
Diagnostic Use of Serum Amyloid A in Dairy Cattle


Abstract
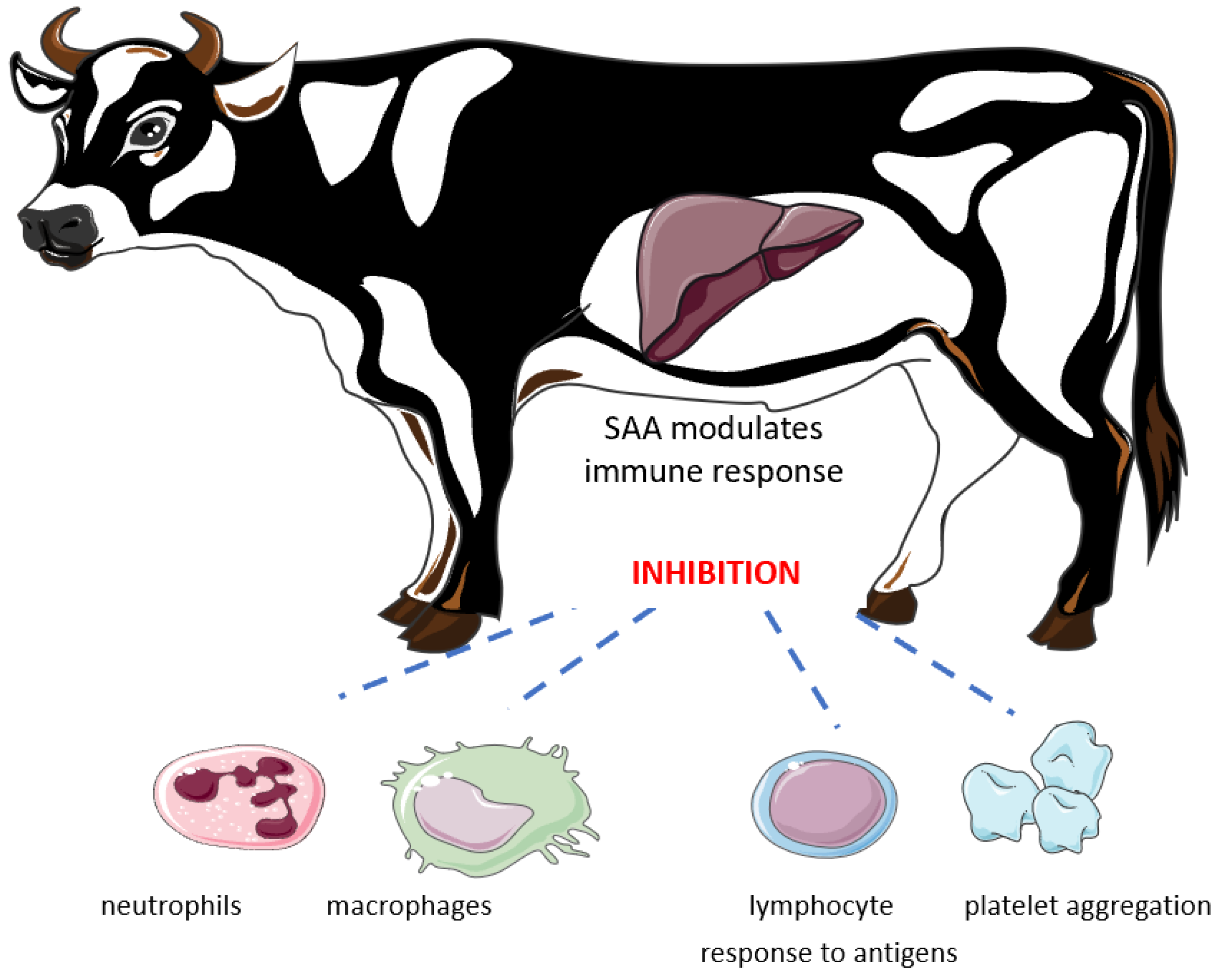
:1. Introduction
2. SAA Concentrations in Healthy Cattle—Impact of Nutrition and Stress
3. SAA as a Diagnostic Tool in Metabolic Disorders in Adult Dairy Cows
4. SAA as a Diagnostic Tool in Gastrointestinal Disorders in Adult Dairy Cows
5. SAA as a Diagnostic Tool in Respiratory Tract Infections
6. SAA as a Diagnostic Tool in Parasitic Infections
7. SAA in Reproduction
8. SAA in Limb Diseases
9. SAA in Mastitis
10. SAA in Calves Diseases
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cray, C.; Zaias, J.; Altman, N.H. Acute phase response in animals: A review. Comp. Med. 2009, 6, 517–526. [Google Scholar]
- Eckersall, P.D.; Bell, R. Acute phase proteins: Biomarkers of infection and inflammation in veterinary medicine. Vet. J. 2010, 185, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Guzelbektes, H.; Sen, I.; Ok, M. Serum amyloid A and haptoglobin concentrations and liver fat percentage in lactating dairy cows with abomasal displacement. J. Vet. Intern. Med. 2010, 24, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suojala, L.; Orro, T.; Järvinen, H. Acute phase response in two consecutive experimentally induced E. coli intramammary infections in dairy cows. Acta Vet. Scand. 2008, 50, 18. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.C.; Waterston, M.; Hastie, P. The major acute phase proteins of bovine milk in a commercial dairy herd. BMC Vet. Res. 2015, 11, 207. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, D.G.; Dunn, S.M.; Ametaj, B.N. Feeding high proportions of barley grain stimulates an inflammatory response in dairy cows. J. Dairy Sci. 2008, 91, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Lomborg, S.R.; Nielsen, L.R.; Heegaard, P.M. Acute phase proteins in cattle after exposure to complex stress. Vet. Res. Commun. 2008, 7, 575–582. [Google Scholar] [CrossRef]
- Du, X.; Chen, L.; Huang, D. Elevated Apoptosis in the Liver of Dairy Cows with Ketosis. Cell. Physiol. Biochem. 2017, 2, 568–578. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Li, X. Inflammatory mechanism of Rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Bradford, B.J.; Bobe, G. Strong Relationships between Mediators of the Acute Phase Response and Fatty Liver in Dairy Cows. Can. J. Anim. Sci. 2005, 85, 165–175. [Google Scholar] [CrossRef]
- Nazifi, S.; Rezakhani, A.; Moaddeli, A. Study on diagnostic values of haptoglobin and serum amyloid A concentration in bovine heart diseases. Comp. Clin. Pathol. 2009, 18, 47–51. [Google Scholar] [CrossRef]
- Espinosa, J.; de la Morena, R.; Benavides, J. Assessment of Acute-Phase Protein Response Associated with the Different Pathological Forms of Bovine Paratuberculosis. Animals 2020, 10, 1925. [Google Scholar] [CrossRef] [PubMed]
- Sharifiyazdia, H.; Nazifi, S.; Nikseresht, K. Evaluation of Serum Amyloid A and Haptoglobin in Dairy Cows Naturally Infected with Brucellosis. J. Bacteriol. Parasitol. 2012, 3, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, S.; Mohammadi, V.; Esmaeilnejad, B. Evaluation of some acute phase proteins in cattle naturally infected with Babesia bigemina. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101642. [Google Scholar] [CrossRef] [PubMed]
- Nazifi, S.; Razavi, S.M.; Kaviani, F.; Rakhshandehroo, E. Acute phase response in cattle infected with Anaplasma marginale. Vet. Microbiol. 2012, 155, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, R.; Pieper, L.; Gonzalez-Grajales, L.A. Evaluation of different acute-phase proteins for herd health diagnostics in early postpartum Holstein Friesian dairy cows. J. Dairy Res. 2021, 88, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Kaya, S.; Merhan, O.; Kacar, C. Determination of ceruloplasmin, some other acute phase proteins, and biochemical parameters in cows with endometritis. Vet. World 2016, 10, 1056–1062. [Google Scholar] [CrossRef]
- Brodzki, P.; Kostro, K.; Brodzki, A. The concentrations of inflammatory cytokines and acute-phase proteins in the peripheral blood and uterine washings in cows with pyometra. Reprod. Domest. Anim. 2015, 3, 417–422. [Google Scholar] [CrossRef]
- Ilievska, K.; Atanasov, B.; Dovenski, T. Acute phase proteins—As indicators of claw diseases in dairy cattle. Mac. Vet. Rev. 2019, 42, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Tóthová, C.; Nagy, O.; Kovác, G. Changes in the concentrations of selected acute phase proteins and variables of energetic profile in dairy cows after parturition. J. Appl. Anim. Res. 2014, 3, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Cardoso, F.C.; Pineda, A. Grain challenge affects systemic and hepatic molecular biomarkers of inflammation, stress, and metabolic responses to a greater extent in Holstein than Jersey cows. J. Dairy Sci. 2017, 11, 9153–9162. [Google Scholar] [CrossRef]
- Zebeli, Q.; Metzler-Zebeli, B.U.; Ametaj, B.N. Meta-analysis reveals threshold level of rapidly fermentable dietary concentrate that triggers systemic inflammation in cattle. J. Dairy Sci. 2012, 5, 2662–2672. [Google Scholar] [CrossRef] [Green Version]
- Al-Qaisi, M.; Horst, E.A.; Mayorga, E.J. Effects of a Saccharomyces cerevisiae fermentation product on heat-stressed dairy cows. J. Dairy Sci. 2020, 10, 9634–9645. [Google Scholar] [CrossRef]
- Coskun, A.; Guzelbektes, H.; Simsek, A. Haptoglobin and SAA concentrations and enzyme activities in bronchoalveolar lavage fluids from calves with bronchopneumonia. Rev. Méd. Vét. 2012, 12, 615–620. [Google Scholar]
- Sevimli, A.; Sevimli, F.K.; Şeker, E. Acute-phase responses in cattle infected with hydatid cysts and microbial agents. J. Helminthol. 2015, 4, 471–479. [Google Scholar] [CrossRef]
- Brodzki, P.; Kostro, K.; Brodzki, A. Inflammatory cytokines and acute-phase proteins concentrations in the peripheral blood and uterus of cows that developed endometritis during early postpartum. Theriogenology 2015, 84, 11–18. [Google Scholar] [CrossRef]
- Alsemgeest, S.P.; Lambooy, I.E.; Wierenga, H.K. Influence of physical stress on the plasma concentration of serum amyloid-A (SAA) and haptoglobin (Hp) in calves. Vet. Q. 1995, 17, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Adnane, M.; Chapwanya, A.; Kaidi, R. Profiling inflammatory biomarkers in cervico-vaginal mucus (CVM) postpartum: Potential early indicators of bovine clinical endometritis? Theriogenology 2017, 103, 117–122. [Google Scholar] [CrossRef]
- Giannetto, C.; Fazio, F.; Casella, S. Acute phase protein response during road transportation and lairage at a slaughterhouse in feedlot beef cattle. J. Vet. Med. Sci. 2011, 11, 1531–1534. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, F.; Oguejiofor, C.F. Endometrial expression of the acute phase molecule SAA is more significant than HP in reflecting the severity of endometritis. Res. Vet. Sci. 2018, 121, 130–133. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D. Alterations in innate immunity reactants and carbohydrate and lipid metabolism precede occurrence of metritis in transition dairy cows. Res. Vet. Sci. 2016, 104, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.A.; Jean, S.G.; Steiner, A. Lameness. In Bovine Surgery and Lameness, 2nd ed.; Blackwell Publishing: Hoboken, NJ, USA, 2005; pp. 198–258. [Google Scholar]
- Zhang, G.; Hailemariam, D.; Dervishi, E. Alterations of Innate Immunity Reactants in Transition Dairy Cows before Clinical Signs of Lameness. Animals 2015, 3, 717–747. [Google Scholar] [CrossRef] [PubMed]
- Bagga, A.; Randhawa, S.S.; Sharma, S.; Bansal, B.K. Acute phase response in lame crossbred dairy cattle. Vet. World 2016, 11, 1204–1208. [Google Scholar] [CrossRef] [PubMed]
- Jawor, S.; Steiner, T.; Stefaniak, W. Determination of selected acute phase proteins during the treatment of limb diseases in dairy cows. Vet. Med. 2008, 4, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Akerstedt, M.; Waller, K.P.; Larsen, L.B. Relationship between haptoglobin and serum amyloid A in milk and milk quality. Int. Dairy J. 2008, 6, 669–674. [Google Scholar] [CrossRef] [Green Version]
- McDonald, T.L.; Larson, M.A.; Mack, D.R. Elevated extrahepatic expression and secretion of mammary-associated serum amyloid A 3 (M-SAA3) into colostrum. Vet. Immunol. Immunopathol. 2001, 83, 203–211. [Google Scholar] [CrossRef]
- Mack, D.R.; McDonald, T.L.; Larson, M.A. The conserved TFLK motif of mammary-associated serum amyloid A3 is responsible for up-regulation of intestinal MUC3 mucin expression in vitro. Pediatr. Res. 2003, 53, 137–142. [Google Scholar] [CrossRef]
- Otsuka, M.; Sugiyama, M.; Ito, T. Diagnostic utility of measuring serum amyloid A with a latex agglutination turbidimetric immunoassay in bovine mastitis: Comparison with haptoglobin and alpha 1 acid glycoprotein. J. Vet. Med. Sci. 2021, 83, 329–332. [Google Scholar] [CrossRef]
- Szczubiał, M.; Dabrowski, R.; Kankofer, M.; Bochniarz, M.; Komar, M. Concentration of serum amyloid A and ceruloplasmin activity in milk from cows with subclinical mastitis caused by different pathogens. Pol. J. Vet. Sci. 2012, 2, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Bochniarz, M.; Szczubiał, M.; Brodzki, P.; Krakowski, L.; Dąbrowski, R. Serum amyloid A as an marker of cow ̀ s mastitis caused by Streptococcus sp. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101498. [Google Scholar] [CrossRef]
- Tothova, C.; Nagy, Y.; Naygova, V. Changes in the concentrations of acute phase proteins in calves during the first months of life. Acta Vet.-Beograd. 2015, 65, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Dudek, K.; Bednarek, D.; Ayling, R.D. Stimulation and analysis of the immune response in calves from vaccinated pregnant cows. Res. Vet. Sci. 2014, 97, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Prohl, A.; Schroedl, W.; Rhode, H. Acute phase proteins as local biomarkers of respiratory infection in calves. BMC Vet. Res. 2015, 11, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orro, T.; Pohjanvirta, T.; Rikula, U. Acute phase protein changes in calves during an outbreak of respiratory disease caused by bovine respiratory syncytial virus. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 23–29. [Google Scholar] [CrossRef]
- Joshi, V.; Gupta, V.K.; Bhanuprakash, A.G. Haptoglobin and serum amyloid A as putative biomarker candidates of naturally occurring bovine respiratory disease in dairy calves. Microb. Pathog. 2018, 116, 33–37. [Google Scholar] [CrossRef]
- Chae, J.B.; Park, J.; Jung, S.H. Acute phase response in bovine coronavirus positive post-weaned calves with diarrhea. Acta Vet. Scand. 2019, 61, 36. [Google Scholar] [CrossRef]
- Tsukano, K.; Shimamori, T.; Fukuda, T. Serum iron concentration as a marker of inflammation in young cows that underwent dehorning operation. J. Vet. Med. Sci. 2019, 4, 626–628. [Google Scholar] [CrossRef] [Green Version]
- Jawor, P.; Mee, J.F.; Stefaniak, T. Perinatal immuno/inflammatory responses in the presence or absence of bovine fetal infection. BMC Vet. Res. 2018, 14, 322. [Google Scholar] [CrossRef]
- Jawor, P.; Stefaniak, T.; Mee, J.F. Immune and inflammatory biomarkers in cases of bovine perinatal mortality with and without infection in utero. J. Dairy Sci. 2017, 100, 1408–1416. [Google Scholar] [CrossRef] [Green Version]
- Trela, M.; Witkowska-Piłaszewicz, O.; Domańska, D.; Kaczmarek, M.M.; Pawliński, B.; Gajewski, Z.; Domino, M. The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle. Int. J. Mol. Sci. 2022, 23, 1235. [Google Scholar] [CrossRef]


{kind=link}
| Diseases/Environmental Conditions | SAA Reported Values/Fold Increase [mg/L] (Reference Values: 0–70 Blood, Milk 0.6–50) [3,5] |
|---|---|
| high grain concentration dry matter (30–45%) | 19–46 [6] |
| heat stress | 4 fold [7] |
| long-term transportation (4–6 h) | 1–2 fold [7] |
| ketosis | 89.2 [8] |
| subacute ruminal acidosis | 313.9 (at least one fold) [9] |
| fatty liver | 80 [10] |
| abomasal displacement | >126 [3] |
| traumatic reticuloperitonitis | 159 [11] |
| paratuberculosis | one fold [12] |
| brucellosis | 123.75 [13] |
| babesiosis | mild 169.8; severe 183.4 [14] |
| Anaplasma marginale | 167–180 [15] |
| one week postpartum | 94 [16] |
| endometritis | low 20.25; mild 28.17; severe 34.62 [17] |
| pyometra | 62.5 [18] |
| acute laminitis | 281.26–590 [19] |
| mastitis | 447.9 (milk 1315.9) [4] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trela, M.; Domańska, D.; Witkowska-Piłaszewicz, O. Diagnostic Use of Serum Amyloid A in Dairy Cattle. Agriculture 2022, 12, 459. https://doi.org/10.3390/agriculture12040459
Trela M, Domańska D, Witkowska-Piłaszewicz O. Diagnostic Use of Serum Amyloid A in Dairy Cattle. Agriculture. 2022; 12(4):459. https://doi.org/10.3390/agriculture12040459
Chicago/Turabian StyleTrela, Michał, Dominika Domańska, and Olga Witkowska-Piłaszewicz. 2022. "Diagnostic Use of Serum Amyloid A in Dairy Cattle" Agriculture 12, no. 4: 459. https://doi.org/10.3390/agriculture12040459