rice genotypes for general and specific adaptation to salt stress in locations in India. Euphytica 2021, 217, 20. [Google Scholar] [CrossRef]
- Shahriari, Z.; Heidari, B.; Dadkhodaie, A. Dissection of genotype × environment interactions for mucilage and seed yield in Plantago species: Application of AMMI and GGE biplot analyses. PLoS ONE 2018, 13, e0196095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Singh, G.P. Stability index based on weighted average of absolute scores of AMMI and yield of wheat genotypes evaluated under restricted irrigated conditions for peninsular zone. Int. J. Agric. Environ. Biotechnol. 2020, 13, 371–381. [Google Scholar] [CrossRef]
- Abdelghany, A.M.; Zhang, S.; Azam, M.; Shaibu, A.S.; Feng, Y.; Qi, J.; Sun, J. Exploring the phenotypic stability of soybean seed compositions using multi-trait stability index approach. Agronomy 2021, 11, 2200. [Google Scholar] [CrossRef]
- Sellami, M.H.; Pulvento, C.; Lavini, A. Selection of suitable genotypes of lentil (Lens culinaris Medik.) under rainfed conditions in south Italy using multi-trait stability index (MTSI). Agronomy 2021, 11, 1807. [Google Scholar] [CrossRef]
- Erfani, A.; Abbasian, A.; Mohaddesi, A. Stability of some of rice genotypes based on WAASB and MTSI indices. Iran. J. Genet. Plant Breed. 2021, 9, 1–11. [Google Scholar]
- Wang, T.; Ma, X.; Li, Y.; Bai, D.; Liu, C.; Liu, Z.; Smith, S. Changes in yield and yield components of single-cross maize hybrids released in China between 1964 and 2001. Crop Sci. 2011, 51, 512–525. [Google Scholar] [CrossRef]
- Qin, X.; Feng, F.; Li, Y.; Xu, S.; Siddique, K.H.; Liao, Y. Maize yield improvements in China: Past trends and future directions. Plant Breed. 2016, 135, 166–176. [Google Scholar] [CrossRef]
- Li, Y.; Ma, X.; Wang, T.; Li, Y.; Liu, C.; Liu, Z.; Smith, S. Increasing maize productivity in China by planting hybrids with germplasm that responds favorably to higher planting densities. Crop Sci. 2011, 51, 2391–2400. [Google Scholar] [CrossRef] [Green Version]
- Ramreddy, V.; Jabeen, F. Narrow sense heritability, correlation and path analysis in maize (Zea mays L.). SABRAO J. Breed. Genet. 2016, 48, 120–126. [Google Scholar]
- Ubi, G.M.; Onabe, M.B.; Kalu, S.E. Path coefficient analysis, character association and variability studies in selected maize (Zea mays L.) genotypes grown in Southern Nigeria. Annu. Res. Rev. Biol. 2019, 33, 1–6. [Google Scholar]





{kind=link}
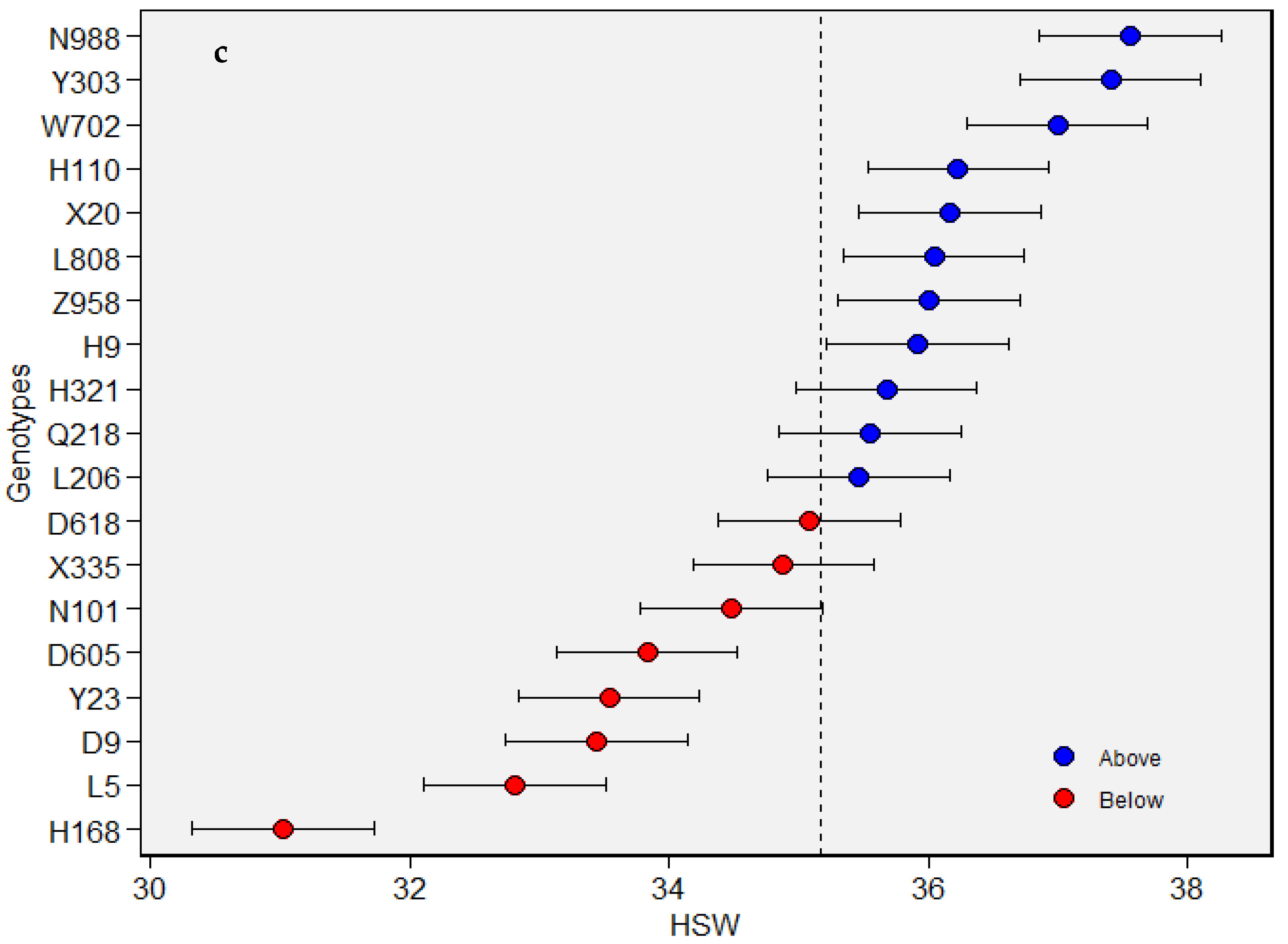{kind=link}
{kind=link}
{kind=link}
| Statistics | Likelihood Ratio Test | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Grain Yield (t/ha) | Ear Length (cm) | Hundred Seed Weight (g) | |||||||
| G | E | GE | G | E | GE | G | E | GE | |
| χ2 | 262 | 129 | 7808 | 300 | 136 | 7116 | 375 | 19.8 | 4025 |
| p value | 4.09 × 10−19 | 5.31 × 10−30 | 0 | 4.12 × 10−27 | 1.71 × 10−31 | 0 | 1.68 × 10−18 | 8.64 × 10−6 | 0 |
| REML | Estimated variance components | ||||||||
| 0.1764 (20.9%) | 0.4486 (23.0%) | 2.839 (26.1%) | |||||||
| 0.6484 (76.7%) | 1.442 (73.9%) | 6.977 (64.1%) | |||||||
| 0.0207 (2.4%) | 0.0605 (3.1%) | 1.068 (9.8%) | |||||||
| 0.846 | 1.951 | 10.880 | |||||||
| H2 | 0.209 | 0.230 | 0.261 | ||||||
| 0.767 | 0.739 | 0.641 | |||||||
| 0.952 | 0.958 | 0.966 | |||||||
| As | 0.976 | 0.979 | 0.983 | ||||||
| 0.969 | 0.960 | 0.867 | |||||||
| 3.98 | 3.692 | 4.792 | |||||||
| 1.36 | 1.355 | 2.940 | |||||||
| 2.92 | 2.724 | 1.630 | |||||||
| SD | 1.95 | 1.72 | 5.70 | ||||||
| SE | 0.03 | 0.03 | 0.09 | ||||||
| Traits | Source of Variance | DF | SS | MS | F Value | Pr (>F) | Percent of Total SS (%) |
|---|---|---|---|---|---|---|---|
| Genotype (G) | 18 | 740.50 | 41.14 | 51.684 *** | <0.001 | 4.63 | |
| Environment (E) | 36 | 9872 | 274.20 | 344.50 *** | <0.001 | 61.74 | |
| Grain yield | Year (Y) | 1 | 22 | 22.45 | 28.21 *** | <0.001 | 0.14 |
| G × E | 648 | 2303 | 3.56 | 3.56 *** | <0.001 | 14.40 | |
| G × Y | 18 | 130.9 | 7.27 | 9.14 *** | <0.001 | 0.82 | |
| E × Y | 36 | 538.3 | 14.95 | 18.79 *** | <0.001 | 3.37 | |
| G × E × Y | 648 | 148.1 | 0.228 | 0.29 ns | 1.00 | 0.93 | |
| Residuals | 2810 | 102.2 | 0.8 | ||||
| Total | 4217 | 15,994.13 | |||||
| Genotype (G) | 18 | 1873.63 | 103.98 | 357.76 *** | <0.001 | 14.96 | |
| Environment (E) | 36 | 4037.21 | 112.14 | 385.97 *** | <0.001 | 32.23 | |
| Ear length | Year (Y) | 1 | 19 | 19.07 | 66.16 *** | <0.001 | 0.15 |
| G × | 648 | 5264.26 | 8.13 | 27.97 *** | <0.001 | 42.02 | |
| G × Y | 18 | 378.87 | 21.06 | 72.47 *** | <0.001 | 3.02 | |
| E × Y | 36 | 21.51 | 0.60 | 2.05 *** | <0.001 | 0.17 | |
| G × E × Y | 648 | 115.06 | 0.18 | 1.03 ns | 0.295 | 0.92 | |
| Residuals | 2810 | 483.11 | 0.17 | ||||
| Total | 4217 | 12,527.07 | |||||
| Genotype (G) | 18 | 11,740.46 | 652.25 | 50.7 *** | <0.001 | 8.57 | |
| Environment (E) | 36 | 45,929.19 | 1275.81 | 99.18 *** | <0.001 | 33.52 | |
| Hundred seed weight | Year (Y) | 1 | 5728.02 | 5728.02 | 445.29 *** | <0.001 | 4.18 |
| G × E | 648 | 24,969.04 | 38.53 | 2.43 *** | <0.001 | 18.22 | |
| G × Y | 18 | 3861.11 | 214.51 | 16.68 *** | <0.001 | 2.82 | |
| E × Y | 36 | 45.18 | 1.25 | 0.1 ns | 1 | 0.03 | |
| G × E × Y | 648 | 84.13 | 0.13 | 0.01 ns | 1 | 0.06 | |
| Residuals | 2810 | 36,146.67 | 12.86 | ||||
| Total | 4217 | 137,015.75 |
| Source of Variance | df | SS | MS | Proportion of Variation | |||
|---|---|---|---|---|---|---|---|
| % of GE Signal and Noise | % of Variability Explained | % of GEI SS | % of GEIs Variation | ||||
| Treatment | 1405 | 13,755.78 | 9.79 *** | 86.01 | |||
| Genotype | 18 | 740.50 | 41.14 *** | ||||
| Environment (E) | 73 | 10,432.84 | 142.92 *** | ||||
| GE interaction (GEI) | 1314 | 2582.42 | 1.97 *** | ||||
| IPC1 | 90 | 741.6 | 8.24 *** | 28.72 | 29.02 | ||
| IPC2 | 88 | 523.49 | 5.95 *** | 20.27 | 20.49 | ||
| IPC3 | 86 | 251.37 | 2.92 *** | 9.73 | 9.84 | ||
| IPC4 | 84 | 174.54 | 2.08 *** | 6.76 | 6.83 | ||
| IPC5 | 82 | 161.41 | 1.97 *** | 6.25 | 6.32 | ||
| IPC6 | 80 | 152.44 | 1.91 *** | 5.90 | 5.97 | ||
| IPC7 | 78 | 106.02 | 1.36 *** | 4.11 | 4.15 | ||
| Residual | 726 | 471.55 | 0.65 *** | 18.26 | 18.45 | ||
| Error | 2812 | 2238.34 | 0.80 | 13.99 | |||
| Blocks/environment | 148 | 2183.21 | 14.75 *** | ||||
| Pure Error | 2664 | 55.13 | 0.02 | ||||
| GEIN | 27.19 | 1.05 | |||||
| GEIS | 2555.23 | 98.95 | |||||
| Total | 4217 | 15,994.13 | 3.79 | 100 | 100 | 100 | 100 |
| Genotype Code | Grain Yield (t/ha) | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | WAASB | rWAASB |
|---|---|---|---|---|---|---|---|---|---|---|
| D605 | 9.98 (17) | −0.459 | −1.58 | 0.135 | −0.181 | 0.33 | −0.451 | 0.147 | 0.594 | 12 |
| D618 | 9.77 (19) | 1.440 | −0.109 | −0.49 | −1.88 | −0.692 | −0.030 | −0.355 | 0.72 | 17 |
| D9 | 9.93 (18) | 2.6 | 0.658 | 0.196 | 0.932 | 0.326 | −0.741 | 0.011 | 1.08 | 19 |
| H110 | 10.8 (6) | 0.696 | −0.572 | 0.266 | 0.976 | −1.89 | 0.296 | −0.111 | 0.595 | 13 |
| H168 | 10.8 (7) | −0.993 | 0.384 | 0.124 | 0.351 | 0.542 | 0.044 | −0.462 | 0.538 | 9 |
| H321 | 10.8 (8) | −0.312 | −0.558 | −0.093 | 0.402 | 0.018 | 0.623 | 0.15 | 0.341 | 3 |
| H9 | 11 (3) | −0.923 | 0.48 | 0.472 | −0.162 | −0.433 | 0.318 | 0.54 | 0.575 | 11 |
| L206 | 11.2 (2) | −0.790 | 0.738 | −0.053 | −0.083 | −0.774 | −1.300 | 1.06 | 0.615 | 14 |
| L5 | 10.6 (10) | 0.0793 | −0.524 | −0.297 | 0.381 | 0.998 | −0.943 | 0.112 | 0.383 | 4 |
| L808 | 11.3 (1) | −0.591 | 0.976 | 1.700 | −0.864 | 0.223 | −0.129 | −0.183 | 0.686 | 16 |
| N101 | 10.5 (11) | −0.258 | −1.34 | 0.439 | 0.206 | 0.216 | 0.483 | −0.2 | 0.546 | 10 |
| N988 | 10.4 (14) | 0.557 | −1.66 | 0.215 | −0.314 | 0.398 | −0.064 | 0.417 | 0.666 | 15 |
| Q218 | 11 (4) | −0.052 | 1.031 | 0.582 | 0.456 | 0.142 | −0.353 | −0.923 | 0.451 | 6 |
| W702 | 9.98 (16) | 1.220 | 1.072 | −0.264 | −0.199 | 0.786 | 1.38 | 0.717 | 0.811 | 18 |
| X20 | 10.2 (15) | −0.667 | 0.493 | −1.27 | 0.063 | 0.053 | 0.0875 | 0.92 | 0.529 | 8 |
| X335 | 10.5 (3) | −0.507 | 0.284 | −1.700 | −0.257 | −0.026 | −0.372 | −1.12 | 0.52 | 7 |
| Y23 | 10.6 (9) | −0.0229 | −0.283 | 0.433 | −0.479 | −0.047 | 0.0571 | −0.318 | 0.25 | 1 |
| Y303 | 10.9 (5) | −0.616 | 0.128 | −0.366 | 0.489 | −0.161 | 0.78 | −0.491 | 0.409 | 5 |
| Z958 | 10.5 (12) | −0.404 | 0.391 | −0.024 | 0.162 | −0.014 | 0.317 | 0.083 | 0.3 | 2 |
| Genotype Code | Ear Length (cm) | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | WAASB | rWAASB |
|---|---|---|---|---|---|---|---|---|---|---|
| D605 | 17.8 (12) | −0.683 | 0.636 | −0.569 | 0.241 | −0.815 | 0.591 | 0.477 | 0.6 | 6 |
| D618 | 17.9 (11) | −0.054 | 1.24 | 0.596 | 0.057 | −0.999 | −0.962 | 1.05 | 0.607 | 9 |
| D9 | 18.8 (4) | −0.119 | −1.6 | 1.73 | −0.21 | 0.586 | −0.415 | −0.577 | 0.709 | 15 |
| H110 | 19.2 (2) | −0.792 | −0.912 | −0.256 | 0.35 | 0.019 | −0.342 | 0.0479 | 0.556 | 4 |
| H168 | 17.4 (17) | −0.417 | −0.027 | −0.568 | −0.045 | −0.253 | 0.668 | −0.372 | 0.365 | 1 |
| H321 | 17.4 (16) | 0.254 | 0.194 | −0.581 | 0.114 | −1.26 | 0.439 | −0.232 | 0.465 | 3 |
| H9 | 19.5 (1) | −1.27 | 0.055 | −0.478 | 0.585 | −0.9 | −1.15 | −0.739 | 0.751 | 17 |
| L206 | 18.6 (7) | 1.22 | −2.25 | −1.03 | −0.673 | −0.007 | −0.285 | −0.040 | 0.789 | 19 |
| L5 | 17.7 (13) | 0.852 | 0.914 | −0.277 | 0.51 | 1.48 | 1.22 | 0.178 | 0.703 | 14 |
| L808 | 17.6 (14) | 0.595 | 0.123 | −1.34 | −1.9 | −0.415 | −0.165 | −0.147 | 0.602 | 8 |
| N101 | 18.2 (8) | −1.81 | −0.227 | 0.424 | 0.33 | −0.21 | 0.227 | 1.17 | 0.677 | 10 |
| N988 | 18.1 (9) | 0.717 | −0.255 | −0.020 | 0.734 | 0.007 | 0.583 | 0.621 | 0.439 | 2 |
| Q218 | 18.1 (10) | 0.731 | 1.66 | 0.001 | 0.253 | 0.752 | −1.17 | −1.77 | 0.757 | 18 |
| W702 | 19.0 (3) | 0.808 | −0.056 | 2.43 | −0.156 | −0.653 | 0.267 | −0.263 | 0.679 | 11 |
| X20 | 18.7 (5) | −1.21 | 0.185 | −0.053 | −1 | 1.39 | −1.3 | 0.552 | 0.734 | 16 |
| X335 | 17.5 (15) | −0.756 | 0.532 | 0.22 | −1.16 | 1.39 | 1.05 | 0.342 | 0.682 | 12 |
| Y23 | 18.7 (6) | −0.028 | −0.62 | −0.706 | 2.27 | 0.847 | −0.006 | −0.256 | 0.6 | 7 |
| Y303 | 17.2 (19) | −0.401 | 0.094 | 0.355 | −0.48 | −0.778 | 1.37 | −1.27 | 0.585 | 5 |
| Z958 | 17.3 (18) | 2.36 | 0.312 | 0.12 | 0.177 | −0.175 | −0.617 | 1.22 | 0.699 | 13 |
| Genotype Code | Hundred Seed Weight (g) | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | WAASB | rWAASB |
|---|---|---|---|---|---|---|---|---|---|---|
| D605 | 33.8 (15) | −0.386 | −1.63 | −0.697 | 2.1 | 0.645 | −0.163 | 0.873 | 0.974 | 11 |
| D618 | 35.1 (12) | 0.776 | 0.139 | 1.9 | −0.789 | 0.513 | −1.19 | −0.429 | 0.774 | 4 |
| D9 | 33.4 (17) | −3.5 | −1.21 | 0.951 | −1.13 | −0.325 | 0.563 | 0.314 | 1.29 | 18 |
| H110 | 36.3 (4) | −1.01 | 2.81 | −0.539 | 0.846 | −0.329 | −1.89 | 0.269 | 1.12 | 15 |
| H168 | 30.9 (19) | 2.24 | −2.34 | 1.24 | −0.346 | −2.42 | 1.14 | 1.01 | 1.47 | 19 |
| H321 | 35.7 (9) | 1.48 | 0.784 | 1.82 | −0.202 | 0.186 | −1.22 | −0.837 | 0.934 | 9 |
| H9 | 35.9 (8) | 1.16 | 0.398 | −1.24 | −1.08 | −1.18 | −0.553 | 1.78 | 0.984 | 12 |
| L206 | 35.5 (11) | 0.941 | −0.529 | −1.51 | 0.41 | −0.701 | −1.55 | 0.212 | 0.898 | 8 |
| L5 | 32.7 (18) | −1.61 | −1.5 | −0.094 | 0.945 | −0.447 | −1.47 | −0.333 | 0.942 | 10 |
| L808 | 36.1 (6) | 1.04 | −1.45 | −1.65 | 0.427 | 2.31 | −0.207 | 1.04 | 1.11 | 13 |
| N101 | 34.5 (14) | −0.143 | 1.42 | 0.55 | 3.09 | −2.08 | 1.7 | −0.729 | 1.13 | 16 |
| N988 | 37.6 (1) | 0.111 | 0.168 | 0.667 | 1.42 | 2.93 | 1.42 | −0.112 | 0.814 | 5 |
| Q218 | 35.6 (10) | 0.614 | −0.203 | −1.13 | −1.92 | 0.191 | 0.378 | −0.668 | 0.764 | 3 |
| W702 | 37.1 (3) | −0.473 | 3.12 | 0.843 | −0.791 | 0.419 | 0.873 | 2.52 | 1.15 | 17 |
| X20 | 36.2 (5) | 0.356 | 0.619 | −0.045 | −0.302 | −0.116 | −1.09 | −1.93 | 0.562 | 1 |
| X335 | 34.9 (13) | −0.26 | 0.256 | −2.5 | 0.0338 | −0.59 | 1.22 | −1.08 | 0.737 | 2 |
| Y23 | 33.5 (16) | −3.05 | −1.14 | 0.706 | −0.96 | −0.151 | −0.059 | −0.037 | 1.11 | 14 |
| Y303 | 37.5 (2) | 0.207 | 0.959 | −1.16 | −1.73 | 0.317 | 1.71 | −1.36 | 0.826 | 7 |
| Z958 | 36.0 (7) | 1.5 | −0.666 | 1.89 | −0.0212 | 0.833 | 0.384 | −0.51 | 0.819 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, H.; Gauch, H.G.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Bu, J.; Jiang, X. Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China. Agriculture 2022, 12, 602. https://doi.org/10.3390/agriculture12050602
Yue H, Gauch HG, Wei J, Xie J, Chen S, Peng H, Bu J, Jiang X. Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China. Agriculture. 2022; 12(5):602. https://doi.org/10.3390/agriculture12050602
Chicago/Turabian StyleYue, Haiwang, Hugh G. Gauch, Jianwei Wei, Junliang Xie, Shuping Chen, Haicheng Peng, Junzhou Bu, and Xuwen Jiang. 2022. "Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China" Agriculture 12, no. 5: 602. https://doi.org/10.3390/agriculture12050602