
Lycium barbarum Polysaccharide Inhibits E. coli-Induced Inflammation and Oxidative Stress in Mammary Epithelial Cells of Dairy Cows via SOCS3 Activation and MAPK Suppression

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Chemicals
2.3. Cell Culture Conditions
2.4. Cell Viability
2.5. RNA Extraction and Quantitative RT-PCR
2.6. ROS Determination
2.7. Antioxidant Enzyme Activity
2.8. Immunofluorescence
2.9. Western Blot
2.10. Statistics
3. Results
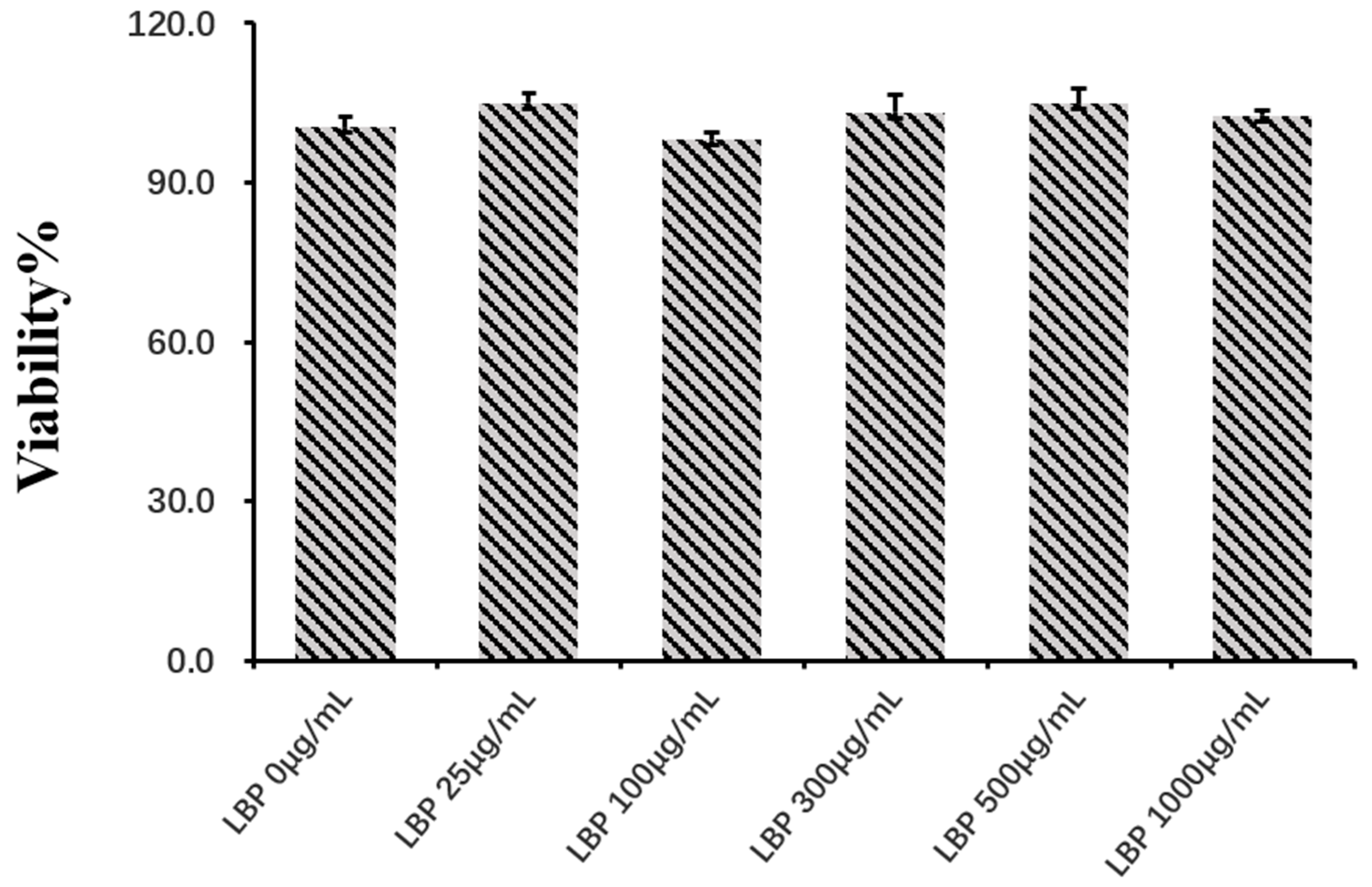
3.1. The Effect of Cell Viability of LBP on pbMEC
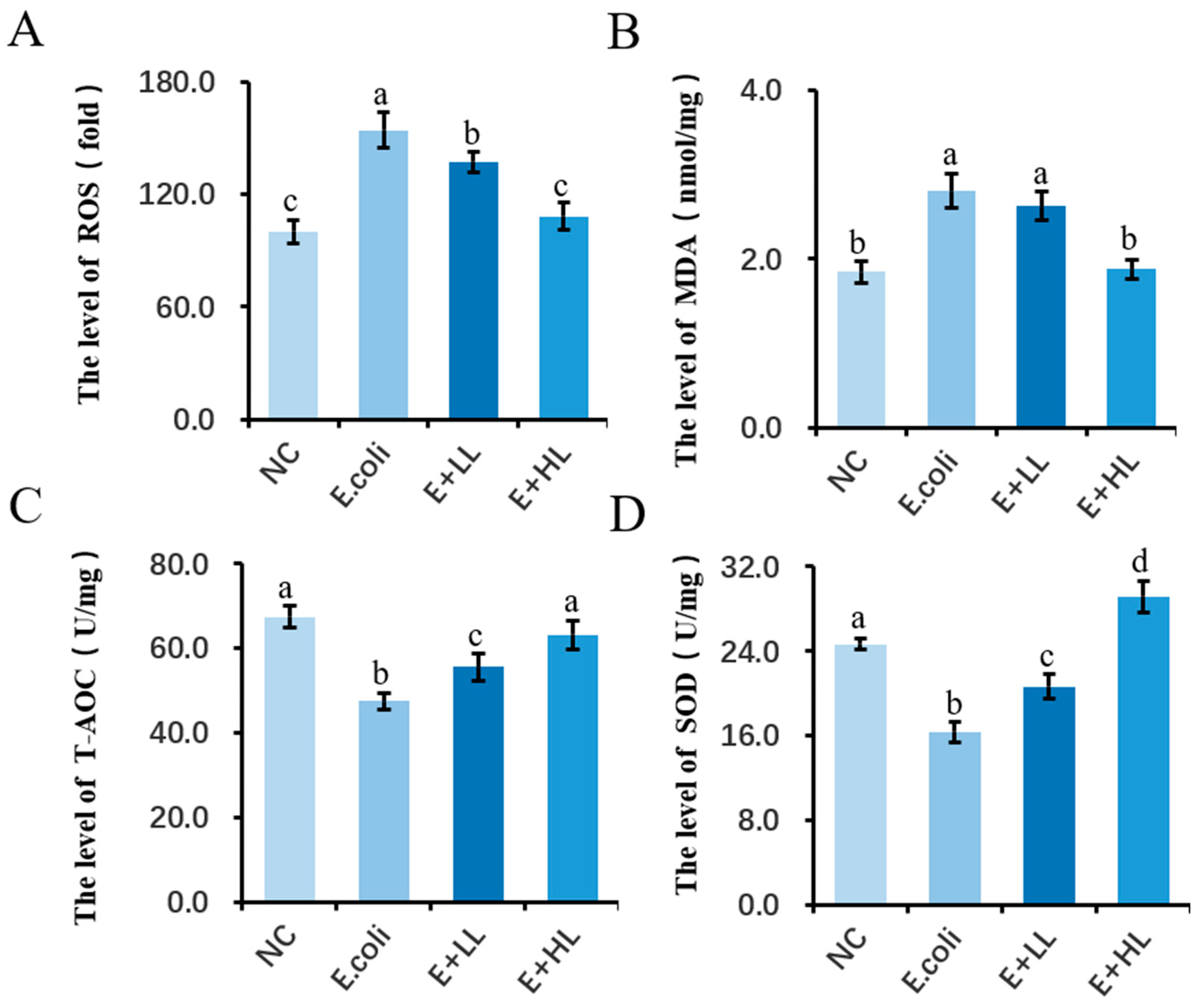
3.2. The Effect of LBP on the Antioxidant Capacity of pbMEC
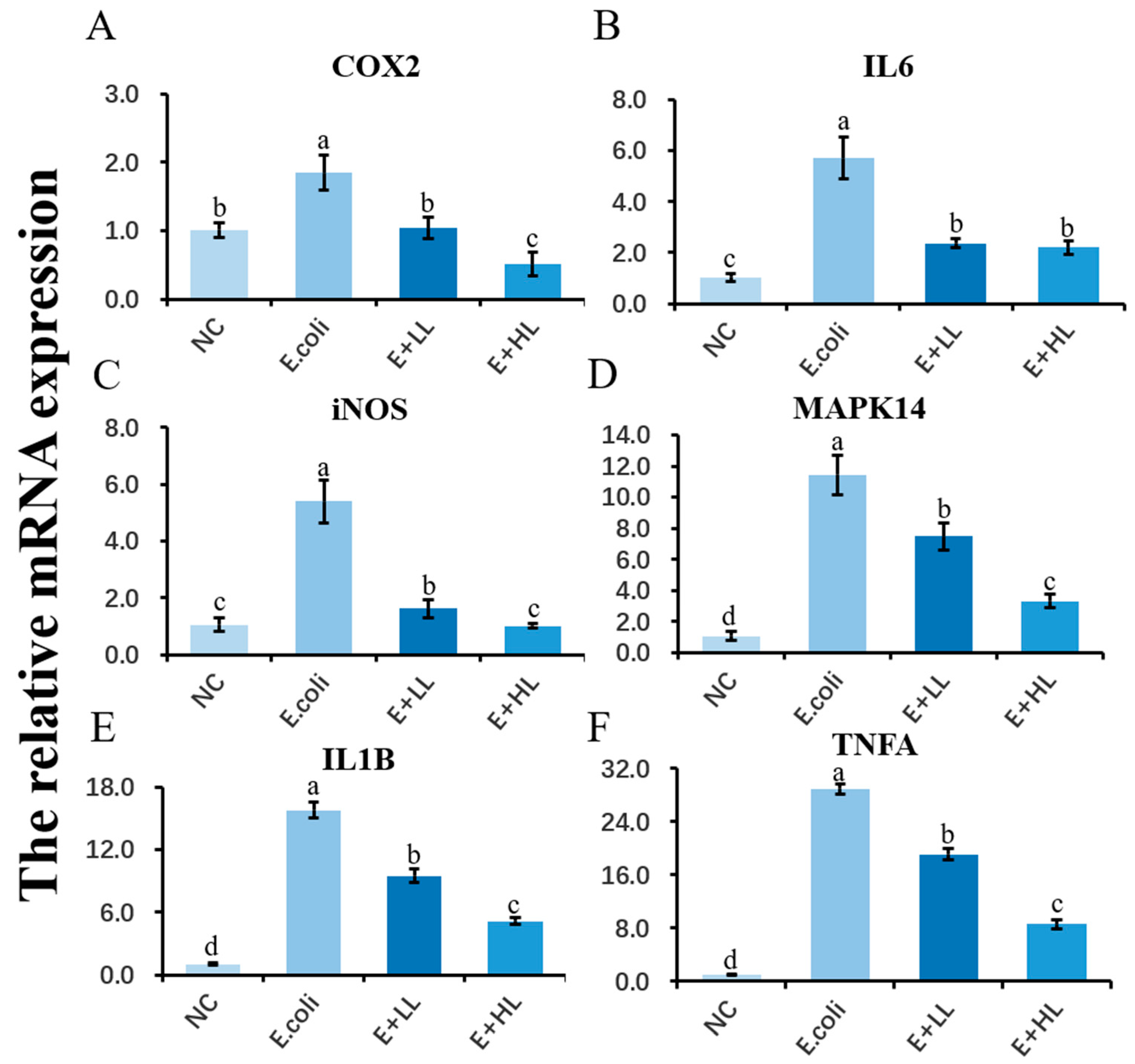
3.3. Effect of LBP on the Expression of Genes Related to Inflammatory Responses in E. coli Stimulated pbMEC
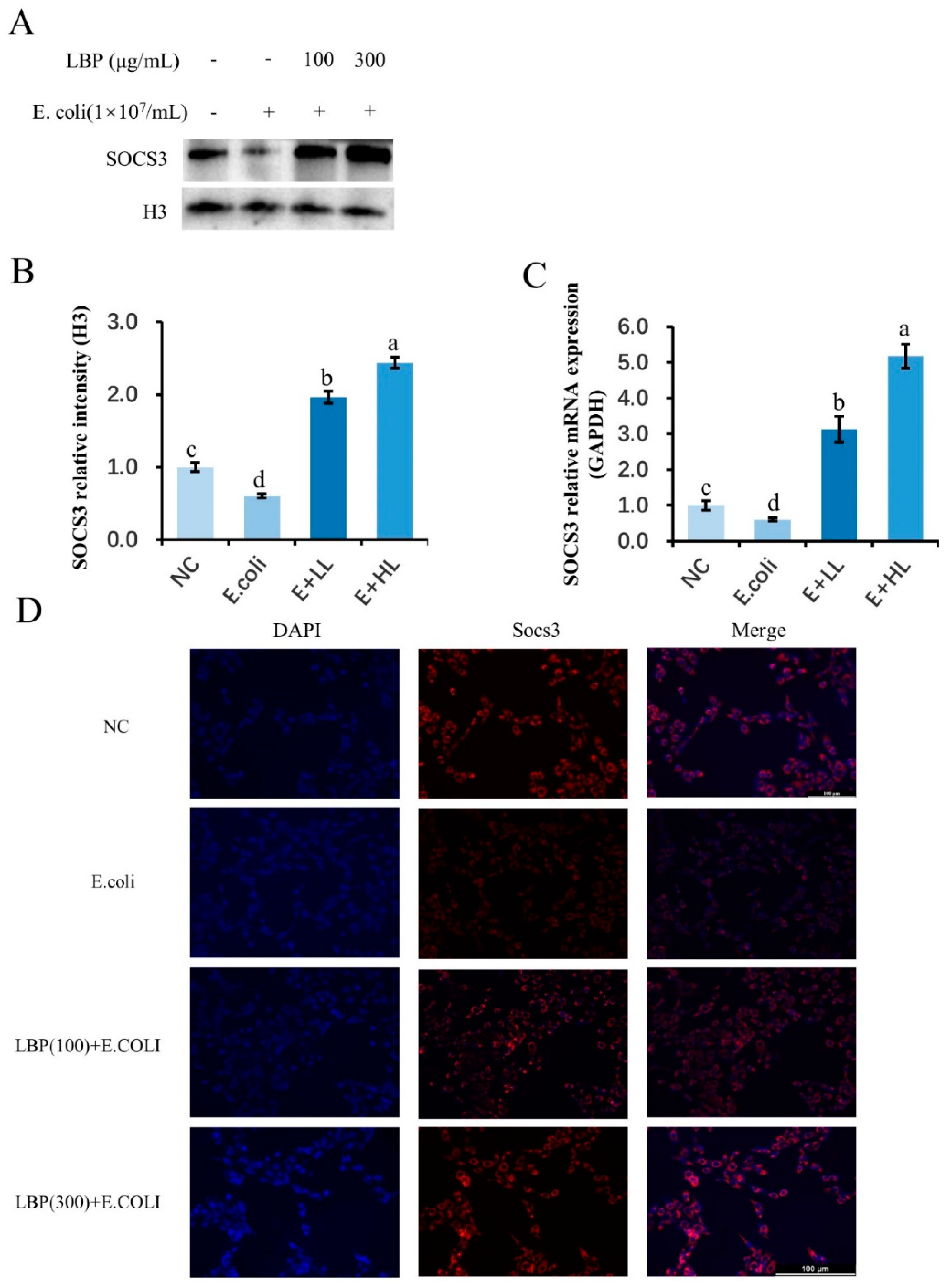
3.4. Pretreatment with LBP Activated SOCS3 in pbMEC Challenged with E. coli
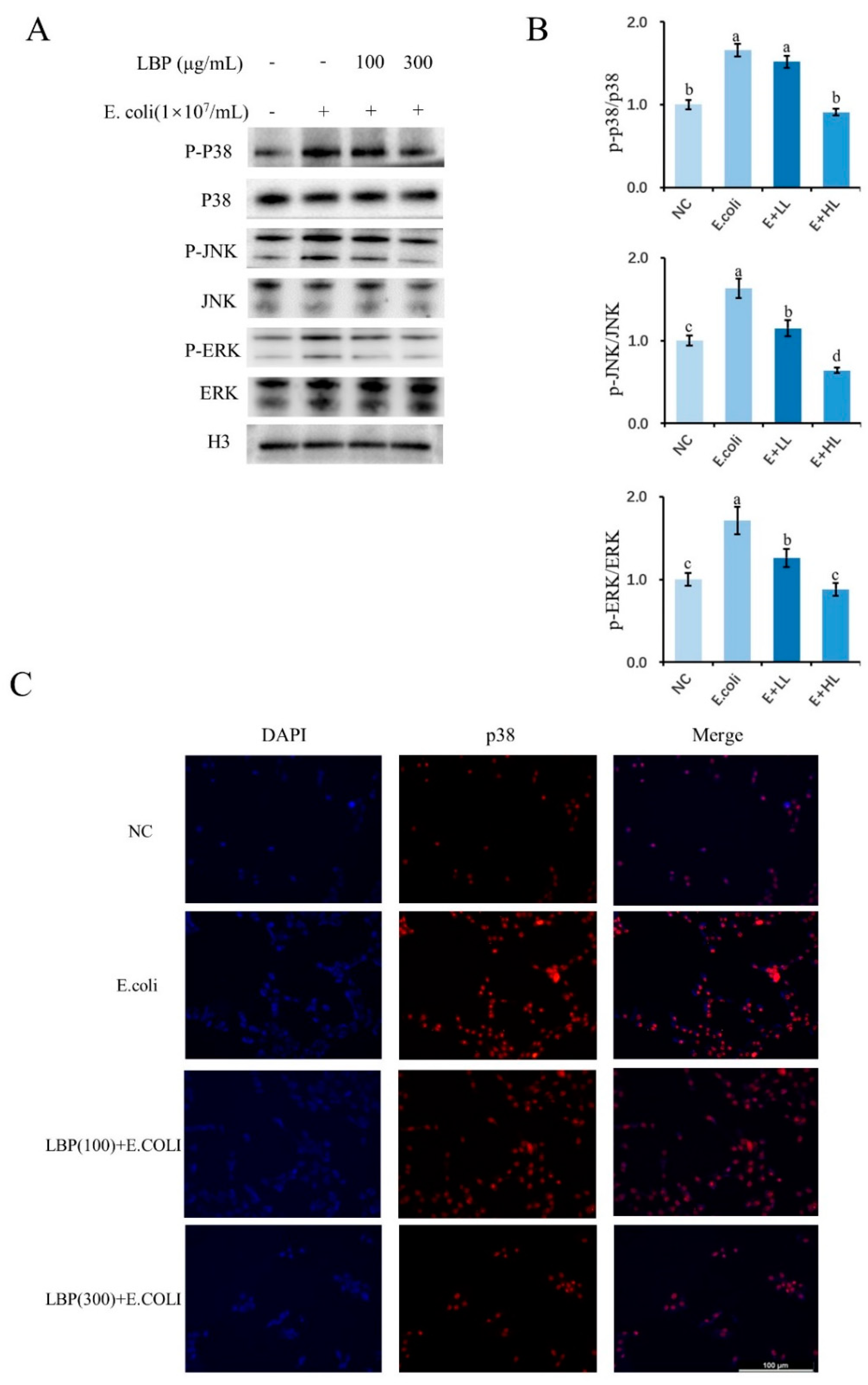
3.5. Effect of LBP on the MAPK Pathway-Related Protein Expression of pbMEC under Different Treatments
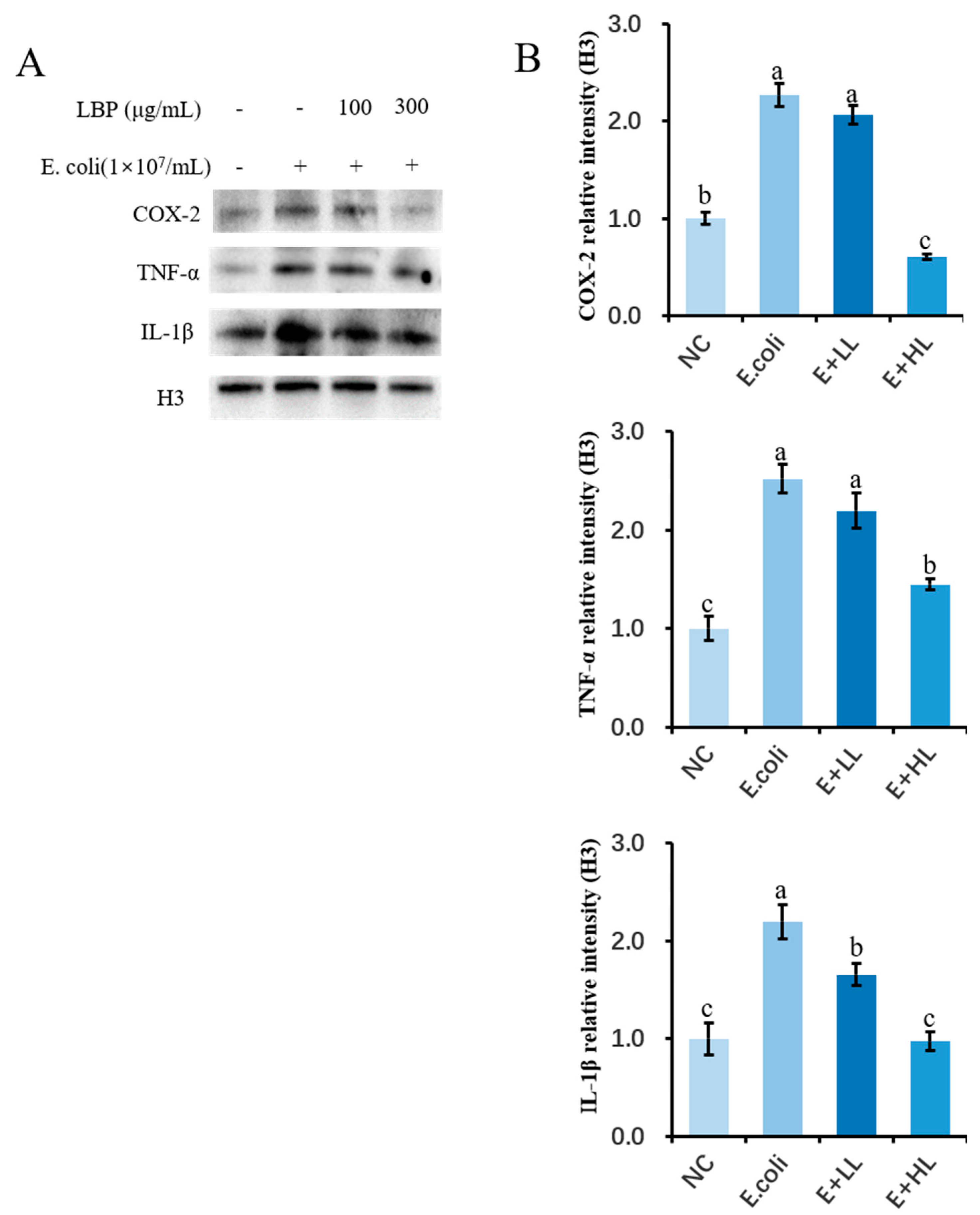
3.6. LBP Pretreatment Inhibits E. coli-Induced Activation of Inflammatory Genes
4. Discussion
4.1. LBP Attenuates the E. coli-Induced Inflammatory and Oxidative Responses in pbMEC
4.2. LBP Inhibits Inflammatory and Oxidative Responses in pbMEC through SOCS3 and MAPK Molecules
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merriman, K.E.; Powell, J.L.; Santos, J.E.P.; Nelson, C.D. Intramammary 25-hydroxyvitamin D 3 treatment modulates innate immune responses to endotoxin-induced mastitis. J. Dairy Sci. 2018, 101, 7593–7607. [Google Scholar] [CrossRef] [PubMed]
- Tomazi, T.; Coura, F.M.; Goncalves, J.L.; Heinemann, M.B.; Santos, M.V. Antimicrobial susceptibility patterns of Escherichia coli phylogenetic groups isolated from bovine clinical mastitis. J. Dairy Sci. 2018, 101, 9406–9418. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Gao, Y.; Xue, Y.; Liu, Y.; Zeng, X.; Cheng, Y.; Ma, J.; Wang, H.; Sun, J.; Wang, Z.; et al. Bacteriophage Cocktails Protect Dairy Cows Against Mastitis Caused by Drug Resistant Escherichia coli Infection. Front. Cell. Infect. Microbiol. 2021, 11, 690377. [Google Scholar] [CrossRef] [PubMed]
- Guerra, S.T.; Paula, C.; Bolaos, C.; Hernandes, R.T.; Ribeiro, M.G. Virulence factors of Escherichia coli: An overview of animal and human infections with emphasis in bovine mastitis. Semin. Cienc. Agrar. 2019, 40, 2087. [Google Scholar] [CrossRef]
- Blum, S.E.; Dan, E.H.; Jacoby, S.; Krifuks, O.; Leitner, G. Physiological response of mammary glands to Escherichia coli infection: A conflict between glucose need for milk production and immune response. Sci. Rep. 2020, 10, 9602. [Google Scholar] [CrossRef]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Health Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Jedrzejczak, M.; Szatkowska, I. Bovine mammary epithelial cell cultures for the study of mammary gland functions. In Vitro Cell. Dev. Biol.-Anim. 2014, 50, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Kehrli, M.E.; Shuster, D.E. Factors Affecting Milk Somatic Cells and Their Role in Health of the Bovine Mammary Gland. J. Dairy Sci. 1994, 77, 619–627. [Google Scholar] [CrossRef]
- Gomes, F.; Henriques, M. Control of Bovine Mastitis: Old and Recent Therapeutic Approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, C.; Wei, Z.; He, X.; Kou, J.; Zhou, E.; Yang, Z.; Fu, Y. Morin suppresses inflammatory cytokine expression by downregulation of nuclear factor-κB and mitogen-activated protein kinase (MAPK) signaling pathways in lipopolysaccharide-stimulated primary bovine mammary epithelial cells. J. Dairy Sci. 2016, 99, 3016–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Han, L.; Li, Y.; Zhao, H.; Zhang, Z.; Zhuang, Y.; Zhong, M.; Wang, Q.; Ma, W.; Wang, Y. Isorhamnetin attenuates TNF-α-induced inflammation, proliferation, and migration in human bronchial epithelial cells via MAPK and NF-κB pathways. Anat Rec (Hoboken). 2021, 304, 901–913. [Google Scholar] [CrossRef]
- Yang, Z.; Yin, R.; Cong, Y.; Yang, Z.; Zhou, E.; Wei, Z.; Liu, Z.; Cao, Y.; Zhang, N. Oxymatrine Lightened the Inflammatory Response of LPS-Induced Mastitis in Mice Through Affecting NF-κB and MAPKs Signaling Pathways. Inflammation 2014, 37, 2047–2055. [Google Scholar] [CrossRef] [PubMed]
- Shu, F.; Junfu, W.; Xiaoqun, X.; Xuemei, C.; Junwen, L.; Qinghong, S.; Meng, L.; Huali, W.; Changsheng, Z. The Expression of SOCS and NF-ϰB p65 in Hypopharyngeal Carcinoma. Iran. J. Public Health 2018, 47, 1874–1882. [Google Scholar]
- Elliott, J.; Johnston, J.A. SOCS: Role in inflammation, allergy and homeostasis. Trends Immunol. 2004, 25, 434–440. [Google Scholar] [CrossRef]
- Souza, J.A.C.d.; Nogueira, A.V.B.; Souza, P.P.C.d.; Kim, Y.J.; Lobo, C.S.; Oliveira, G.J.P.L.d.; Cirelli, J.A.; Garlet, G.P.; Rossa, C.; López-Collazo, E. SOCS3 Expression Correlates with Severity of Inflammation, Expression of Proinflammatory Cytokines, and Activation of STAT3 and p38 MAPK in LPS-Induced Inflammation In Vivo. Mediat. Inflamm. 2013, 2013, 650812. [Google Scholar]
- Delgado-Ortega, M.; Melo, S.; Meurens, F. Expression of SOCS1-7 and CIS mRNA in porcine tissues. Vet. Immunol. Immunopathol. 2011, 144, 493–498. [Google Scholar] [CrossRef]
- Hu, J.; Wang, W.; Hao, Q.; Zhang, T.; Yin, H.; Wang, M.; Zhang, C.; Zhang, C.; Zhang, L.; Zhang, X.; et al. Suppressors of cytokine signalling (SOCS)-1 inhibits neuroinflammation by regulating ROS and TLR4 in BV2 cells. Inflamm. Res. 2020, 69, 27–39. [Google Scholar] [CrossRef]
- Mingzhao, D.; Xinyu, H.; Ling, K.; Baohai, Z.; Chaopu, Z. Lycium barbarum Polysaccharide Mediated the Antidiabetic and Antinephritic Effects in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats via Regulation of NF-κB. BioMed Res. Int. 2016, 2016, 3140290. [Google Scholar]
- Xiao-rui, Z.; Chun-hui, Q.; Jun-ping, C.; Gang, L.; Lin-juan, H.; Zhong-fu, W.; Wen-xia, Z.; Yong-xiang, Z. Lycium barbarum polysaccharide LBPF4-OL may be a new Toll-like receptor 4/MD2-MAPK signaling pathway activator and inducer. Int. Immunopharmacol. 2014, 19, 132–141. [Google Scholar]
- Péter, F.; Ildikó, S.; Hajnalka, L.; Mariann, H.; Sándor, S.; György, P. Association of chemerin with oxidative stress, inflammation and classical adipokines in non-diabetic obese patients. J. Cell. Mol. Med. 2014, 18, 1313–1320. [Google Scholar]
- Qi, B.; Ji, Q.; Wen, Y.; Lian, L.; Guo, X.; Hou, G.; Wang, G.; Zhong, J.; Ferenc, G. Lycium barbarum Polysaccharides Protect Human Lens Epithelial Cells against Oxidative Stress–Induced Apoptosis and Senescence. PLoS ONE 2014, 9, e110275. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Ma, Y.L.; Liu, X.J. Effect of the Lycium barbarum polysaccharides on age-related oxidative stress in aged mice. J. Ethnopharmacol. 2007, 111, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y. Biological activities and potential health benefits of polysaccharides from Poria cocos and their derivatives. Int. J. Biol. Macromol. 2014, 68, 131–134. [Google Scholar] [CrossRef]
- Sargeant, J.M.; Gillespie, J.R.; Oberst, R.D.; Phebus, R.K.; Hyatt, D.R.; Bohra, L.K.; Galland, J.C. Results of a longitudinal study of the prevalence of Escherichia coli O157:H7 on cow-calf farms. Am. J. Vet. Res. 2000, 61, 1375–1379. [Google Scholar] [CrossRef] [Green Version]
- Shuster, D.E.; Kehrli, M.E.; Stevens, M.G. Cytokine production during endotoxin-induced mastitis in lactating dairy cows. Am. J. Vet. Res. 1993, 54, 80–85. [Google Scholar]
- Larson, M.A.; Weber, A.; Mcdonald, T.L. Bovine serum amyloid A3 gene structure and promoter analysis: Induced transcriptional expression by bacterial components and the hormone prolactin. Gene 2006, 380, 104–110. [Google Scholar] [CrossRef]
- Xiao, J.; Xing, F.; Huo, J.; Fung, M.L.; Liong, E.C.; Ching, Y.P.; Xu, A.; Chang, R.; So, K.F.; Tipoe, G.L. Lycium barbarum polysaccharides therapeutically improve hepatic functions in non-alcoholic steatohepatitis rats and cellular steatosis model. Sci. Rep. 2014, 4, 5587. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A.; Nebreda, A. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Liu, H.; Shi, S.; Li, M. Lycium ruthenicum polysaccharide attenuates inflammation through inhibiting TLR4/NF-κB signaling pathway. Int. J. Biol. Macromol. 2014, 67, 330–335. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, F.; Liong, E.C.; So, K.F.; Tipoe, G.L. Lycium barbarum polysaccharides improve hepatic injury through NFkappa-B and NLRP3/6 pathways in a methionine choline deficient diet steatohepatitis mouse model. Int. J. Biol. Macromol. 2018, 120, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Granucci, F. The Family of LPS Signal Transducers Increases: The Arrival of Chanzymes. Immunity 2018, 48, 4–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, Y.; Xu, Y.; Wu, W.; Li, W.; Sun, Y.; Yang, J.; Zhu, Y.; Zhang, C. Tetrandrine Suppresses Lipopolysaccharide-Induced Microglial Activation by Inhibiting NF-κB and ERK Signaling Pathways in BV2 Cells. PLoS ONE 2014, 9, e102522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, G.; Li, Y.; He, M.; Guo, W.; Kan, X.; Xu, D.; Liu, J.; Fu, S. Peiminine Protects against Lipopolysaccharide-Induced Mastitis by Inhibiting the AKT/NF-κB, ERK1/2 and p38 Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 2637. [Google Scholar]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Willson, J.A.; Arienti, S.; Sadiku, P.; Reyes, L.; Coelho, P.; Morrison, T.; Rinaldi, G.; Dockrell, D.H.; Whyte, M.K.B.; Walmsley, S.R. Neutrophil HIF-1α stabilization is augmented by mitochondrial ROS produced via the glycerol 3-phosphate shuttle. Blood 2022, 139, 281–286. [Google Scholar] [CrossRef]
- Tian, X.; Liang, T.; Liu, Y.; Ding, G.; Zhang, F.; Ma, Z. Extraction, Structural Characterization, and Biological Functions of Lycium Barbarum Polysaccharides: A Review. Biomolecules 2019, 9, 389. [Google Scholar] [CrossRef] [Green Version]
- Tuo, X.U.; Ling, H.Y.; Long, J.; Jian-Qin, H.E.; Yang, S.S.; Zhu, Z.M.; Yan, C.Q.; Feng, S.D.; Physiology, D.O. Ameliorative effects and the mechanism of lycium barbarum polysaccharide on insulin resistance of HepG2 cell. Chin. J. Appl. Physiol. 2017, 33, 568–571. [Google Scholar]
- Kwon, K.; Jung, J.; Sahu, A.; Tae, G. Nanoreactor for cascade reaction between SOD and CAT and its tissue regeneration effect. J. Control Release 2022, 344, 160–172. [Google Scholar] [CrossRef]
- Tamiya, T.; Kashiwagi, I.; Takahashi, R.; Yasukawa, H.; Yoshimura, A. Suppressors of cytokine signaling (SOCS) proteins and JAK/STAT pathways: Regulation of T-cell inflammation by SOCS1 and SOCS3. Arter. Thromb Vasc Biol 2011, 31, 980–985. [Google Scholar] [CrossRef] [Green Version]
- Latvala, S.; Miettinen, M.; Kekkonen, R.A.; Korpela, R.; Julkunen, I. Lactobacillus rhamnosus GG and Streptococcus thermophilus induce suppressor of cytokine signalling 3 (SOCS3) gene expression directly and indirectly via interleukin-10 in human primary macrophages. Clin. Exp. Immunol. 2011, 165, 94–103. [Google Scholar] [CrossRef]
- Liu, Z.; Gan, L.; Zhou, Z.; Jin, W.; Sun, C. SOCS3 promotes inflammation and apoptosis via inhibiting JAK2/STAT3 signaling pathway in 3T3-L1 adipocyte. Immunobiology 2015, 220, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ye, L.; Liu, L.; Bian, Y.; Li, Z.; Gao, X.; Li, Q. 14-3-3γ Regulates Lipopolysaccharide-Induced Inflammatory Responses and Lactation in Dairy Cow Mammary Epithelial Cells by Inhibiting NF-κB and MAPKs and Up-Regulating mTOR Signaling. Int. J. Mol. Sci. 2015, 16, 16622–16641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junttila, M.R.; Li, S.P.; Westermarck, J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008, 22, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahra, A.; Vaiva, K.; Volodymyr, S. Dual control of MAPK activities by AP2C1 and MKP1 MAPK phosphatases regulates defence responses in Arabidopsis. J. Exp. Bot. 2022, 73, 2369–2384. [Google Scholar]
- Liu, K.; Liu, E.; Lin, L.; Hu, Y.; Yuan, Y.; Xiao, W. L-Theanine mediates the p38MAPK signaling pathway to alleviate heat-induced oxidative stress and inflammation in mice. Food Funct. 2022, 13, 2120–2130. [Google Scholar] [CrossRef]
- Liu, M.; Song, S.; Li, H.; Jiang, X.; Yin, P.; Wan, C.; Liu, X.; Liu, F.; Xu, J. The protective effect of caffeic acid against inflammation injury of primary bovine mammary epithelial cells induced by lipopolysaccharide. J. Dairy Sci. 2014, 97, 2856–2865. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Zhu, H.; Zhao, J.; Wu, X.; Lu, X.; Xu, T.; Yang, Z. Lycium barbarum Polysaccharide Inhibits E. coli-Induced Inflammation and Oxidative Stress in Mammary Epithelial Cells of Dairy Cows via SOCS3 Activation and MAPK Suppression. Agriculture 2022, 12, 598. https://doi.org/10.3390/agriculture12050598
Liu R, Zhu H, Zhao J, Wu X, Lu X, Xu T, Yang Z. Lycium barbarum Polysaccharide Inhibits E. coli-Induced Inflammation and Oxidative Stress in Mammary Epithelial Cells of Dairy Cows via SOCS3 Activation and MAPK Suppression. Agriculture. 2022; 12(5):598. https://doi.org/10.3390/agriculture12050598
Chicago/Turabian StyleLiu, Run, Hao Zhu, Jingwen Zhao, Xinyue Wu, Xubin Lu, Tianle Xu, and Zhangping Yang. 2022. "Lycium barbarum Polysaccharide Inhibits E. coli-Induced Inflammation and Oxidative Stress in Mammary Epithelial Cells of Dairy Cows via SOCS3 Activation and MAPK Suppression" Agriculture 12, no. 5: 598. https://doi.org/10.3390/agriculture12050598