
Does a Soybean Intercrop Increase Nodule Number, N Uptake and Grain Yield of the Followed Main Crop Soybean?

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Site Description
2.2. Site Conditions
2.3. Determination of Yield and Nutrient Analysis
2.4. Statistical Analysis
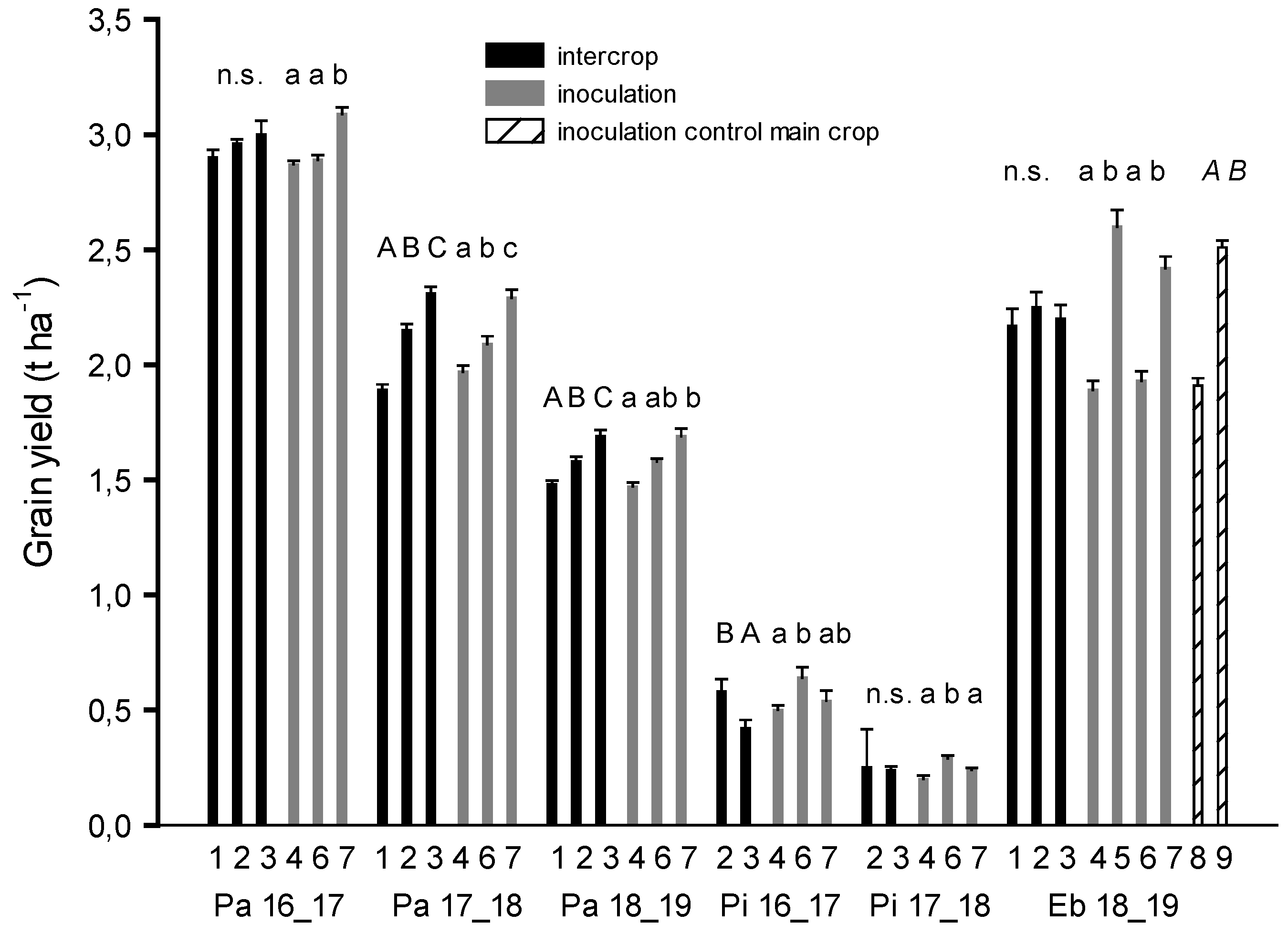
3. Results
3.1. Evaluation of Intercrop
3.2. Evaluation of Main Crop Soybean
4. Discussion
4.1. Effects of Intercropping with Soybeans on Nodule Development
4.2. Effect of Intercropping on the Main Crop Soybean
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Main Crop Soybean | Intercrop Buckwheat/Soybean | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield (t ha−1) | Grain N (%) | No. of Nodules per Plant | Weight of Nodules per Plant (mg) | Nodules_N (%) | Nodules_S* (%) | No. of Nodules per Plant m.r. | No. of Nodules per Plant l.r. | SPAD | Shoot_Mg (%) | Shoot_Na (%) | Shoot_S* (%) | Shoot_Ca (%) | Shoot_K (%) | Shoot_P (%) | DM (t ha−1) | Weed DM (t ha−1) | Weed_N (%) | B_N (%) | S_N (%) | S_N (kg ha−1) | No. of Nodules per Plant | Weight of Nodules per Plant (mg) | Nodules_N (%) | Nodules_S*(%) | ||
| Inoculation | 0.00 | 0.00 | 0.00 | 0.04 | 0.06 | 0.04 | 0.01 | 0.57 | 0.99 | 0.58 | 0.47 | 0.37 | 0.29 | 0.88 | 0.00 * | 0.00 | 0.85 | 0.93 | ||||||||
| Pi 16_17 N = 24 | Intercrop | 0.03 | 0.03 | 0.76 | 0.48 | 0.47 | 0.62 | 0.93 | 0.76 | 0.18 | 0.26 | 0.00 | 0.01 | 0.91 | 0.07 | 0.04 * | 0.20 | 0.23 | 0.83 | |||||||
| Inoculation × Intercrop | 0.25 | 0.59 | 0.52 | 0.04 | 0.17 | 0.94 | 0.55 | 0.38 | 0.24 | 0.43 | 0.37 | 0.05 | 0.36 | 0.53 | 0.24 | 0.67 | 0.06 | 0.91 | ||||||||
| Inoculation | 0.02 | 0.00 | 0.00 | 0.02 | 0.17 | 0.11 | 0.28 | 0.85 | 0.59 | 0.68 | 0.68 | 0.08 | 0.04 | 0.01 | 0.21 | 0.64 | 0.92 | 0.00 | 0.00 | 0.00 | 0.00 * | 0.34 | 0.06 | |||
| Pi 17_18 N = 24 | Intercrop | 0.65 | 0.28 | 0.12 | 0.08 | 0.62 | 0.49 | 0.41 | 0.50 | 0.91 | 0.32 | 0.17 | 0.78 | 0.88 | 0.00 | 0.51 | 0.59 | 0.77 | 0.39 | 0.00 | 0.83 | 0.58 * | 0.44 | 0.75 | ||
| Inoculation × Intercrop | 0.14 | 0.21 | 0.39 | 0.13 | 0.90 | 0.68 | 0.70 | 0.96 | 0.68 | 0.20 | 0.78 | 0.68 | 0.11 | 0 | 0.79 | 0.65 | 0.82 | 0.28 | 0.03 | 0.99 | 0.88 | 0.07 | 0.19 | |||
| Inoculation | 0.00 | 0.00 | 0.00 | 0.00 * | 0.00 | 0.00 * | 0.87 | 0.99 | 0.75 | 0.35 | 0.12 | 0.74 | 0.93 * | 0.45 | 0.42 | 0.33 | 0.31 | 0.02 | 0.73 | 0.28 | ||||||
| Eb 18_19 N = 56 | Intercrop | 0.89 | 0.63 | 0.81 | 0.95 * | 0.62 | 0.86 * | 0.37 | 0.83 | 0.83 | 0.01 | 0.12 | 0.15 | 0.00 * | 0.43 | 0.38 | 0.18 | 0.83 | 0.39 | 0.82 | 0.70 | |||||
| Inoculation × Intercrop | 0.67 | 0.94 | 0.48 | 0.88 | 0.11 | 0.71 | 0.66 | 0.83 | 0.17 | 0.59 | 0.22 | 0.88 | 0.55 | 0.11 | 0.50 | 0.82 | 0.40 | 0.85 | 0.71 | 0.12 | 0.76 | |||||
| Inoculation | 0.00 | 0.53 | 0.28 | 0.93 | 0.29 | 0.66 | 0.58 | 0.21 | ||||||||||||||||||
| Pa 16_17 N = 36 | Intercrop | 0.07 | 0.65 | 0.54 | 0.47 | 0.00 | 0.09 | 0.96 | 0.00 | |||||||||||||||||
| Inoculation × Intercrop | 0.10 | 0.98 | 0.79 | 0.94 | 0.79 | 0.14 | 0.15 | 0.09 | ||||||||||||||||||
| Inoculation | 0.00 | 0.43 | 0.22 | 0.29 | 0.08 | 0.02 | 0.38 | 0.00 | 0.00 | 0.02 | 0.86 | |||||||||||||||
| Pa 17_18 N = 36 | Intercrop | 0.00 | 0.00 | 0.00 | 0.05 | 0.04 | 0.19 | 0.00 | 0.02 | 0.00 | 0.01 | 0.43 | ||||||||||||||
| Inoculation × Intercrop | 0.56 | 0.36 | 0.07 | 0.28 | 0.29 | 0.09 | 0.77 | 0.22 | 0.02 | 0.05 | 0.59 | |||||||||||||||
| Inoculation | 0.01 | 0.25 | 0.96 | 0.07 | 0.84 | 0.94 | 0.00 | 0.00 * | 0.00 | 0.00 | 0.81 | |||||||||||||||
| Pa 18_19 N = 36 | Intercrop | 0.00 | 0.00 | 0.00 | 0.43 | 0.26 | 0.01 | 0.00 | 0.04 * | 0.83 | 0.83 | 0.39 | ||||||||||||||
| Inoculation × Intercrop | 0.08 | 0.04 | 0.2 | 0.75 | 0.78 | 0.1 | 0.05 | 0.09 | 0.94 | 0.08 | 0.58 | |||||||||||||||
References
- Li, L.; Sun, J.; Zhang, F.; Li, X.; Yang, S.; Rengel, Z. Wheat/maize or wheat/soybean strip intercropping. Field Crop. Res. 2001, 71, 123–137. [Google Scholar] [CrossRef]
- Bedoussac, L.; Journet, E.-P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Maitra, S.; Hossain, A.; Brestic, M.; Skalicky, M.; Ondrisik, P.; Gitari, H.; Brahmachari, K.; Shankar, T.; Bhadra, P.; Palai, J.B.; et al. Intercropping—A Low Input Agricultural Strategy for Food and Environmental Security. Agronomy 2021, 11, 343. [Google Scholar] [CrossRef]
- Kivelitz, H. Die Richtige Zwischenfrucht: Auswahl von Zwischenfrüchten unter Berücksichtigung von Fruchtfolge und Saatzeit. Available online: https://www.landwirtschaftskammer.de/riswick/versuche/pflanzenbau/zwischenfruechte/veroeffentlichungen/Zwischenfruechte_Fruchtfolgen.pdf (accessed on 20 January 2021).
- Carruthers, K.; Prithiviraj, B.; Fe, Q.; Cloutier, D.; Martin, R.; Smith, D. Intercropping corn with soybean, lupin and forages: Yield component responses. Eur. J. Agron. 2000, 12, 103–115. [Google Scholar] [CrossRef]
- Redfearn, D.D.; Buxton, D.R.; Devine, T.E. Sorghum Intercropping Effects on Yield, Morphology, and Quality of Forage Soybean. Crop Sci. 1999, 39, 1380–1384. [Google Scholar] [CrossRef]
- Pal, U.R.; Oseni, T.O.; Norman, J.C. Effect of Component Densities on the Productivity of Soybean/Maize and Soybean/Sorghum Intercrop. J. Agron. Crop Sci. 1993, 170, 66–70. [Google Scholar] [CrossRef]
- Li, X.; Mu, Y.; Cheng, Y.; Liu, X.; Nian, H. Effects of intercropping sugarcane and soybean on growth, rhizosphere soil microbes, nitrogen and phosphorus availability. Acta Physiol. Plant. 2013, 35, 1113–1119. [Google Scholar] [CrossRef]
- Biszczak, W.; Różyło, K.; Kraska, P. Yielding parameters, nutritional value of soybean seed and weed infestation in relay-strip intercropping system with buckwheat. Acta Agric. Scand. Plant Soil 2020, 70, 640–647. [Google Scholar] [CrossRef]
- Cheriere, T.; Lorin, M.; Corre-Hellou, G. Species choice and spatial arrangement in soybean-based intercropping: Levers that drive yield and weed control. Field Crops Res. 2020, 256, 107923. [Google Scholar] [CrossRef]
- Obaton, M.; Bouniols, A.; Piva, G.; Vadez, V. Are Bradyrhizobium japonicum stable during a long stay in soil? Plant Soil 2002, 245, 315–326. [Google Scholar] [CrossRef]
- Griebsch, A.; Matschiavelli, N.; Lewandowska, S.; Schmidtke, K. Presence of Bradyrhizobium sp. under Continental Conditions in Central Europe. Agriculture 2020, 10, 446. [Google Scholar] [CrossRef]
- Narożna, D.; Pudełko, K.; Króliczak, J.; Golińska, B.; Sugawara, M.; Mądrzak, C.J.; Sadowsky, M.J. Survival and Competitiveness of Bradyrhizobium japonicum Strains 20 Years after Introduction into Field Locations in Poland. Appl. Environ. Microbiol. 2015, 81, 5552–5559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argaw, A. Symbiotic effectiveness of inoculation with Bradyrhizobium isolates on soybean Glycine max (L.) Merrill genotypes with different maturities. SpringerPlus 2014, 3, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albareda, M.; Rodríguez-Navarro, D.N.; Temprano, F.J. Use of Sinorhizobium (Ensifer) fredii for soybean inoculants in South Spain. Eur. J. Agron. 2009, 30, 205–211. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, X.; Smith, D.L. Enhanced Soybean Plant Growth Resulting from Coinoculation of Bacillus Strains with Bradyrhizobium japonicum. Crop Sci. 2003, 43, 1774–1781. [Google Scholar] [CrossRef]
- Wichern, J.-M. Vergleich von Rhizobienpräparaten und Deren Applikation bei Sojabohnen in Nordrhein-Westfalen. Master’s Thesis, Rheinische Friedrich-Wilhelms-University of Bonn, Bonn, Germany, 2018. [Google Scholar]
- Grossman, J.M.; Schipanski, M.E.; Sooksanguan, T.; Seehaver, S.; Drinkwater, L.E. Diversity of rhizobia in soybean [Glycine max (Vinton)] nodules varies under organic and conventional management. Appl. Soil Ecol. 2011, 50, 14–20. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Andrade, D.d.S.; Chueire, L.M.d.O.; Takemura, S.M.; Hungria, M. Tillage method and crop rotation effects on the population sizes and diversity of bradyrhizobia nodulating soybean. Soil Biol. Biochem. 2000, 32, 627–637. [Google Scholar] [CrossRef]
- Hardarson, G.; Golbs, M.; Danso, S.K.A. Nitrogen fixation in soybean (Glycine max L. merrill) as affected by nodulation patterns. Soil Biol. Biochem. 1989, 21, 783–787. [Google Scholar] [CrossRef]
- Cassán, F.; Coniglio, A.; López, G.; Molina, R.; Nievas, S.; de Carlan, C.L.N.; Donadio, F.; Torres, D.; Rosas, S.; Pedrosa, F.O.; et al. Everything you must know about Azospirillum and its impact on agriculture and beyond. Biol. Fertil. Soils 2020, 56, 461–479. [Google Scholar] [CrossRef]
- Link, M.; Vorderbrügge, T.; Michalski, A.; Kowalkowski, A.; Harrach, T. Interpretation of German and Polish soil assessment data in order to deduce and to evaluate soil parameters and functions. Die Bodenkul. 2010, 61, 11–24. [Google Scholar]
- State Office for Environment, Agriculture and Geology, LfULG. Agrarmeteorologisches Messnetz Sachsen—Wetterdaten. Available online: https://www.landwirtschaft.sachsen.de/Wetter09/asp/inhalt.asp?seite=twerte (accessed on 24 June 2020).
- Deutscher Wetterdienst, DWD. CDC—Climate Data Center. Available online: https://cdc.dwd.de/portal/ (accessed on 24 June 2020).
- Association of German Agricultural Analytic and Research Institutes. Methode A 5.1.1: Bestimmung des pH-Wertes: 7. Teillfg. 2016. In Handbuch der Landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch); Association of German Agricultural Analytic and Research Institutes, Ed.; VDLUFA-Verlag: Darmstadt, Germany, 1991; ISBN 978-3-941273-30-5. [Google Scholar]
- Association of German Agricultural Analytic and Research Institutes. Methode A 6.2.1.1: Bestimmung von Phosphor und Kalium im Calcium-Acetat-Lactat-Auszug: 6. Teillfg. 2012. In Handbuch der Landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch); Association of German Agricultural Analytic and Research Institutes, Ed.; VDLUFA-Verlag: Darmstadt, Germany, 1991; ISBN 978-3-941273-30-5. [Google Scholar]
- Association of German Agricultural Analytic and Research Institutes. Methode A 6.2.4.1: Bestimmung des pflanzenverfügbaren Magnesiums im Calciumchlorid-Auszug. In Handbuch der Landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch); Association of German Agricultural Analytic and Research Institutes, Ed.; VDLUFA-Verlag: Darmstadt, Germany, 1991; ISBN 978-3-941273-30-5. [Google Scholar]
- DIN EN 15936:2012-11; Schlamm, Behandelter Bioabfall, Boden und Abfall—Bestimmung des Gesamten Organischen Kohlenstoffs (TOC) Mittels Trockener Verbrennung; Deutsche Fassung EN 15936:2012. Beuth Verlag GmbH: Berlin, Germay, 2012.
- Landwirtschaftliche Kommunikations- und Servicegesellschaft mbH, LKS. LMUAA 027 Bestimmung des Gesamtstickstoffs in Böden durch Verbrennung und Gasanalyse. In Anlage zur Akkreditierungsurkunde D-PL-14632-01-00 nach DIN EN ISO/IEC 17025:2018; DAkkS: Amberg, Germany, 2019. [Google Scholar]
- DIN ISO 11277:2002-08; Bodenbeschaffenheit—Bestimmung der Partikelgrößenverteilung in Mineralböden—Verfahren mittels Siebung und Sedimentation (ISO 11277:1998 + ISO 11277:1998 Corrigendum 1:2002). Beuth Verlag: Berlin, Germany, 2002. [CrossRef]
- Meier, U. Entwicklungsstadien Mono- und Dikotyler Pflanzen: BBCH Monografie; Blackwell: Berlin, Germany, 2018; ISBN 978-3-95547-070-8. [Google Scholar]
- Karasek, F.W.; Ray, E.C. Basic Gas Chromatography-Mass Spectrometry: Principles and Technique; Elsevier Science: Burlington, VT, USA; Wiley/Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 9780444427601. [Google Scholar]
- Munzert, M. Landwirtschaftliche und Gartenbauliche Versuche mit SAS; Springer: Berlin/Heidelberg, Germany, 2015; p. 449. ISBN 978-3-642-54505-4. [Google Scholar]
- Revellin, C.; Pinochet, X.; Beauclair, P.; Catroux, G. Influence of soil properties and soyabean cropping history on the Bradyrhizobium japonicum population in some French soils. Eur. J. Soil Sci. 1996, 47, 505–510. [Google Scholar] [CrossRef]
- Brunel, B.; Cleyet-Marel, J.C.; Normand, P.; Bardin, R. Stability of Bradyrhizobium japonicum Inoculants after Introduction into Soil. Appl. Environ. Microbiol. 1988, 54, 2636–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrzak, C.J.; Golinska, B.; Kroliczak, J.; Pudelko, K.; Lazewska, D.; Lampka, B.; Sadowsky, M.J. Diversity among Field Populations of Bradyrhizobium japonicum in Poland. Appl. Environ. Microbiol. 1995, 61, 1194–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, T.R.; Graham, P.H. Bradyrhizobium japonicum Inoculant Mobility, Nodule Occupancy, and Acetylene Reduction in the Soybean Root System. Appl. Environ. Microbiol. 1989, 55, 2493–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.C.; Hungria, M. Recovery of soybean inoculant strains from uncropped soils in Brazil. Field Crop. Res. 2002, 79, 139–152. [Google Scholar] [CrossRef]
- Parker, C.A.; Trinick, M.J.; Chatel, D.L. Rhizobia as soil and rhizosphere inhabitants. In A Treatise on Dinitrogen Fixation, Section III: Biology; John Wiley & Sons: New York, NY, USA, 1977; pp. 311–353. [Google Scholar]
- Zhang, Y.Z.; Wang, E.T.; Li, M.; Li, Q.Q.; Zhang, Y.M.; Zhao, S.J.; Jia, X.L.; Zhang, L.H.; Chen, W.F.; Chen, W.X. Effects of rhizobial inoculation, cropping systems and growth stages on endophytic bacterial community of soybean roots. Plant Soil 2011, 347, 147–161. [Google Scholar] [CrossRef]
- Weaver, R.W.; Frederick, L.R. Effect of Inoculum Rate on Competitive Nodulation of Glycine max L. Merrill. II. Field Studies. Agron. J. 1974, 66, 233–236. [Google Scholar] [CrossRef]
- Yan, J.; Han, X.Z.; Ji, Z.J.; Li, Y.; Wang, E.T.; Xie, Z.H.; Chen, W.F. Abundance and diversity of soybean-nodulating rhizobia in black soil are impacted by land use and crop management. Appl. Environ. Microbiol. 2014, 80, 5394–5402. [Google Scholar] [CrossRef] [Green Version]
- Weaver, R.W.; Frederick, L.R.; Dumenil, L.C. Effect of Soybean cropping and soil properties on numbers of Rhizobium japonicum in Iowa soils. Soil Sci. 1972, 114, 137–141. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Raizada, M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol. Biochem. 2017, 105, 177–196. [Google Scholar] [CrossRef]
- Vollmann, J.; Schweiger, P. Effekte der symbiontischen Stickstoff-Fixierung der Sojabohne. In 4. Umweltökologisches Symposium; Lehr- und Forschungszentrum für Landwirtschaft Raumberg-Gumpenstein: Irdning, Austria, 2014; pp. 105–107. [Google Scholar]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought Stress in Plants: An Overview. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.-C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef] [PubMed]
- Alam, F.; Bhuiyan, M.A.H.; Alam, S.S.; Waghmode, T.R.; Kim, P.J.; Lee, Y.B. Effect of Rhizobium sp. BARIRGm901 inoculation on nodulation, nitrogen fixation and yield of soybean (Glycine max) genotypes in gray terrace soil. Biosci. Biotechnol. Biochem. 2015, 79, 1660–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muneer, F.; Hovmalm, H.P.; Svensson, S.-E.; Newson, W.R.; Johansson, E.; Prade, T. Economic viability of protein concentrate production from greenbiomass of intermediate crops: A pre-feasibility study. J. Clean. Prod. 2021, 294, 126304. [Google Scholar] [CrossRef]
- Amann, C.; Amberger, A. Phosphorus Efficiency of Buckwheat (Fagopyrum esculentum). J. Plant Nutr. Soil Sci. 1989, 152, 181–189. [Google Scholar] [CrossRef]
- Bekere, W.; Hailemariam, A. Influences of Inoculation Methods and Phosphorus Levels on Nitrogen Fixation Attributes and Yield of Soybean (Glycine max L.) At Haru, Western Ethiopia. Am. J. Plant Nutr. Fertil. Technol. 2012, 2, 45–55. [Google Scholar] [CrossRef]
- Santachiara, G.; Salvagiotti, F.; Rotundo, J.L. Nutritional and environmental effects on biological nitrogen fixation in soybean: A meta-analysis. Field Crop. Res. 2019, 240, 106–115. [Google Scholar] [CrossRef]
- Zaeem, M.; Nadeem, M.; Pham, T.H.; Ashiq, W.; Ali, W.; Gilani, S.S.M.; Elavarthi, S.; Kavanagh, V.; Cheema, M.; Galagedara, L.; et al. The potential of corn-soybean intercropping to improve the soil health status and biomass production in cool climate boreal ecosystems. Sci. Rep. 2019, 9, 13148. [Google Scholar] [CrossRef] [Green Version]





| Date of Sowing | Date of Harvesting | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Site | Acronym Environment | Coordinates | m. a.s.l. (m) | Soil Texture | Soil Index | Mean Annual Temperature a (°C) | Annual Precipitation a (mm) | Inter-Crop | Main Crop | Inter-Crop | Main Crop | pH | P K Mg (mg·100 g−1 Soil) | Nt (%) | Humus (%) | ||
| 2016/2017 | Pillnitz | Pi 16_17 | 51°00′04.7″ N 13°53′23.3″ E | 118 | sL | 65 | 10.5 | 542 | 26 July 2016 | 1 May 2017 | 25 October 2016 | 12 October 2017 | 5.9 | 5.2 | 17.8 | 12.1 | 0.10 | 1.7 |
| Pawlowice | Pa 16_17 | 51°17′70.6″ N 17°11′60.2″ E | 240 | sL | 70 * | 9.8 | 557 | 24 August 2016 | 25 April 2017 | 31 October 2016 | 29 September 2017 | 5.8 | 8.0 | 13.6 | 11.5 | 0.10 | 1.6 | |
| 2017/2018 | Pillnitz | Pi 17_18 | 51°00′03.5″ N 13°53′26.2″ E | 118 | sL | 65 | 11.6 | 298 | 24 July 2017 | 2 May 2018 | 30 October 2017 | 29 August 2018 | 6.9 | 6.7 | 12.3 | 13.9 | 0.12 | 2,6 |
| Pawlowice | Pa 17_18 | 51°17′72.6″ N 17°11′40.2″ E | 240 | sL | 70 * | 10.5 | 399 | 16 August 2017 | 27 April 2018 | 29 October 2017 | 3 September 2018 | 6.4 | 8.2 | 14.5 | 8.2 | 0.10 | 1.7 | |
| 2018/2019 | Ebersbach | Eb 18_19 | 51°09′40.1″ N 14°55′37.2″ E | 208 | sL | 63 | 10.7 | 406 | 15 August 2018 | 25 April 2019 | 27 October 2018 | 12 September 2019 | 6.3 | 3.5 | 12.0 | 12.8 | 0.13 | 2.1 |
| Pawlowice | Pa 18_19 | 51°17′76.1″ N 17°10′79.6″ E | 240 | sL | 70 * | 10.8 | 448 | 7 August 2018 | 6 May 2019 | 31 October 2018 | 20 September 2019 | 6.3 | 5.8 | 16.0 | 9.0 | 0.10 | 1.7 | |
| Precipitation per Month (mm) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Environment | Precipitation Four Weeks before Sowing | July I | August I | September I | October I | November I | December I | January | February | March | April | May M | June M | July M | August M | September M | October M | ∑ |
| 16_17 | Pi | 91.4 | 93.2 | 43.1 | 51.5 | 71.5 | 29.7 | 26.8 | 10.6 | 22.0 | 41.0 | 39.9 | 26.4 | 56.3 | 92.0 | 82.8 | 32.2 | 69.4 | 788.4 |
| Pa | 33.2 | − | 27.1 | 44.7 | 83.8 | 36.3 | 36.1 | 16.9 | 24.2 | 31.1 | 57.0 | 24.1 | 52.5 | 112.2 | 43.6 | 65.7 | − | 655.3 | |
| 17_18 | Pi | 73.1 | 92.0 | 82.8 | 32.2 | 69.4 | 41.5 | 27.8 | 21.9 | 2.0 | 20.3 | 36.5 | 18.5 | 30.2 | 18.5 | 44.6 | − | − | 538.2 |
| Pa | 110.2 | − | 43.6 | 65.7 | 71.4 | 28.4 | 29.6 | 22.6 | 3.1 | 27.6 | 19.0 | 54.3 | 36.6 | 79.1 | 20.3 | 38.4 | − | 539.7 | |
| 18_19 | Eb | 22.6 | 36.9 | 16.0 | 52.2 | 35.7 | 7.7 | 67.4 | 40.6 | 24.0 | 47.6 | 14.5 | 74.2 | 31.4 | 42.3 | 53.2 | 22.4 | − | 566.1 |
| Pa | 16.5 | − | 20.3 | 38.4 | 45.3 | 14.7 | 38.0 | 56.2 | 27.7 | 22.5 | 24.2 | 76.8 | 27.0 | 44.5 | 59.8 | 42.0 | − | 537.4 | |
| Intercrop | Soybean Main Crop | |||||
|---|---|---|---|---|---|---|
| i | u | i | i | u | u | |
| Cultivated Crop | soybean (i) | soybean (u) | (ii) | (ui) ** | (iu) | (uu) |
| buckwheat (i) * | buckwheat (u) * | (ii) | (ui) ** | (iu) | (uu) | |
| soybean/buckwheat (i) | soybean/buckwheat (u) | (ii) | (ui) ** | (iu) | (uu) | |
| winter fallow (u) ** | (ui) ** | (uu) ** | ||||
| Intercrop | Inoculation | No. of Nodules per Plant | ±SE | Weight of Nodules per Plant (mg) | ±SE | Nodules_N (%) | ±SE | Nodules_S * (%) | ± SE | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pi 16_17 | BS | 2.6 | 0.4 | 16.99 | 3.53 | 5.45 | 0.19 | 0.26 | 0.006 | ||||
| S | 3.9 | 0.7 | 26.26 | 5.49 | 5.87 | 0.15 | 0.25 | 0.005 | |||||
| u/u | 0.2 | 0.0 | 1.12 | A | 0.36 | 5.49 | 0.34 | 0.26 | 0.009 | ||||
| i/u | 3.9 | 0.3 | 20.22 | B | 2.40 | 5.77 | 0.09 | 0.25 | 0.003 | ||||
| i/i | 5.6 | 0.4 | 43.54 | C | 4.60 | 5.72 | 0.13 | 0.25 | 0.005 | ||||
| Pi 17_18 | BS | 14.6 | 1.9 | 139.67 | 15.34 | 5.42 | 0.11 | 0.23 | 0.005 | ||||
| S | 14.9 | 2.2 | 150.94 | 16.48 | 5.31 | 0.13 | 0.23 | 0.007 | |||||
| u/u | 2.8 | A | 0.4 | 55.87 | 6.46 | 5.17 | 0.15 | 0.26 | 0.006 | ||||
| i/u | 19.6 | B | 0.9 | 189.9 | 7.77 | 5.28 | 0.09 | 0.21 | 0.004 | ||||
| i/i | 21.8 | B | 1.4 | 190.15 | 11.28 | 5.65 | 0.09 | 0.23 | 0.003 | ||||
| Eb 18_19 | BS | 0.3 | 0.04 | 0.56 | 0.11 | - | - | - | - | ||||
| S | 0.3 | 0.05 | 0.76 | 0.05 | - | - | - | - | |||||
| B | - | - | - | - | - | - | - | - | |||||
| u/u | 0.3 | 0.03 | 1.27 | 0.06 | - | - | - | - | |||||
| u/i | 0.2 | 0.01 | 0.13 | 0.02 | - | - | - | - | |||||
| i/u | 0.5 | 0.06 | 0.65 | 0.06 | - | - | - | - | |||||
| i/i | 0.3 | 0.05 | 0.58 | 0.15 | - | - | - | - | |||||
| Pa 17_18 | BS | 5.5 | B | 0.7 | 35.01 | B | 3.90 | 5.13 | B | 0.07 | 0.21 | 0.002 | |
| S | 3.5 | A | 0.5 | 12.82 | A | 1.70 | 4.81 | A | 0.04 | 0.22 | 0.002 | ||
| B | - | - | - | - | - | - | - | - | |||||
| u/u | 0.35 | A | 0.1 | 4.05 | A | 0.80 | 4.64 | A | 0.03 | 0.21 | 0.002 | ||
| i/u | 7.00 | B | 0.4 | 35.38 | B | 3.20 | 5.24 | B | 0.05 | 0.22 | 0.002 | ||
| i/i | 6.10 | B | 0.5 | 32.32 | B | 3.20 | 5.03 | B | 0.07 | 0.22 | 0.002 | ||
| Pa 18_19 | BS | 2.45 | A | 0.4 | 7.43 | 0.001 | 4.72 | 0.05 | 0.21 | 0.003 | |||
| S | 3.27 | B | 0.5 | 7.69 | 0.001 | 4.78 | 0.10 | 0.22 | 0.003 | ||||
| B | - | - | - | - | - | - | - | - | |||||
| u/u | 0.05 | A | 0.0 | 0.22 | 0.000 | 4.39 | A | 0.07 | 0.21 | 0.004 | |||
| i/u | 3.68 | B | 0.3 | 11.86 | 0.001 | 5.10 | C | 0.10 | 0.22 | 0.003 | |||
| i/i | 4.86 | B | 0.3 | 10.60 | 0.001 | 4.75 | B | 0.08 | 0.22 | 0.003 |
| Inter-Crop | Inoculation | No. of Nodules per Plant | ±SE | Weight of Nodules per Plant (mg) | ±SE | Nodules N [%] | ±SE | Nodules S * [%] | ±SE | No. of Nodules per Plant m.r. | ±SE | No. of Nodules per Plant l.r. | ±SE | Shoot_Ca [%] | ±SE | Shoot_P [%] | ±SE | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pi 16_17 | BS | 13.0 | 2.80 | 143.54 | 13.75 | - | - | - | - | - | - | - | - | 1.25 | 0.014 | 0.24 | 0.24 | ||||||
| S | 13.8 | 2.84 | 132.63 | 17.95 | - | - | - | - | - | - | - | - | 1.24 | 0.016 | 0.23 | 0.23 | |||||||
| u/u | 2.6 | A | 0.55 | 77.78 | A | 15.53 | - | - | - | - | - | - | - | - | 1.20 | 0.018 | 0.24 | 0.24 | |||||
| i/u | 7.8 | A | 0.53 | 126.09 | AB | 7.62 | - | - | - | - | - | - | - | - | 1.26 | 0.014 | 0.23 | 0.23 | |||||
| i/i | 29.9 | B | 2.12 | 210.38 | B | 8.93 | - | - | - | - | - | - | - | - | 1.22 | 0.012 | 0.22 | 0.22 | |||||
| Pi 17_18 | BS | 8.5 | 1.48 | 29.01 | 5.09 | 5.85 | 0.05 | 0.38 | 0.03 | - | - | - | - | 1.55 | 0.026 | 0.24 | 0.24 | ||||||
| S | 11.5 | 1.39 | 43.62 | 5.24 | 5.77 | 0.07 | 0.33 | 0.03 | - | - | - | - | 1.65 | 0.030 | 0.24 | 0.24 | |||||||
| u/u | 4.4 | A | 0.72 | 21.64 | A | 4.06 | 6.00 | 0.06 | 0.36 | 0.04 | - | - | - | - | 1.61 | 0.042 | 0.25 | B | 0.25 | ||||
| i/u | 7.8 | B | 1.26 | 34.38 | AB | 5.76 | 5.71 | 0.06 | 0.31 | 0.02 | - | - | - | - | 1.66 | 0.015 | 0.24 | B | 0.24 | ||||
| i/i | 17.8 | C | 0.56 | 52.92 | B | 4.18 | 5.73 | 0.05 | 0.40 | 0.03 | - | - | - | - | 1.67 | 0.027 | 0.23 | A | 0.23 | ||||
| Eb 18_19 | BS | 7.3 | 1.15 | 65.65 | 11.11 | - | - | - | - | 14.3 | 2.5 | 24.0 | 3.6 | 1.35 | B | 0.014 | 0.19 | 0.19 | |||||
| S | 6.6 | 1.05 | 65.35 | 10.44 | - | - | - | - | 13.4 | 2.0 | 24.8 | 3.7 | 1.27 | A | 0.010 | 0.18 | 0.18 | ||||||
| B | 7.7 | 1.30 | 70.31 | 11.38 | - | - | - | - | 17.1 | 3.2 | 21.4 | 4.1 | 1.27 | A | 0.015 | 0.18 | 0.18 | ||||||
| u/u | 0.1 | A | 0.02 | 0.14 | 0.05 | - | - | - | - | 0.0 | A | 0.0 | 0.2 | 0.1 | 1.33 | 0.018 | 0.19 | 0.19 | |||||
| u/i | 16.7 | C | 1.02 | 159.69 | 9.64 | - | - | - | - | 35.6 | C | 2.6 | 50.3 | 3.4 | 1.30 | 0.012 | 0.18 | 0.18 | |||||
| i/u | 0.2 | A | 0.05 | 1.03 | 0.38 | - | - | - | - | 0.0 | A | 0.0 | 0.6 | 0.2 | 1.25 | 0.010 | 0.19 | 0.19 | |||||
| i/i | 11.9 | B | 0.72 | 107.56 | 6.15 | - | - | - | - | 24.2 | B | 1.9 | 42.4 | 2.7 | 1.30 | 0.009 | 0.18 | 0.18 | |||||
| IMc u | 0.1 | A | 0.02 | 1.42 | 0.39 | - | - | - | - | 0.0 | A | 0.0 | 0.4 | 0.1 | 1.31 | 0.005 | 0.19 | 0.19 | |||||
| IMc i | 14.4 | B | 1.15 | 133.18 | 12.27 | - | - | - | - | 29.9 | B | 2.9 | 46.4 | 4.3 | 1.28 | 0.015 | 0.18 | 0.18 | |||||
| Pa 16_17 | BS | - | - | 41.87 | 5.69 | 6.41 | 0.05 | 0.31 | 0.01 | - | - | - | - | - | - | - | - | ||||||
| S | - | - | 54.05 | 7.89 | 6.49 | 0.04 | 0.30 | 0.00 | - | - | - | - | - | - | - | - | |||||||
| B | - | - | 53.03 | 6.62 | 0.13 | 0.00 | - | - | - | - | - | - | - | - | |||||||||
| u/u | - | - | 35.87 | 6.00 | 6.64 | 0.04 | 0.31 | 0.01 | - | - | - | - | - | - | - | - | |||||||
| i/u | - | - | 66.45 | 7.52 | 6.33 | 0.06 | 0.31 | 0.01 | - | - | - | - | - | - | - | - | |||||||
| i/i | - | - | 46.62 | 5.84 | 6.37 | 0.13 | 0.30 | 0.00 | - | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porte, A.; Lux, G.; Lewandowska, S.; Kozak, M.; Feller, J.; Schmidtke, K. Does a Soybean Intercrop Increase Nodule Number, N Uptake and Grain Yield of the Followed Main Crop Soybean? Agriculture 2022, 12, 467. https://doi.org/10.3390/agriculture12040467
Porte A, Lux G, Lewandowska S, Kozak M, Feller J, Schmidtke K. Does a Soybean Intercrop Increase Nodule Number, N Uptake and Grain Yield of the Followed Main Crop Soybean? Agriculture. 2022; 12(4):467. https://doi.org/10.3390/agriculture12040467
Chicago/Turabian StylePorte, Anne, Guido Lux, Sylwia Lewandowska, Marcin Kozak, Jörg Feller, and Knut Schmidtke. 2022. "Does a Soybean Intercrop Increase Nodule Number, N Uptake and Grain Yield of the Followed Main Crop Soybean?" Agriculture 12, no. 4: 467. https://doi.org/10.3390/agriculture12040467