Evaluation of the Antagonistic Effect of Pseudomonas Rhizobacteria on Fusarium Wilt of Chickpea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Foc Isolates
2.2. Isolation of Pseudomonas from the Rhizosphere of Chickpea
2.3. In Vitro Evaluation of Antagonistic Activities of Isolated Pseudomonas
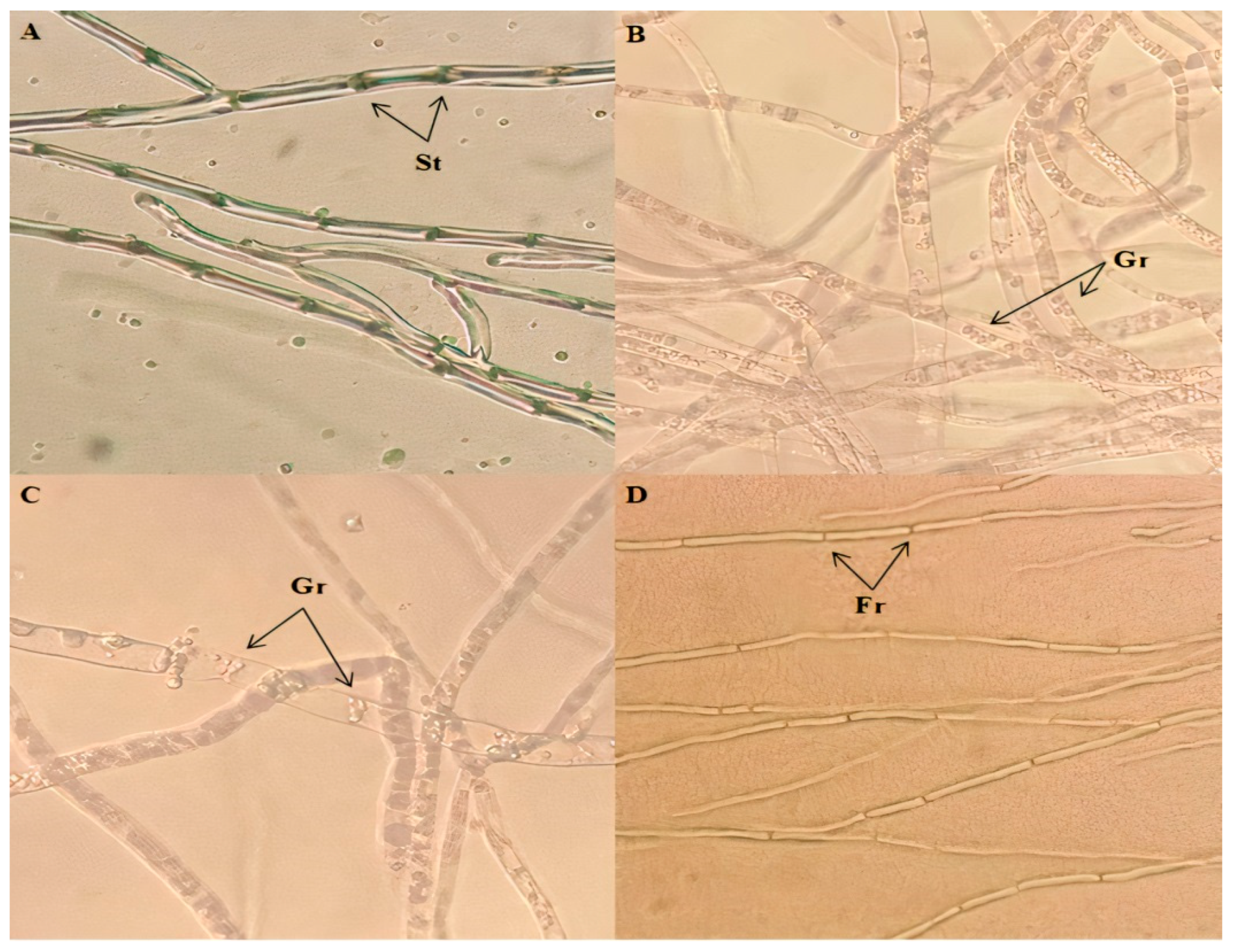
2.4. Effect of Pseudomonas Strains on Hyphae Morphology of Foc Isolates
2.5. Evaluation of PGPP
2.5.1. Protease Production
2.5.2. α-. Amylase Production
2.5.3. Lipase Production
2.5.4. Cellulase Production
2.5.5. Hydrogen Cyanide (HCN) Production
2.5.6. Indole Acetic Acid (IAA) Production
2.5.7. Phosphate Solubilization
2.5.8. Ammonia Production
2.5.9. Chitinase Production
2.5.10. Siderophores Production
2.6. Effect of Selected Pseudomonas Strains on the Shoot and Root Growth of Chickpea
2.7. Control of Wilt Disease of Chickpea by Selected Pseudomonas Strains
2.8. Molecular Identification of Selected Pseudomonas
2.9. Data Analysis
3. Results
3.1. In Vitro Evaluation of Antagonistic Activities of Isolated Pseudomonas
3.2. Effect of Pseudomonas Strains on Hyphae Morphology of Foc Isolates
3.3. Evaluation of PGPP
3.4. Effect of Selected Pseudomonas Strains on the Shoot and Root Growth of Chickpea
3.5. Control of Wilt Disease of Chickpea by Selected Pseudomonas Strains
3.6. Molecular Identification of Selected Pseudomonas
4. Discussion
5. Conclusions
6. Future Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawal, V.; Navarro, D.K. The Global Economy of Pulses; FAO: Rome, Italy, 2019. [Google Scholar] [CrossRef]
- Wood, J.A.; Grusak, M.A. Nutritional value of chickpea. In Chickpea Breeding and Management; Yadav, S.S., Redden, R.J., Chen, W., Eds.; CAB International: Wallingford, UK, 2007; pp. 101–142. [Google Scholar] [CrossRef]
- Latham, M.C. Human Nutrition in the Developing World; FAO: Rome, Italy, 1997. [Google Scholar]
- Aslam, M.; Mahmood, I.A.; Peoples, M.B.; Schwenke, G.D.; Herridge, D.F. Contribution of chickpea nitrogen fixation to increased wheat production and soil organic fertility in rain-fed cropping. Biol. Fertil. Soils 2003, 38, 59–64. [Google Scholar] [CrossRef]
- Landa, B.B.; Navas-Cortés, J.A.; del Mar Jimenez-Gasco, M.; Katan, J.; Retig, B.; Jiménez-Díaz, R.M. Temperature response of chickpea cultivars to races of Fusarium oxysporum f. sp. ciceris, causal agent of Fusarium wilt. Plant Dis. 2006, 90, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knights, E.J. Chickpea overview. In Encyclopedia of Grain Science; Wrigley, C., Corke, H., Walker, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2004; Volume 1, pp. 280–287. [Google Scholar] [CrossRef]
- Haware, M.P.; Jiménez-Diaz, R.M.; Amin, K.S.; Phillips, J.C.; Halila, H. Integrated management of wilt and root rots of chickpea. In Proceedings of the Chickpea in the Nineties, ICRISAT (International Crops Research Institute for the Semi-Arid Tropics), Patancheru, Andhra Pradesh, India, 4–8 December 1990; pp. 129–134. [Google Scholar]
- Gopalakrishnan, S.; Srinivas, V. Management of soil-borne diseases of grain legumes through broad-spectrum actinomycetes having plant growth-promoting and biocontrol traits. In Plant Microbe Interface; Varma, A., Tripathi, S., Prasad, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 129–144. [Google Scholar] [CrossRef]
- Landa, B.B.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M. Integrated management of Fusarium wilt of chickpea with sowing date, host resistance, and biological control. Phytopathology 2004, 94, 946–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arfaoui, A.; Sifi, B.; Boudabous, A.; El Hadrami, I.; Cherif, M. Identification of Rhizobium isolates possessing antagonistic activity against Fusarium oxysporum f. sp. ciceris, the causal agent of Fusarium wilt of chickpea. J. Plant Pathol. 2006, 88, 67–75. [Google Scholar]
- Jiménez-Gasco, M.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M. The Fusarium oxysporum f. sp. ciceris/Cicer arietinum pathosystem: A case study of the evolution of plant-pathogenic fungi into races and pathotypes. Int. Microbiol. 2004, 7, 95–104. [Google Scholar] [PubMed]
- Ramezani, H. Efficacy of some fungal and bacterial bioagents against Fusarium oxysporum f. sp. ciceri on chickpea. Plant Prot. J. 2009, 1, 108–113. [Google Scholar]
- Hayes, W.J.; Laws, E.R. Handbook of Pesticide Toxicology; Academic Press Inc.: New Delhi, India, 1991; Volume 1. [Google Scholar]
- Abed, H.; Rouag, N.; Mouatassem, D.; Rouabhi, A. Screening for Pseudomonas and Bacillus antagonistic rhizobacteria strains for the biocontrol of Fusarium wilt of chickpea. Eurasian J. Soil Sci. 2016, 5, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghafri, H.M.; Velazhahan, R.; Shahid, M.S.; Al-Sadi, A.M. Antagonistic activity of Pseudomonas aeruginosa from compost against Pythium aphanidermatum and Fusarium solani. Biocontrol. Sci. Technol. 2020, 30, 642–658. [Google Scholar] [CrossRef]
- Kumari, S.; Khanna, V. Biocidal mechanisms in biological control of Fusarium wilt in chickpea (Cicer arietinum L.) by antagonistic rhizobacteria: A current perspective in soil borne fungal pest management. Int. J. Curr. Microbiol. App. Sci. 2019, 8, 1494–1510. [Google Scholar] [CrossRef]
- Rathore, R.; Vakharia, D.N.; Rathore, D.S. In vitro screening of different Pseudomonas fluorescens isolates to study lytic enzyme production and growth inhibition during antagonism of Fusarium oxysporum f. sp. cumini, wilt causing pathogen of cumin. Egypt. J. Biol. Pest Control 2020, 30, 1–8. [Google Scholar] [CrossRef]
- Melo, I.S.; Souza, W.R.; Silva, L.J.; Santos, S.N.; Assalin, M.R.; Zucchi, T.D.; Queiroz, S.D.N. Antifungal activity of Pseudomonas frederiksbergensis CMAA 1323 isolated from the Antarctic hair grass Deschampsia antarctica. Br. Microbiol. Res. J. 2016, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium oxysporum Schlecht. Emend. Snyd. & Hans. In Fusarium Species: An Illustrated Manual for Identification; The Pennsylvania State University Press: Pennsylvania, PA, USA, 1983; pp. 142–145. [Google Scholar]
- Mishra, P.K.; Fox, R.T.; Culham, A. Development of a PCR-based assay for rapid and reliable identification of pathogenic Fusaria. FEMS Microbiol. Lett. 2003, 218, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Gasco, M.D.M.; Jiménez-Díaz, R.M. Development of a specific polymerase chain reaction-based assay for the identification of Fusarium oxysporum f. sp. ciceris and its pathogenic races 0, 1A, 5, and 6. Phytopathology 2003, 93, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, N.; Shimizu, M.; Hyakumachi, M. Occurrence of root rot and vascular wilt diseases in Roselle (Hibiscus sabdariffa L.) in Upper Egypt. Mycobiology 2014, 42, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowsalya, A.; Ponnusami, V.; Sugumaran, K.R. Isolation of bacteria from soil sample for exo-polysaccharide production. Int. J. Chemtech. Res. 2014, 6, 2925–2928. [Google Scholar]
- Cowan, S.T.; Steel, K.J. Diagnostic tables for the common medical bacteria. Epidemiol. Infect. 1961, 59, 357–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdogan, O.; Benlioglu, K. Biological control of Verticillium wilt on cotton by the use of fluorescent Pseudomonas spp. under field conditions. Biol. Control 2010, 53, 39–45. [Google Scholar] [CrossRef]
- Dilantha Fernando, W.G.; Linderman, R.G. Inhibition of Phytophthora vignae and stem and root rot of cowpea by soil bacteria. Biol. Agric. Hortic. 1995, 12, 1–14. [Google Scholar] [CrossRef]
- Etebarian, H.-R.; Sholberg, P.L.; Eastwell, K.C.; Sayler, R.J. Biological control of apple blue mold with Pseudomonas fluorescens. Can. J. Microbiol. 2005, 51, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Sheeba, J.; Dhamotharan, R.; Baskar, K. Isolation, screening and characterization of plant growth promoting bacteria and their antifungal effect on Rhizoctonia solani (JG Kühn 1858). Adv. Plants Agric. Res. 2017, 7, 369–375. [Google Scholar] [CrossRef]
- Zimbro, M.J.; Power, D.A.; Miller, S.M.; Wilson, G.E.; Johnson, J.A. (Eds.) Difco & BBL Manual: Manual of Microbiological Culture Media, 2nd ed.; Becton, Dickinson and Company: Maryland, MD, USA, 2009. [Google Scholar]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Khatiwada, P.; Ahmed, J.; Sohag, M.H.; Islam, K.; Azad, A.K. Isolation, screening and characterization of cellulase producing bacterial isolates from municipal solid wastes and rice straw wastes. J. Bioprocess Biotech. 2016, 6, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp.-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Arasu, V.S.; Kathiresan, K. Effect of Trichoderma on soil phosphate solubilization and growth improvement of Avicennia marina. Aquat. Bot. 2013, 104, 101–105. [Google Scholar] [CrossRef]
- Cappuccino, J.C.; Sherman, N. Microbiology: A Laboratory Manual, 3rd ed.; Benjamin Cummings Pub. Co.: New York, NY, USA, 1992. [Google Scholar]
- Murthy, N.; Bleakley, B. Simplified method of preparing colloidal chitin used for screening of chitinase-producing microorganisms. Internet J. Microbiol. 2012, 10, 1–5. [Google Scholar] [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.-P.; Xu, J.-G. A simple double-layered Chrome Azurol S Agar (SD-CASA) plate assay to optimize the production of siderophores by a potential biocontrol agent Bacillus. Afr. J. Microbiol. Res. 2011, 5, 4321–4327. [Google Scholar] [CrossRef]
- Kaur, R.; Singh, R.S.; Alabouvette, C. Antagonistic activity of selected isolates of fluorescent Pseudomonas against Fusarium oxysporum f. sp. ciceris. Asian J. Plant Sci. 2007, 6, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Zemouli-Benfreha, F.; Djamel-eddine, H.; Merzoug, A. Fusarium wilt of chickpea (Cicer arietinum L.) in North-West Algeria. Afr. J. Agric. Res. 2014, 9, 168–175. [Google Scholar] [CrossRef]
- Townsend, G.R. Methods for estimating losses caused by diseases in fungicide experiments. Plant Dis. Rep. 1943, 27, 340–343. [Google Scholar]
- Bektas, I.; Kusek, M. Biological control of onion basal rot disease using phosphate solubilising rhizobacteria. Biocontrol Sci. Technol. 2021, 31, 190–205. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitlo, S.A.; Rajput, N.A.; Syed, R.N.; Khanzada, M.A.; Rajput, A.Q.; Lodhi, A.M. Microbial control of Fusarium wilt of chickpea caused by Fusarium oxysporum f. sp. ciceris. Pak. J. Bot. 2019, 51, 2261–2268. [Google Scholar] [CrossRef]
- Trivedi, S.; Srivastava, M.; Ratan, V.; Mishra, A.; Dixit, S.; Pandey, S. Evaluation of microbial consortia on systemic resistance against chickpea wilt. Bangladesh J. Bot. 2020, 49, 653–661. [Google Scholar] [CrossRef]
- Ankati, S.; Srinivas, V.; Pratyusha, S.; Gopalakrishnan, S. Streptomyces consortia-mediated plant defense against Fusarium wilt and plant growth-promotion in chickpea. Microb. Pathog. 2021, 157, 104961. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Porter, J.K.; Norred, W.P.; Leslie, J.F. Production of fusaric acid by Fusarium species. Appl. Environ. Microbiol. 1996, 62, 4039–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Türkkan, M.; Dolar, F.S. Determination of fusaric acid production by Fusarium oxysporum f. sp. ciceris with thin layer chromatography and spectrophotometric methods. Anadolu Tarım Bilimleri Derg. 2010, 25, 146–150. [Google Scholar]
- Quecine, M.C.; Kidarsa, T.A.; Goebel, N.C.; Shaffer, B.T.; Henkels, M.D.; Zabriskie, T.M.; Loper, J.E. An interspecies signaling system mediated by fusaric acid has parallel effects on antifungal metabolite production by Pseudomonas protegens Strain Pf-5 and antibiosis of Fusarium spp. Appl. Environ. Microbiol. 2016, 82, 1372–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Khanna, V. Biological management of vascular wilt of chickpea (Cicer arietinum L.) incited by Fusarium oxysporum f. sp. ciceris by antagonistic rhizobacteria co-inoculated with native Mesorhizobium. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 920–941. [Google Scholar] [CrossRef]
- Vlassi, A.; Nesler, A.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile-mediated inhibitory activity of rhizobacteria as a result of multiple factors interaction: The case of Lysobacter capsici AZ78. Microorganisms 2020, 8, 1761. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Chinchilla, D.; Tan, C.; Mène-Saffrané, L.; L’Haridon, F.; Weisskopf, L. Contribution of hydrogen cyanide to the antagonistic activity of Pseudomonas strains against Phytophthora infestans. Microorganisms 2020, 8, 1144. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.S.; El-Sayyad, G.S.; Abd Elkodous, M.; El-Batal, A.I. The effective antagonistic potential of plant growth-promoting rhizobacteria against Alternaria solani-causing early blight disease in tomato plant. Sci. Hortic. 2020, 266, 109289. [Google Scholar] [CrossRef]
- Dharni, S.; Maurya, A.; Samad, A.; Srivastava, S.K.; Sharma, A.; Patra, D.D. Purification, characterization, and in vitro activity of 2, 4-di-tert-butylphenol from Pseudomonas monteilii PsF84: Conformational and molecular docking studies. J. Agric. Food Chem. 2014, 62, 6138–6146. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Gognies, S.; Nowak, J.; Audran, J.-C.; Belarbi, A. Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote the grapevine growth. Biol. Control 2002, 24, 135–142. [Google Scholar] [CrossRef]
- Giorgio, A.; De Stradis, A.; Lo Cantore, P.; Iacobellis, N.S. Biocide effects of volatile organic compounds produced by potential biocontrol rhizobacteria on Sclerotinia sclerotiorum. Front. Microbiol. 2015, 6, 1056. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, T.; Xu, M.; Guo, J.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Xing, K.; Qin, S. Antifungal effect of volatile organic compounds produced by Pseudomonas chlororaphis subsp. aureofaciens SPS-41 on oxidative stress and mitochondrial dysfunction of Ceratocystis fimbriata. Pestic. Biochem. Physiol. 2021, 173, 104777. [Google Scholar] [CrossRef]
- Verma, S.K.; Kingsley, K.L.; Bergen, M.S.; Kowalski, K.P.; White, J.F. Fungal disease prevention in seedlings of rice (Oryza sativa) and other grasses by growth-promoting seed-associated endophytic bacteria from invasive Phragmites australis. Microorganisms 2018, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara-Avendaño, E.; Bejarano-Bolívar, A.A.; Kiel-Martínez, A.-L.; Ramírez-Vázquez, M.; Méndez-Bravo, A.; von Wobeser, E.A.; Sánchez-Rangel, D.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Avocado rhizobacteria emit volatile organic compounds with antifungal activity against Fusarium solani, Fusarium Sp. associated with Kuroshio Shot Hole Borer, and Colletotrichum gloeosporioides. Microbiol. Res. 2019, 219, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Neilands, J.B. Methodology of siderophores. In Proceedings of the Siderophores from Microorganisms and Plants; Structure and Bonding; Springer: Berlin/Heidelberg, Germany, 1984; Volume 58, pp. 1–24. [Google Scholar] [CrossRef]
- Di Francesco, A.; Baraldi, E. How siderophore production can influence the biocontrol activity of Aureobasidium pullulans against Monilinia laxa on peaches. Biol. Control 2021, 152, 104456. [Google Scholar] [CrossRef]
- Nabila; Kasiamdari, R.S. Antagonistic activity of siderophore-producing bacteria from black rice rhizosphere against rice blast fungus Pyricularia oryzae. Microbiol. Biotechnol. Lett. 2021, 49, 217–224. [Google Scholar] [CrossRef]
- Zouari, I.; Masmoudi, F.; Medhioub, K.; Tounsi, S.; Trigui, M. Biocontrol and plant growth-promoting potentiality of bacteria isolated from compost extract. Antonie Leeuwenhoek 2020, 113, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Dewi, R.T.K.; Mubarik, N.R.; Suhartono, M.T. Medium optimization of β-glucanase production by Bacillus subtilis SAHA 32.6 used as biological control of oil palm pathogen. Emir. J. Food Agric. 2016, 28, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Gupta, M.; Gupta, S.; Kashyap, P.L.; Zargar, S.M.; Mallick, S.A. Antibiotic gene specific characterization and ARDRA analysis of native isolates of Pseudomonas spp. from Jammu, India. Indian Phytopathol. 2018, 71, 225–233. [Google Scholar] [CrossRef]
- Arora, N.K.; Khare, E.; Oh, J.H.; Kang, S.C.; Maheshwari, D.K. Diverse mechanisms adopted by fluorescent Pseudomonas PGC2 during the inhibition of Rhizoctonia solani and Phytophthora capsici. World J. Microbiol. Biotechnol. 2008, 24, 581–585. [Google Scholar] [CrossRef]
- Chiranjeevi, N.; Kumar, M.R.; Padmodaya, B.; Venkateswarlu, N.C.; Sudhakar, P.; Devi, R.S.J.; Sri, P.A. Studies on extraction, evaluation of crude metabolite extract from endophytic Bacillus subtillis and its mechanistic effect on chickpea dry root rot causing pathogen Rhizoctonia bataticola (Taub.) Butler. Pharma Innov. 2021, 10, 898–905. [Google Scholar]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Varma, P.K.; Kumar, V.K.; Suresh, M.; Chandrasekhar, V.; Bharathalakshmi, M.; Jamuna, P. Bacillus amyloliquefaciens (RB19): A potential PGPR in managing sugarcane red rot disease. J. Pharmacogn. Phytochem. 2019, 8, 2255–2261. [Google Scholar]
- Kalayu, G. Phosphate solubilizing microorganisms: Promising approach as biofertilizers. Int. J. Agron. 2019, 2019, 4917256. [Google Scholar] [CrossRef]
- Ul Hassan, T.; Bano, A. Construction of IAA-deficient mutants of Pseudomonas moraviensis and their comparative effects with wild type strains as bio-inoculant on wheat in saline sodic soil. Geomicrobiol. J. 2019, 36, 376–384. [Google Scholar] [CrossRef]
- Herlina, L.; Pukan, K.K.; Mustikaningtyas, D. The endophytic bacteria producing IAA (Indole Acetic Acid) in Arachis hypogaea. Cell Biol. Dev. 2017, 1, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Joshi, D.; Chandra, R.; Suyal, D.C.; Kumar, S. Impacts of bioinoculants Pseudomonas jesenii MP1 and Rhodococcus qingshengii S10107 on chickpea (Cicer arietinum L.) yield and soil nitrogen status. Pedosphere 2019, 29, 388–399. [Google Scholar] [CrossRef]
- Rawat, N.; Sharma, M.; Suyal, D.C.; Singh, D.K.; Joshi, D.; Singh, P.; Goel, R. Psyhcrotolerant bio-inoculants and their co-inoculation to improve Cicer arietinum growth and soil nutrient status for sustainable mountain agriculture. J. Soil Sci. Plant Nutr. 2019, 19, 639–647. [Google Scholar] [CrossRef]
- Gaurav, A.; Pallavi, K.P. Effect of plant growth-promoting rhizobacteria (PGPR) on growth and physiological parameters in chickpea. J. Pharmacogn. Phytochem. 2020, 9, 29–34. [Google Scholar]
- Besset-Manzoni, Y.; Joly, P.; Brutel, A.; Gerin, F.; Soudière, O.; Langin, T.; Prigent-Combaret, C. Does in vitro selection of biocontrol agents guarantee success in planta? A study case of wheat protection against Fusarium seedling blight by soil bacteria. PLoS ONE 2019, 14, e0225655. [Google Scholar] [CrossRef] [Green Version]
- Hussaini, S.Z.; Shaker, M.; Iqbal, M.A. Isolation of bacterial for degradation of selected pesticides. Adv Biores 2013, 4, 82–85. [Google Scholar]
- Chatterjee, P.; Samaddar, S.; Anandham, R.; Kang, Y.; Kim, K.; Selvakumar, G.; Sa, T. Beneficial soil bacterium Pseudomonas Frederiksbergensis OS261 augments salt tolerance and promotes red pepper plant growth. Front. Plant Sci. 2017, 8, 705. [Google Scholar] [CrossRef]
- Ferchichi, N.; Toukabri, W.; Boularess, M.; Smaoui, A.; Mhamdi, R.; Trabelsi, D. Isolation, identification and plant growth promotion ability of endophytic bacteria associated with lupine root nodule grown in Tunisian soil. Arch. Microbiol. 2019, 201, 1333–1349. [Google Scholar] [CrossRef]
- Hong, C.E.; Kim, J.U.; Lee, J.W.; Lee, S.W.; Jo, I.-H. Diversity of bacterial endophytes in Panax ginseng and their protective effects against pathogens. 3 Biotech 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Ben Zineb, A.; Trabelsi, D.; Ayachi, I.; Barhoumi, F.; Aroca, R.; Mhamdi, R. Inoculation with elite strains of phosphate-solubilizing bacteria enhances the effectiveness of fertilization with rock phosphates. Geomicrobiol. J. 2020, 37, 22–30. [Google Scholar] [CrossRef]
- Riera, N.; Handique, U.; Zhang, Y.; Dewdney, M.M.; Wang, N. Characterization of antimicrobial-producing beneficial bacteria isolated from Huanglongbing escape citrus trees. Front. Microbiol. 2017, 8, 2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swift, R.G. Plant Growth-Promoting Bacteria from Western Australian Soils. Ph.D. Thesis, Murdoch University, Perth, Australia, 2016. [Google Scholar]



{kind=link}
{kind=link}
| Isolates | Foc-S1 | Foc-S2 | ||
|---|---|---|---|---|
| Assays | Co-Culture | Divided Plate | Co-Culture | Divided Plate |
| E2PP3 | 4.66 ± 2.48 a | 44.70 ± 4.09 b | 17.13 ± 4.32 abc | 23.62 ± 3.25 bcd |
| E1PP14 | 11.81 ± 5.59 abc | 44.51 ± 2.14 b | 14.96 ± 4.38 ab | 12.57 ± 7.62 ab |
| E1FP14 | 9.36 ± 3.64 ab | 24.89 ± 4.09 a | 22.98 ± 2.78 abcd | 32.69 ± 5.85 cde |
| E2PP4 | 17.43 ± 1.06 bcd | 43.95 ± 8.90 b | 17.81 ± 0.24 abc | 22.69 ± 5.80 abcd |
| E1PP15 | 21.85 ± 3.27 cdef | 47.43 ± 8.58 b | 14.69 ± 1.23 ab | 45.38 ± 4.68 e |
| E2PP7 | 24.10 ± 3.90 def | 44.22 ± 2.29 b | 13.24 ± 6.67 a | 19.37 ± 6.75 abc |
| E2PP5 | 20.49 ± 1.29 bcde | 44.61 ± 0.09 b | 17.18 ± 8.82 abc | 19.17 ± 7.68 abc |
| E1FP8 | 22.16 ± 2.02 cdef | 45.01 ± 0.41 b | 22.75 ± 5.54 abcd | 14.25 ± 4.86 ab |
| E2PP8 | 17.08 ± 3.38 bcd | 43.51 ± 0.37 b | 28.72 ± 5.28 cdef | 18.56 ± 8.86 abc |
| E1FP9 | 24.59 ± 2.97 def | 44.89 ± 0.51 b | 23.97 ± 1.99 abcde | 13.09 ± 2.64 ab |
| E1PP2 | 22.95 ± 3.31 cdef | 43.77 ± 2.47 b | 27.36 ± 0.19 bcde | 20.49 ± 6.92 abc |
| E2PP6 | 30.10 ± 8.48 ef | 45.00 ± 1.34 b | 21.12 ± 5.74 abcd | 12.26 ± 4.34 ab |
| E1PP7 | 25.61 ± 2.81 def | 47.97 ± 5.04 b | 32.33 ± 4.67 def | 35.55 ± 3.37 de |
| E1FP4 | 23.24 ± 1.41 cdef | 48.37 ± 6.31 b | 40.52 ± 4.43 f | 6.88 ± 0.24 a |
| E2PP2 | 33.03 ± 8.54f | 44.19 ± 2.26 b | 35.70 ± 3.68 ef | 19.88 ± 4.57 abc |
| E1PP6 | 19.06 ± 0.73 bcde | 43.44 ± 6.67 b | 52.20 ± 4.51 g | 7.02 ± 2.18 a |
| E1FP13 | 19.24 ± 1.00 bcde | 52.77 ± 2.32 b | 52.09 ± 2.65 g | 15.06 ± 6.13 ab |
| Mean | 20.40 | 44.31 | 26.75 | 19.91 |
| Tests | E1PP7 | E1FP4 | E1PP15 | E1PP6 | E1FP13 |
|---|---|---|---|---|---|
| α-amylase production | - | - | - | - | - |
| Lipase production | ++++ | + | +++ | - | + |
| Protease production | + | - | ++ | - | - |
| Cellulase production | + | + | + | ++ | + |
| Chitinase production | - | - | - | - | - |
| HCN production | ++ | - | + | ++ | - |
| IAA production | + | + | ++ | + | ++ |
| Ammonia production | +++ | + | ++ | + | + |
| Siderophores production | ++ | +++ | ++ | +++ | +++ |
| Qualitative phosphate solubilization | + | + | + | ++ | ++ |
| Quantitative phosphate solubilization (µg mL−1) | 223.09 ± 18.99 a | 240.69 ± 0.06 a | 173.80 ± 1.47 b | 62.82 ± 6.01 c | 57.66 ± 11.40 c |
| Selected Pseudomonas | Shoot Length (cm) | Root Length (cm) | Shoot Dry Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|
| E1PP6 | 28.80 ± 0.69 b | 12.13 ± 1.33 a | 0.23 ± 0.02 b | 0.14 ± 0.04 ab |
| E1FP13 | 30.17 ± 1.46 b | 17.97 ± 0.96 bc | 0.26 ± 0.01 b | 0.20 ± 0.01 b |
| E1FP4 | 30.23 ± 1.66 b | 14.83 ± 1.56 ab | 0.27 ± 0.02 b | 0.18 ± 0.02 b |
| E1PP7 | 35.77 ± 1.10 c | 20.53 ± 0.38 c | 0.32 ± 0.04 c | 0.20 ± 0.04 b |
| E1PP15 | 23.57 ± 0.57 a | 14.53 ± 1.19 ab | 0.18 ± 0.02 a | 0.17 ± 0.01 b |
| Control | 25.07 ± 1.87 a | 11.17 ± 2.40 a | 0.17 ± 0.02 a | 0.11 ± 0.01 a |
| Strains | Foc-S1 | Strains | Foc-S2 | ||
|---|---|---|---|---|---|
| DS% | DR% | DS% | DR% | ||
| E1FP13 | 31.94 ± 6.36 a | 55.77 ± 8.81 a | E1FP13 | 38.89 ± 4.82 a | 53.33 ± 5.78 b |
| E1FP4 | 37.50 ± 4.17 a | 48.08 ± 5.78 a | E1PP6 | 44.44 ± 9.62 ab | 46.67 ± 11.55 ab |
| E1PP7 | 38.89 ± 9.62 a | 46.16 ± 13.33 a | E1PP15 | 58.33 ± 0.00 b | 30.00 ± 0.00 a |
| Control | 72.22 ± 12.73 b | 00.00 | Control | 83.33 ± 8.34 c | 00.00 |
| Isolates | Accession | Best Match | PI * | E Value |
|---|---|---|---|---|
| E1PP6 | MT774541 | Pseudomonas frederiksbergensis | 99.55% | 0.0 |
| E1FP13 | MT774542 | Pseudomonas frederiksbergensis strain LMR 708 | 99.56% | 0.0 |
| E1PP7 | MT774544 | Pseudomonas granadensis strain F-278,770T | 100% | 0.0 |
| E1PP15 | MT774545 | Pseudomonas granadensis strain F-278,770T | 99.77% | 0.0 |
| E1FP4 | MT774548 | Pseudomonas frederiksbergensis strain LMR708 | 99.89% | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalifa, M.W.; Rouag, N.; Bouhadida, M. Evaluation of the Antagonistic Effect of Pseudomonas Rhizobacteria on Fusarium Wilt of Chickpea. Agriculture 2022, 12, 429. https://doi.org/10.3390/agriculture12030429
Khalifa MW, Rouag N, Bouhadida M. Evaluation of the Antagonistic Effect of Pseudomonas Rhizobacteria on Fusarium Wilt of Chickpea. Agriculture. 2022; 12(3):429. https://doi.org/10.3390/agriculture12030429
Chicago/Turabian StyleKhalifa, Meriem Wafaa, Noureddine Rouag, and Mariem Bouhadida. 2022. "Evaluation of the Antagonistic Effect of Pseudomonas Rhizobacteria on Fusarium Wilt of Chickpea" Agriculture 12, no. 3: 429. https://doi.org/10.3390/agriculture12030429