
Transpiration Responses of Herbicide-Resistant and -Susceptible Palmer Amaranth (Amaranthus palmeri (S.) Wats.) to Progressively Drying Soil

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Comparison of Transpiration Rates between Glyphosate-Resistant and -Susceptible Plants That Differ in EPSPS Copy Number
Plant Material Preparation
2.2. Differences in Drought Tolerance between S-Metolachlor-Susceptible and -Resistant Accessions
Plant Material Preparation
2.3. Dry-Down Treatment
2.3.1. Daily Transpiration and Normalized Transpiration Ratio
2.3.2. Drought Stress Level in the Root Zone
2.4. Data Analysis
3. Results
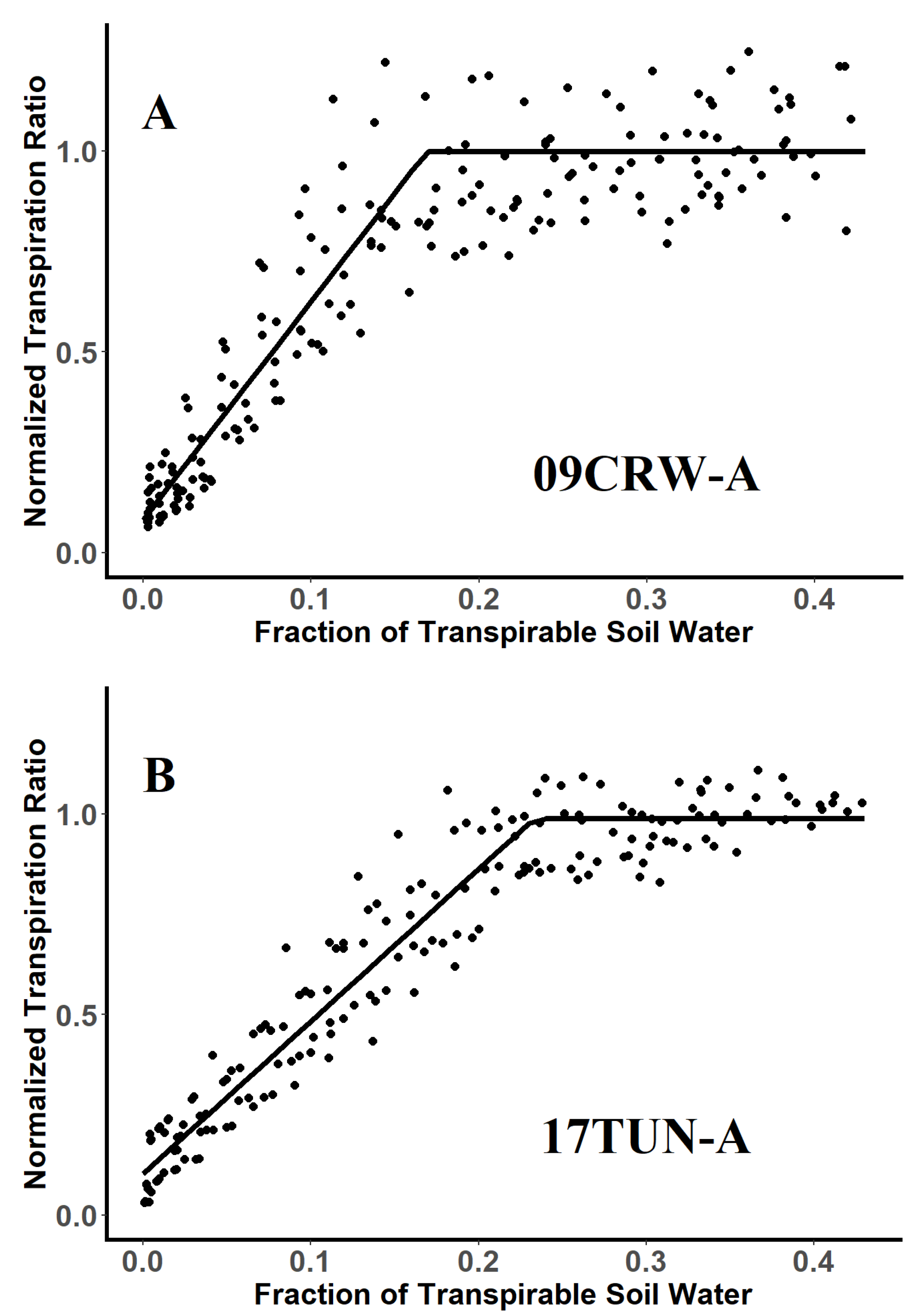
3.1. Differential Response of S-Metolachlor-Resistant and -Suceptible Accessions to Progressive Drying
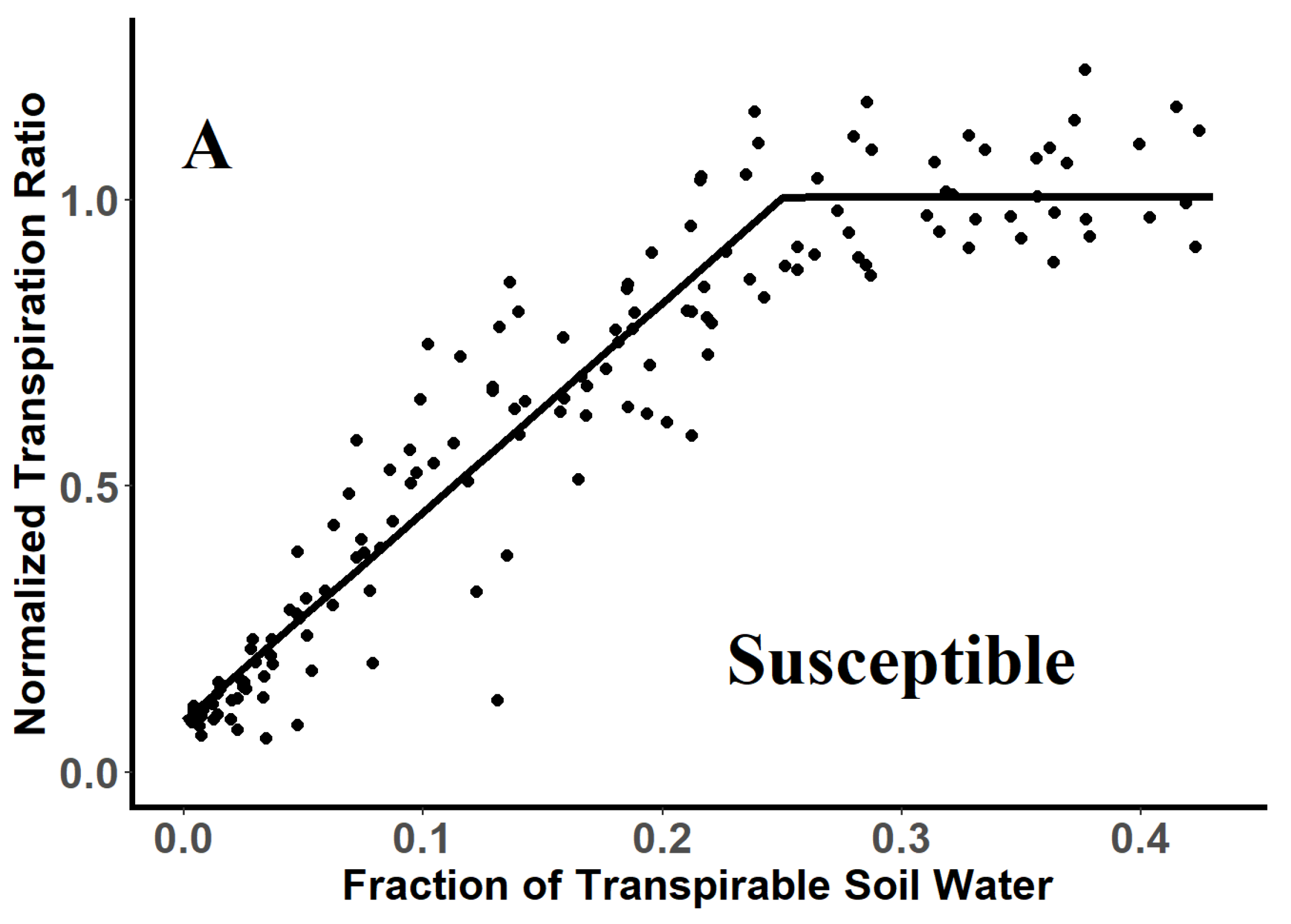
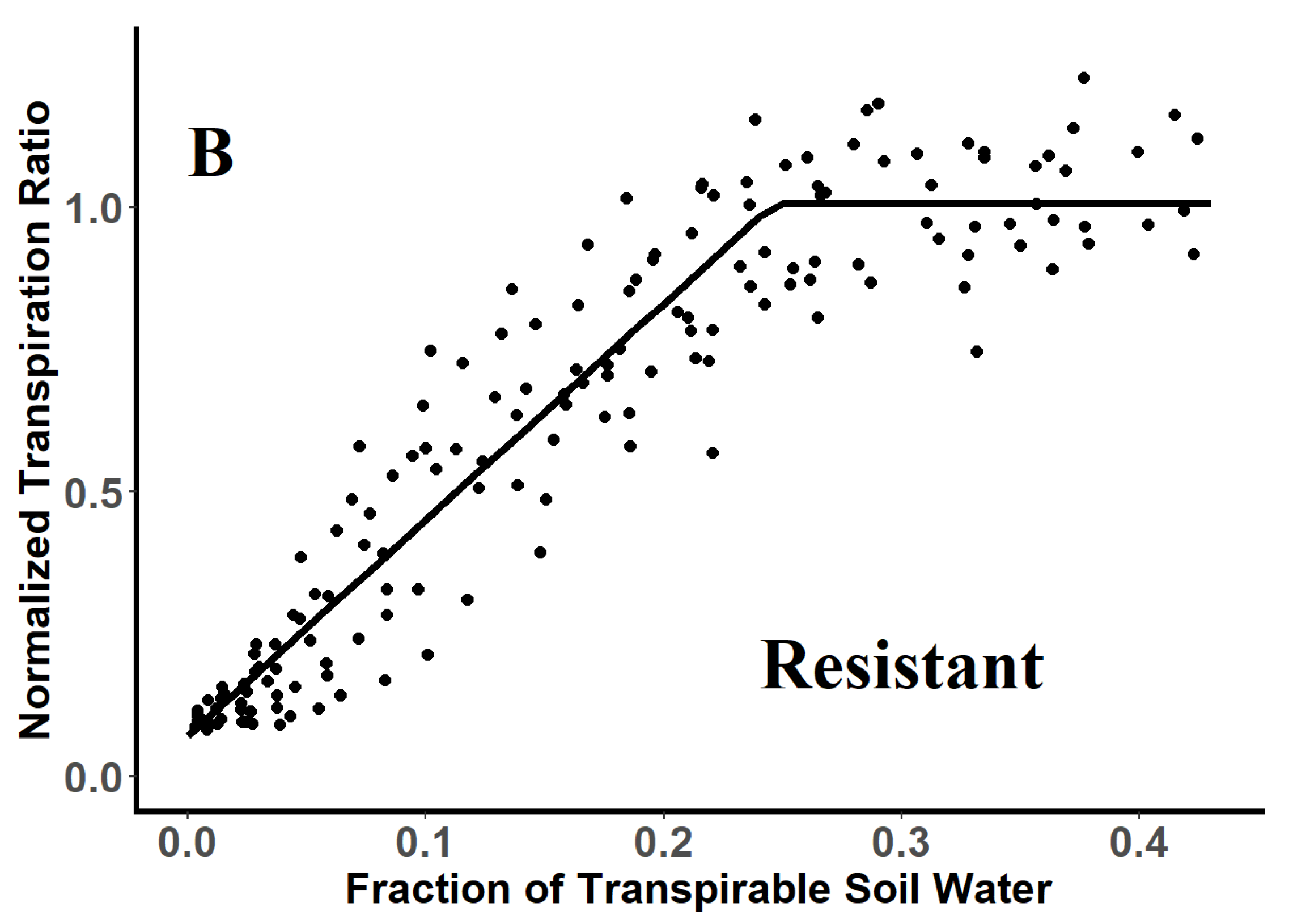
3.2. Threshold Values for Stomatal Closure of Glyphosate-Resistant and -Susceptible Plants
4. Discussion
4.1. Differential Response of S-Metolachlor-Resistant and -Suceptible Accessions to Daily Transpiration
4.2. No Difference in Threshold Values for Stomatal Closure between Glyphosate-Resistant and -Susceptible Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Khan, A.; Pan, X.D.; Najeeb, U.; Tan, D.K.Y.; Fahad, S.; Zahoor, R.; Luo, H.H. Coping with drought: Stress and adaptive mechanisms, and management through cultural and molecular alternatives in cotton as vital constituents for plant stress resilience and fitness. Biol. Res. 2018, 51, 47. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.S.; Byrne, P.; Cassman, K.G.; Cooper, M.; Delmer, D.; Greene, T.; Gruis, F.; Habben, J.; Hausmann, N.; Kenny, N.; et al. The US drought of 2012 in perspective: A call to action. Glob. Food Secur.-Agric. Policy Econ. Environ. 2013, 2, 139–143. [Google Scholar] [CrossRef]
- Jin, C.; Luo, X.; Xiao, X.M.; Dong, J.W.; Li, X.M.; Yang, J.; Zhao, D.Y. The 2012 Flash Drought Threatened US Midwest Agroecosystems. Chin. Geogr. Sci. 2019, 29, 768–783. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Basso, B. Impacts of climate variability and adaptation strategies on crop yields and soil organic carbon in the US Midwest. PLoS ONE 2020, 15, e0225433. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Deines, J.M.; Di Tommaso, S. Changes in the drought sensitivity of US maize yields. Nat. Food 2020, 1, 729–735. [Google Scholar] [CrossRef]
- Bowling, L.C.; Cherkauer, K.A.; Lee, C.I.; Beckerman, J.L.; Brouder, S.; Buzan, J.R.; Doering, O.C.; Dukes, J.S.; Ebner, P.D.; Frankenberger, J.R.; et al. Agricultural impacts of climate change in Indiana and potential adaptations. Clim. Chang. 2020, 163, 2005–2027. [Google Scholar] [CrossRef]
- Kropff, M.; van Laar, H. Modelling Crop-Weeds Interactions; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Kimm, H.; Guan, K.Y.; Gentine, P.; Wu, J.; Bernacchi, C.J.; Sulman, B.N.; Griffis, T.J.; Lin, C.J. Redefining droughts for the US Corn Belt: The dominant role of atmospheric vapor pressure deficit over soil moisture in regulating stomatal behavior of Maize and Soybean. Agric. For. Meteorol. 2020, 287, 107930. [Google Scholar] [CrossRef]
- Grassini, P.; Torrion, J.A.; Yang, H.S.; Rees, J.; Andersen, D.; Cassman, K.G.; Specht, J.E. Soybean yield gaps and water productivity in the western US Corn Belt. Field Crops Res. 2015, 179, 150–163. [Google Scholar] [CrossRef]
- Yang, Y.; Anderson, M.C.; Gao, F.; Johnson, D.M.; Sun, L.; Dulaney, W.; Hain, C.R.; Otkin, J.A.; Prueger, J.; Meyers, T.P.; et al. Phenological corrections to a field-scale, ET-based crop stress indicator: An application to yield forecasting across the US Corn Belt. Remote Sens. Environ. 2021, 257, 112337. [Google Scholar] [CrossRef]
- Li, L.J.; Song, X.Y.; Xia, L.; Fu, N.; Feng, D.; Li, H.Y.; Li, Y.L. Modelling the effects of climate change on transpiration and evaporation in natural and constructed grasslands in the semi-arid Loess Plateau, China. Agric. Ecosyst. Environ. 2020, 302, 107077. [Google Scholar] [CrossRef]
- Saradadevi, R.; Palta, J.A.; Siddique, K.H.M. ABA-mediated stomatal response in regulating water use during the development of terminal drought in wheat. Front. Plant Sci. 2017, 8, 743–764. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R. Is transpiration efficiency a viable plant trait in breeding for crop improvement? Funct. Plant Biol. 2012, 39, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cathey, S.E.; Kruse, J.K.; Sinclair, T.R.; Dukes, M.D. Transpiration and visual appearance of warm season turfgrasses during soil drying. Environ. Exp. Bot. 2013, 89, 36–43. [Google Scholar] [CrossRef]
- Sinclair, T.; Ludlow, M. Influence of soil-water supply on the plant water-balance of 4 tropical grain legumes. Aust. J. Plant Physiol. 1986, 13, 329–341. [Google Scholar] [CrossRef]
- Ray, J.; Gesch, R.; Sinclair, T.; Allen, L. The effect of vapor pressure deficit on maize transpiration response to a drying soil. Plant Soil 2002, 239, 113–121. [Google Scholar] [CrossRef]
- Ray, J.; Sinclair, T. Stomatal closure of maize hybrids in response to drying soil. Crop Sci. 1997, 37, 803–807. [Google Scholar] [CrossRef]
- Ray, J.; Samson, B.; Sinclair, T. Vegetative growth and soil water extraction of two maize hybrids during water deficits. Field Crops Res. 1997, 52, 135–142. [Google Scholar] [CrossRef]
- Ray, J.; Sinclair, T. The effect of pot size on growth and transpiration of maize and soybean during water deficit stress. J. Exp. Bot. 1998, 49, 1381–1386. [Google Scholar] [CrossRef]
- Bravo, W.; Leon, R.; Ferrell, J.; Mulvaney, M.; Wood, C. Evolutionary adaptations of Palmer amaranth (Amaranthus palmeri) to nitrogen fertilization and crop rotation history affect morphology and nutrient-use efficiency. Weed Sci. 2018, 66, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Ehleringer, J. Ecophysiology of Amaranthus palmeri: A Sonoran desert summer annual. Oecologia 1983, 57, 107–112. [Google Scholar] [CrossRef]
- Butts, T.R.; Vieira, B.C.; Latorre, D.O.; Werle, R.; Kruger, G.R. Competitiveness of herbicide-resistant waterhemp (Amaranthus tuberculatus) with soybean. Weed Sci. 2018, 66, 729–737. [Google Scholar] [CrossRef]
- Tardif, F.J.; Rajcan, I.; Costea, M. A mutation in the herbicide target site acetohydroxyacid synthase produces morphological and structural alterations and reduces fitness in Amaranthus powellii. New Phytol. 2006, 169, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Montellano, P. Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Plenum Publishers: New York, NY, USA, 2005. [Google Scholar]
- Singh, S.; Singh, V.; Lawton-Rauh, A.; Bagavathiannan, M.; Roma-Burgos, N. EPSPS gene amplification primarily confers glyphosate resistance among Arkansas Palmer amaranth (Amaranthus palmeri) populations. Weed Sci. 2018, 66, 293–300. [Google Scholar] [CrossRef]
- Rangani, G.; Noguera, M.; Salas-Perez, R.; Benedetti, L.; Roma-Burgos, N. Mechanism of resistance to S-metolachlor in Palmer amaranth. Front. Plant Sci. 2021, 12, 13. [Google Scholar] [CrossRef]
- King, C.; Purcell, L. Evaluation of methods for estimating transpiration response to soil drying for container-grown plants. Crop Sci. 2017, 57, 2143–2148. [Google Scholar] [CrossRef]
- Schmidt, J.; Blankenship, E.; Lindquist, J. Corn and velvetleaf (Abutilon theophrasti) transpiration in response to drying soil. Weed Sci. 2011, 59, 50–54. [Google Scholar] [CrossRef] [Green Version]
- King, C.; Purcell, L.; Brye, K. Differential wilting among soybean genotypes in response to water deficit. Crop Sci. 2009, 49, 290–298. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, 3.5.1; R Foundation for Statistical Computin: Vienna, Austria, 2018. [Google Scholar]
- Sinclair, T.R.; Manandhar, A.; Belko, N.; Riar, M.; Vadez, V.; Roberts, P.A. Variation among cowpea genotypes in sensitivity of transpiration rate and symbiotic nitrogen fixation to soil drying. Crop Sci. 2015, 55, 2270–2275. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Shekoofa, A.; Isleib, T.G.; Balota, M.; Zhang, H. Identification of Virginia-type peanut genotypes for water-deficit conditions based on early decrease in transpiration rate with soil drying. Crop Sci. 2018, 58, 2607–2612. [Google Scholar] [CrossRef]
- Devi, M.J.; Reddy, V. Cotton genotypic variability for transpiration decrease with progressive soil drying. Agronomy 2020, 10, 1290. [Google Scholar] [CrossRef]
- Fuentealba, M.P.; Zhang, J.; Kenworthy, K.; Erickson, J.; Kruse, J.; Trenholm, L. Transpiration responses of warm-season turfgrass in relation to progressive soil drying. Sci. Hortic. 2016, 198, 249–253. [Google Scholar] [CrossRef]
- Miller, G.L. Physiological response of bermudagrass grown in soil amendments during drought stress. Hortscience 2000, 35, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Devi, M.J.; Sinclair, T.R.; Vadez, V.; Krishnamurthy, L. Peanut genotypic variation in transpiration efficiency and decreased transpiration during progressive soil drying. Field Crops Res. 2009, 114, 280–285. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jo, H.; Kong, K. A plant-specific tau class glutathione S-transferase from Oryza sativa having significant detoxification activity towards chloroacetanilide herbicides. Bull. Korean Chem. Soc. 2011, 32, 3756–3759. [Google Scholar] [CrossRef] [Green Version]
- George, S.; Venkataraman, G.; Parida, A. A chloroplast-localized and auxin-induced glutathione S-transferase from phreatophyte Prosopis juliflora confer drought tolerance on tobacco. J. Plant Physiol. 2010, 167, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Zhu, Y.M.; Li, Y.; Yang, L.A.; Zhao, X.W.; Cai, H.; Bai, X. Over-expression of a glutathione S-transferase gene, GsGST, from wild soybean (Glycine soja) enhances drought and salt tolerance in transgenic tobacco. Biotechnol. Lett. 2010, 32, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, Y.; Rao, J.; Wang, G.; Li, H.; Ge, F.; Chen, C. Overexpression of the glutathione S-transferase gene from Pyrus pyrifolia fruit improves tolerance to abiotic stress in transgenic tobacco plants. Mol. Biol. 2013, 47, 515–523. [Google Scholar] [CrossRef]
- Xu, J.; Zheng, A.Q.; Xing, X.J.; Chen, L.; Fu, X.Y.; Peng, R.H.; Tian, Y.S.; Yao, Q.H. Transgenic Arabidopsis plants expressing grape glutathione S-transferase gene (VvGSTF13) show enhanced tolerance to abiotic stress. Biochemistry 2018, 83, 755–765. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, Y.J.; Zeng, Q.Y. Overexpression of three orthologous glutathione S-transferases from Populus increased salt and drought resistance in Arabidopsis. Biochem. Syst. Ecol. 2019, 83, 57–61. [Google Scholar] [CrossRef]
- Osipitan, O.A.; Dille, J.A. No Impact of Increased EPSPS Gene Copy Number on Growth and Fecundity of Glyphosate-Resistant Kochia (Bassia scoparia). Weed Sci. 2019, 67, 22–28. [Google Scholar] [CrossRef]
- Giacomini, D.; Westra, P.; Ward, S.M. Impact of genetic background in fitness cost studies: An example from glyphosate-resistant Palmer amaranth. Weed Sci. 2014, 62, 29–37. [Google Scholar] [CrossRef]
- Vila-Aiub, M.; Goh, S.; Gaines, T.; Han, H.; Busi, R.; Yu, Q.; Powles, S. No fitness cost of glyphosate resistance endowed by massive EPSPS gene amplification in Amaranthus palmeri. Planta 2014, 239, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Lorraine-Colwill, D.F.; Powles, S.B.; Hawkes, T.R.; Hollinshead, P.H.; Warner, S.A.J.; Preston, C. Investigations into the mechanism of glyphosate resistance in Lolium rigidum. Pestic. Biochem. Physiol. 2002, 74, 62–72. [Google Scholar] [CrossRef]
- Pedersen, B.; Neve, P.; Andreasen, C.; Powles, S. Ecological fitness of a glyphosate-resistant Lolium rigidum population: Growth and seed production along a competition gradient. Basic Appl. Ecol. 2007, 8, 258–268. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | Breakpoint a | SE | R2 | Confidence Intervals b | |
|---|---|---|---|---|---|
| 09CRW-A | 0.17 a | 0.007 | 0.85 | 0.15 | 0.19 |
| 17TUN-A | 0.23 b | 0.004 | 0.93 | 0.22 | 0.25 |
| Genotype | Breakpoint a | SE | R2 | Confidence Intervals b | |
|---|---|---|---|---|---|
| Glyphosate-resistant | 0.25 a | 0.007 | 0.90 | 0.23 | 0.26 |
| Glyphoste-susceptible | 0.25 a | 0.008 | 0.91 | 0.23 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouame, K.B.-J.; Savin, M.C.; Rangani, G.; Butts, T.R.; Bertucci, M.B.; Roma-Burgos, N. Transpiration Responses of Herbicide-Resistant and -Susceptible Palmer Amaranth (Amaranthus palmeri (S.) Wats.) to Progressively Drying Soil. Agriculture 2022, 12, 335. https://doi.org/10.3390/agriculture12030335
Kouame KB-J, Savin MC, Rangani G, Butts TR, Bertucci MB, Roma-Burgos N. Transpiration Responses of Herbicide-Resistant and -Susceptible Palmer Amaranth (Amaranthus palmeri (S.) Wats.) to Progressively Drying Soil. Agriculture. 2022; 12(3):335. https://doi.org/10.3390/agriculture12030335
Chicago/Turabian StyleKouame, Koffi Badou-Jeremie, Mary C. Savin, Gulab Rangani, Thomas R. Butts, Matthew B. Bertucci, and Nilda Roma-Burgos. 2022. "Transpiration Responses of Herbicide-Resistant and -Susceptible Palmer Amaranth (Amaranthus palmeri (S.) Wats.) to Progressively Drying Soil" Agriculture 12, no. 3: 335. https://doi.org/10.3390/agriculture12030335