Differential Physiological Response and Antioxidant Activity Relative to High-Power Micro-Waves Irradiation and Temperature of Tomato Sprouts

 ,
,
Abstract
:1. Introduction
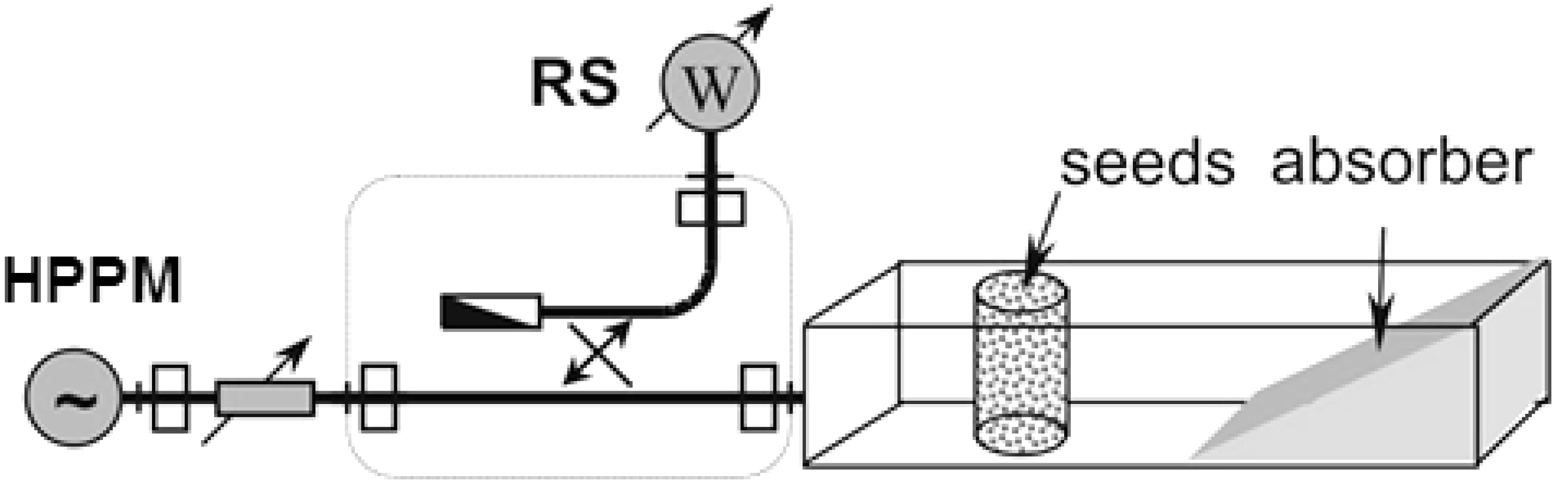
2. Materials and Methods
3. Results and Discussion
4. Conclusions
- HPM (9.3 GHz) and elevated temperature exposure upon tomato seeds and sprouts in their primary ontogenetic development level showed a slightly incentive effect on plants growing parameters: dry mass, fresh mass, plants height, and assimilation area.
- Elevated temperature (30 °C) increased the amount of photosynthetic pigments (chlorophyll a, chlorophyll b, and carotenoids) in tomato sprout leaves.
- Tomato plants (+R) and seeds (R) irradiation significantly reduced the content of non-structural carbohydrates (raphinose, sucrose, mannose, fructose, and glucose) at the temperature of 21 °C.
- Seed irradiation by HPM and the elevated temperature caused significantly lower scavenging of DPPH free-radical activity in tomato sprouts.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glazier, E.V.D.; Lamont, H.R.L. Services Textbook of Radio, 5th ed.; Her Majesty’s Stationary Office: Goring-by-Sea, UK, 1958; pp. 190–450. [Google Scholar]
- Radzevičius, A.; Viškelis, J.; Karklelienė, R.; Juškevičienė, D.; Viškelis, P. Determination of tomato quality attributes using near infrared spectroscopy and reference analysis. Zemdirb. Agric. 2016, 103, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Abbey, L.; Udenigwe, C.; Mohan, A.; Anom, E. Microwave irradiation effects on vermicasts potency, and plant growth and antioxidant activity in seedlings of Chinese cabbage (Brassica rapa subsp. pekinensis). J. Radiat. Res. Appl. Sci. 2017, 10, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, R.; Bianchi, G. Microwave and RF Engineering; John Wiley & Sons: Chichester, UK, 2010; pp. 25–107. [Google Scholar]
- Chiang, H.; Yao, G.D.; Fang, Q.S.; Wang, K.Q.; Lu, D.Z.; Zhou, Y.K. Health effects of environmental electromagnetic fields. J. Bioelectric 1989, 8, 127–131. [Google Scholar] [CrossRef]
- Roy, I.; Gupta, M.N. Applications of microwaves in biological sciences. Curr. Sci. 2003, 12, 1685–1693. [Google Scholar]
- Hamada, E. Effects of microwave treatment on growth, photosynthetic pigments and some metabolites of wheat. Biol. Plant. 2007, 2, 343–345. [Google Scholar] [CrossRef]
- Halmagyi, A.; Surducan, E.; Surducan, V. The effect of low- and high-power microwave irradiation on in vitro grown Sequoia plants and their recovery after cryostorage. J. Biol. Phys. 2017, 43, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Sharma, V.; Kumari, N. Microwave pretreatment of tomato seeds and fruit to enhance plant photosynthesis, nutritive quality and shelf life of fruit. Postharvest Biol. Technol. 2020, 159, 111015. [Google Scholar] [CrossRef]
- Ursache, M.; Mindru, G.; Creanga, D.E.; Tufescu, F.M.; Goiceanu, C. The effects of high frequency electromagnetic waves on the vegetal organisms. Rom. J. Phys. 2007, 54, 133–145. [Google Scholar]
- Zhang, J.; Zhanga, D.; Fan, Y.; He, J.; Ge, X.; Zhang, X.; Ju, J.; Xun, T. Progress in narrowband high-power microwave sources. Phys. Plasmas 2020, 27, 010501. [Google Scholar] [CrossRef]
- Radzevičius, A.; Sakalauskienė, S.; Dagys, M.; Simniškis, R.; Karklelienė, R.; Bobinas, Č.; Duchovskis, P. The effect of strong microwave electric field radiation on: (1) vegetable seed germination and seedling growth rate. Zemdirb. Agric. 2013, 100, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Stašelis, A. Influence of direct electromagnetic fields on germination dynamics of carrot seeds, carrot yield and quality. Zemdirb. Agric. 2007, 4, 201–207. (In Lithuanian) [Google Scholar]
- Yanenko, A.F.; Matsibura, A.P.; Peregudov, S.N.; Uniyaka, T.L. Impact of microwave radiation on vegetable biological objects. In Proceedings of the 14th international Crimean conference Microwave and Telecommunication Technology, Sevastopol, Ukraine, 13–17 September 2004; IEEE: Piscataway, NJ, USA, 2004. [Google Scholar]
- Aladjadjiyan, A. Effect of microwave irradiation on seeds of lentils (Lens culinaris, med.). Rom. J. Biophys. 2010, 3, 213–221. [Google Scholar]
- Rifna, E.J.; Ratish, R.K.; Mahendran, R. Emerging technology applications for improving seed germination. Trends Food Sci. Technol. 2019, 86, 95–108. [Google Scholar] [CrossRef]
- Chen, H.H.; Shen, Z.Y.; Li, P.H. Adaptability of crop plants to high-temperature stress. Crop Sci. 1982, 22, 719–725. [Google Scholar] [CrossRef]
- Gavrilenko, V.F.; Zigalova, T.V.A. Great Practical on Photosynthesis; Academia: Moscow, Russia, 2003; pp. 100–242. (In Lithuanian) [Google Scholar]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.O.; Bhat, K.T. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Janghel, E.K.; Gupta, V.K.; Rai, M.K.; Rai, J.K. Micro determination of ascorbic acid using methyl viologen. Talanta 2007, 72, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Heuvelink, E. Dry Matter Production in a Tomato Crop: Measurements and Simulation. Ann. Bot. 1995, 75, 369–379. [Google Scholar] [CrossRef]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A.; Gilroy, S. Glutamate triggers long-distance, calcium-based plant defence signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjarvi, J.; Mittler, R. Rapid responses to abiotic stress: Priming the landscape for the signal transduction network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Brainina, K.; Tarasov, A.; Khamzina, E.; Stozhko, N.; Vidrevich, M. Contact hybrid potentiometric method for on-site and in situ estimation of the antioxidant activity of fruits and vegetables. Food Chem. 2020, 309, 125703. [Google Scholar] [CrossRef]
- Ponomarev, L.L.; Dolgodvorov, V.E.; Popov, V.V.; Rodin, S.V.; Roman, O.A. Effect of low intensity electromagnetic microwave field on the germination and power of cereal seeds. Proc. Timiryazev Agric. Acad. 1996, 2, 42–46. [Google Scholar]
- Dukic, V.; Miladinov, Z.; Dozet, G.; Cvijanovic, M.; Tatis, M.; Miladinovic, J.; Balesevic-Tubic, S. Pulsed electromagnetic field—A cultivation practice used to increase soybean seed germination and yield. Zemdirb. Agric. 2017, 104, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Dario, A.C.; Salgado, J.M. Supplementation of irradiated and non-irradiated cowpea bean (Vigna unguiculata L. Walp) protein with cereal proteins. Plants Foods Hum. Nutr. 1994, 46, 213–219. [Google Scholar] [CrossRef]
- Racuciu, M.; Miclaus, S. Low-level 900 MHz electromagnetic field influence on vegetal tissue. Rom. J. Biophys. 2007, 3, 149–156. [Google Scholar]
- Salama, H.M.H.; Watban, A.A.A.; Al-Fughom, A.T. Effect of ultraviolet radiation on chlorophyll, carotenoid, protein and proline contents of some annual desert plants. Saudi J. Biol. Sci. 2011, 18, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Vian, A.; Davies, E.; Gendraud, M.; Bonnet, P. Plant Responses to High Frequency Electromagnetic Field. BioMed Res. Int. 2016, 2016, 1830262. [Google Scholar] [CrossRef] [Green Version]
- Karklelienė, R.; Juškevičienė, D.; Radzevičius, A.; Sasnauskas, A. Productivity and adaptability of the new carrot and garlic cultivars in Lithuania. Zemdirb. Agric. 2018, 105, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, S.A.; Abdelrahim, H.A.H.; Mohamed, A.G.K.; Samy, S.; Soad, K.A.J.; Hamada, E. Laser light as a promising approach to improve the nutritional value, antioxidant capacity and anti-inflammatory activity of flavonoid-rich buckwheat sprouts. Food Chem. 2021, 345, 128788. [Google Scholar]
- Ortiz, J.; Suarez, D.; Puentes, A.; Velasquez, P.; Santis, N.A. Comparison of the effects in the germination and growth of corn seeds (Zea mays L.) by exposure to magnetic, electrical and electromagnetic fields. Chem. Eng. Trans. 2015, 43, 169–174. [Google Scholar]
- Farid, M.; Ali, S.; Rizwan, M.; Saeed, R.; Tauqeer, H.M.; Sallah-Ud-Din, M.; Azam, A.; Raza, N. Microwave irradiation and citric acid assisted seed germination and phytoextraction of nickel (Ni) by Brassica napus L.: Morpho-physiological and biochemical alterations under Ni stress. Environ. Sci. Pollut. Res. 2017, 24, 21050–21064. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Moore, B.; Sheen, J. Sugar sensing and signaling in plants. Plant Cell 2002, 14, 185–205. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 3, 869–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramel, F.; Sulmon, C.; Gousbest, G.; Couee, I. Natural variation reveals relationship between pre-stress carbohydrate nutritional status and subsequent responses to xenobiotic and oxiditave stress in Arabidobsis thaliana. Ann. Bot. 2009, 104, 1323–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.A.K.; Hectors, K.; O’Brien, N.M.; Guisez, Y.; Potters, G. Plant stress and human health: Do human consumers benefit from UV-B acclimated crops? Plant Sci. 2008, 175, 449–458. [Google Scholar] [CrossRef]
- Sakalauskaitė, J.; Viškelis, P.; Dambrauskienė, E.; Sakalauskienė, S.; Samuolienė, G.; Brazaitytė, A.; Duchovskis, P.; Urbonavičienė, D. The effects of different UV-B radiation intensities on morphological and biochemical characteristics in Ocimum basilicum L. J. Sci. Food Agric. 2013, 93, 1266–1271. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Arbogast, J.; Mittler, R. Coordinated and rapid whole-plant systemic stomatal responses. New Phytol. 2019, 225, 21–25. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Impact | Height, cm | STDEV | Dry Weight, g | STDEV | Fresh Weight, g | STDEV | Assimilation Area, cm2 | STDEV |
|---|---|---|---|---|---|---|---|---|
| T—21 °C | ||||||||
| Control | 12.20 | ±0.770 | 0.30 | ±0.017 | 2.81 | ±0.147 | 85.33 | ±5.691 |
| R | 13.30 | ±0.361 | 0.33 | ±0.011 | 2.93 | ±0.078 | 95.33 | ±1.543 |
| +R | 13.47 | ±0.424 | 0.30 | ±0.014 | 3.13 | ±0.094 | 91.70 | ±1.764 |
| T—30 °C | ||||||||
| Control | 18.20 | ±0.529 | 0.27 | ±0.026 | 3.12 | ±0.116 | 74.66 | ±15.823 |
| R | 19.50 | ±0.416 | 0.29 | ±0.019 | 3.52 | ±0.241 | 105.67 | ±8.471 |
| +R | 19.20 | ±0.395 | 0.26 | ±0.030 | 3.20 | ±0.184 | 96.33 | ±5.275 |
| Impact | Carotenoidsmg g−1, FW | STDEV | Chlorophyll a mg g−1, FW | STDEV | Chlorophyll b mg g−1, FW | STDEV | Chlorophylls a + b mg g−1, FW | STDEV |
|---|---|---|---|---|---|---|---|---|
| T—21 °C | ||||||||
| Control | 0.33 | 0.050 | 1.15 | 0.146 | 0.45 | 0.026 | 1.60 | 0.098 |
| R | 0.35 | 0.012 | 1.21 | 0.091 | 0.45 | 0.020 | 1.66 | 0.080 |
| +R | 0.28 | 0.026 | 1.00 | 0.072 | 0.47 | 0.029 | 1.47 | 0.084 |
| T—30 °C | ||||||||
| Control | 0.41 | 0.030 | 1.51 | 0.121 | 0.68 | 0.067 | 2.19 | 0.203 |
| R | 0.44 | 0.023 | 1.50 | 0.059 | 0.59 | 0.027 | 2.09 | 0.083 |
| +R | 0.46 | 0.019 | 1.43 | 0.083 | 0.69 | 0.059 | 2.11 | 0.061 |
| Impact | Raphinose, mg g−1, FW | STDEV | Sucrose, mg g−1, FW | STDEV | Glucose, mg g−1, FW | STDEV | Manosse, mg g−1, FW | STDEV | Fructose, mg g−1, FW | STDEV |
|---|---|---|---|---|---|---|---|---|---|---|
| T—21 °C | ||||||||||
| Control | 2.02 | 0.261 | 2.53 | 0.315 | 1.69 | 0,475 | 1.98 | 0.510 | 0.66 | 0.028 |
| R | 1.43 | 0.218 | 2.33 | 0.397 | 0.64 | 0.279 | 0.77 | 0.461 | 0.01 | 0.019 |
| +R | 0.87 | 0.447 | 1.08 | 0.283 | 0.47 | 0.237 | 0.28 | 0.297 | 0.23 | 0.370 |
| T—30 °C | ||||||||||
| Control | 1.48 | 0.139 | 1.23 | 0.147 | 0.68 | 0.230 | 0.00 | 0.000 | 1.21 | 0.405 |
| R | 1.02 | 0.217 | 0.15 | 0.083 | 0.04 | 0.015 | 0.37 | 0.271 | 0.46 | 0.291 |
| +R | 0.74 | 0.294 | 1.20 | 0.196 | 0.95 | 0.367 | 1.26 | 1.043 | 0.26 | 0.195 |
| Impact | Total Phenols, mg g−1, FW | STDEV | DPPH, µmol g−1, FW | STDEV | Ascorbic Acid, mg g−1, FW | STDEV |
|---|---|---|---|---|---|---|
| T—21 °C | ||||||
| Control | 0.69 | 0.092 | 10.64 | 0.107 | 0.27 | 0.048 |
| R | 0.92 | 0.103 | 10.29 | 0.196 | 1.32 | 0.719 |
| +R | 0.60 | 0.118 | 9.09 | 1.009 | 0.48 | 0.103 |
| T—30 °C | ||||||
| Control | 0.59 | 0.173 | 9.00 | 0.208 | 0.23 | 0.065 |
| R | 0.57 | 0.167 | 7.03 | 1.517 | 0.33 | 0.027 |
| +R | 0.49 | 0.139 | 8.51 | 0.231 | 0.37 | 0.064 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzevičius, A.; Sakalauskienė, S.; Dagys, M.; Simniškis, R.; Karklelienė, R.; Juškevičienė, D.; Račkienė, R.; Brazaitytė, A. Differential Physiological Response and Antioxidant Activity Relative to High-Power Micro-Waves Irradiation and Temperature of Tomato Sprouts. Agriculture 2022, 12, 422. https://doi.org/10.3390/agriculture12030422
Radzevičius A, Sakalauskienė S, Dagys M, Simniškis R, Karklelienė R, Juškevičienė D, Račkienė R, Brazaitytė A. Differential Physiological Response and Antioxidant Activity Relative to High-Power Micro-Waves Irradiation and Temperature of Tomato Sprouts. Agriculture. 2022; 12(3):422. https://doi.org/10.3390/agriculture12030422
Chicago/Turabian StyleRadzevičius, Audrius, Sandra Sakalauskienė, Mindaugas Dagys, Rimantas Simniškis, Rasa Karklelienė, Danguolė Juškevičienė, Roma Račkienė, and Aušra Brazaitytė. 2022. "Differential Physiological Response and Antioxidant Activity Relative to High-Power Micro-Waves Irradiation and Temperature of Tomato Sprouts" Agriculture 12, no. 3: 422. https://doi.org/10.3390/agriculture12030422