Simultaneous Detection of Plant- and Fungus-Derived Genes Constitutively Expressed in Single Pseudoidium neolycopersici-Inoculated Type I Trichome Cells of Tomato Leaves via Multiplex RT-PCR and Nested PCR

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Fungal Materials
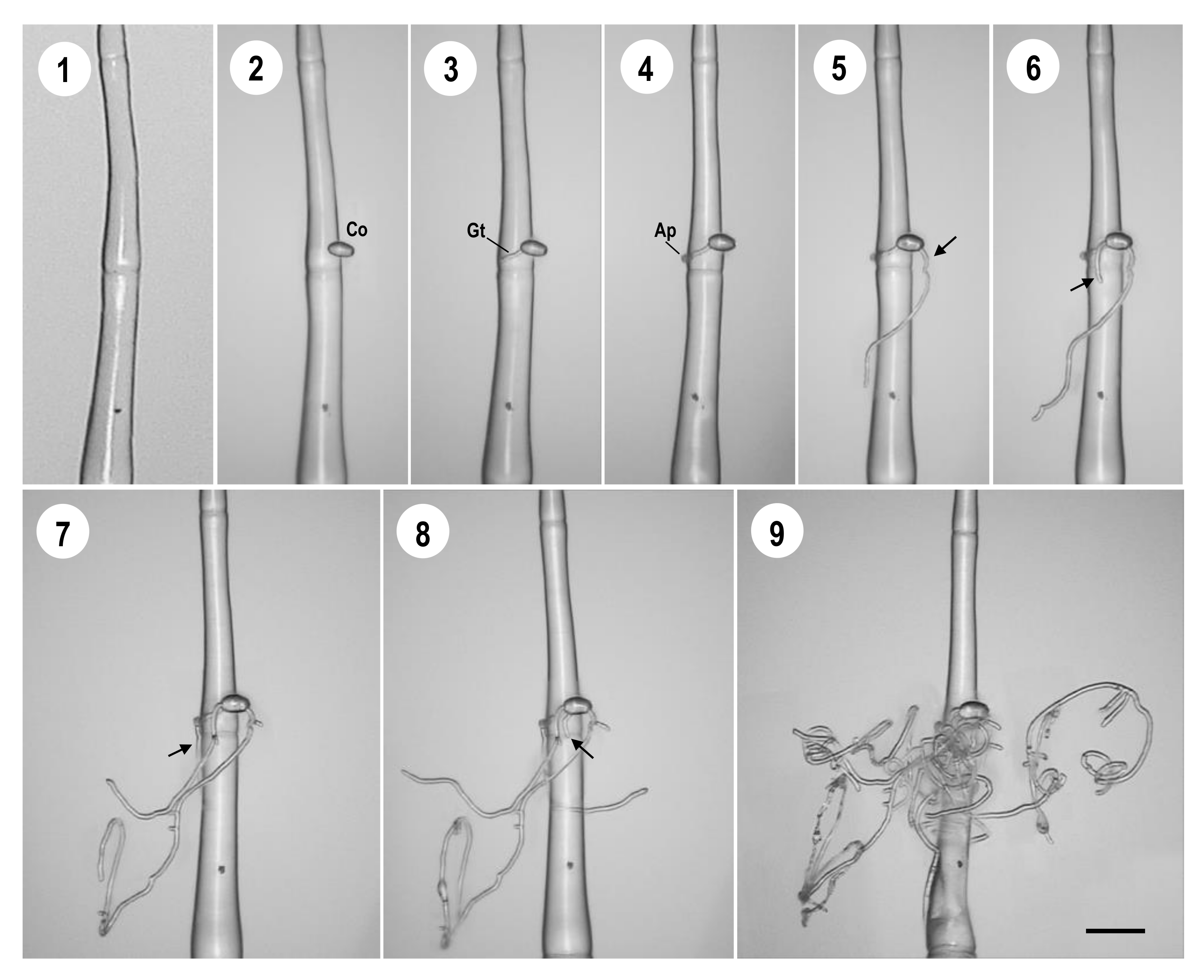
2.3. Dynamic Analysis of P. neolycopersici Infection in Tomato Leaf Type I Trichomes
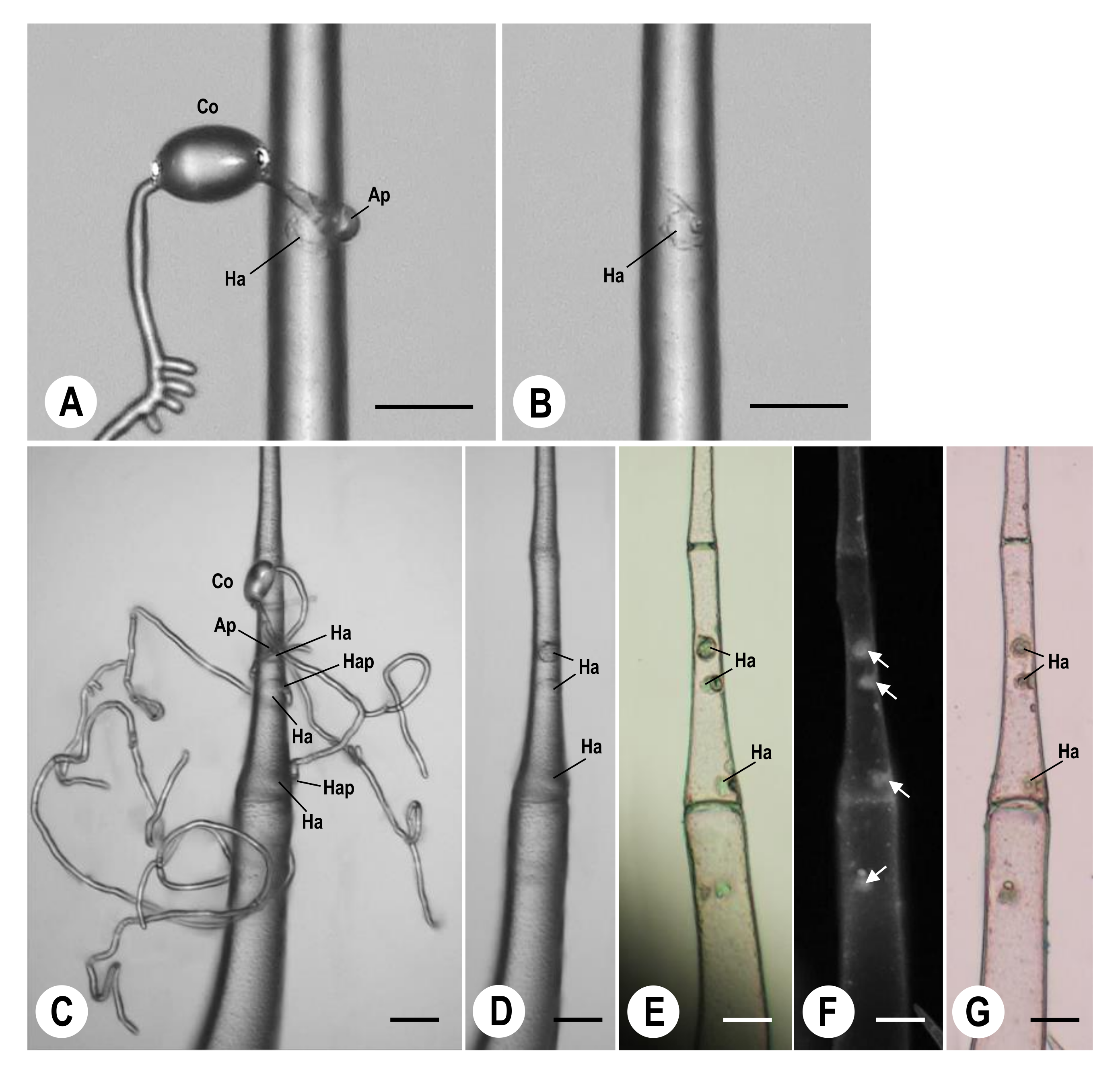
2.4. Microscopic Observation of P. neolycopersici Haustoria Formed in Tomato Leaf Type I Trichomes
2.5. Collection of Single P. neolycopersici-Inoculated Tomato Leaf Type I Trichomes with Microscissors
2.6. Amplification of 5.8S rDNA-ITS Sequence by PCR
2.7. Transcript Amplification of Tomato Plant- and Powdery Mildew Fungus-Derived Genes by suc-PCR (RT-PCR/N-PCR)
2.8. Simultaneous Transcript Amplification of Tomato Plant- and Powdery Mildew Fungus-Derived Genes by Multiplex RT-PCR/N-PCR
3. Results
3.1. Analysis of Infection Processes of P. neolycopersici on Leaf Type I Trichome Cells of MM
3.2. Collection of Single P. neolycopersici-Inoculated Type I Trichomes Using Microscissors
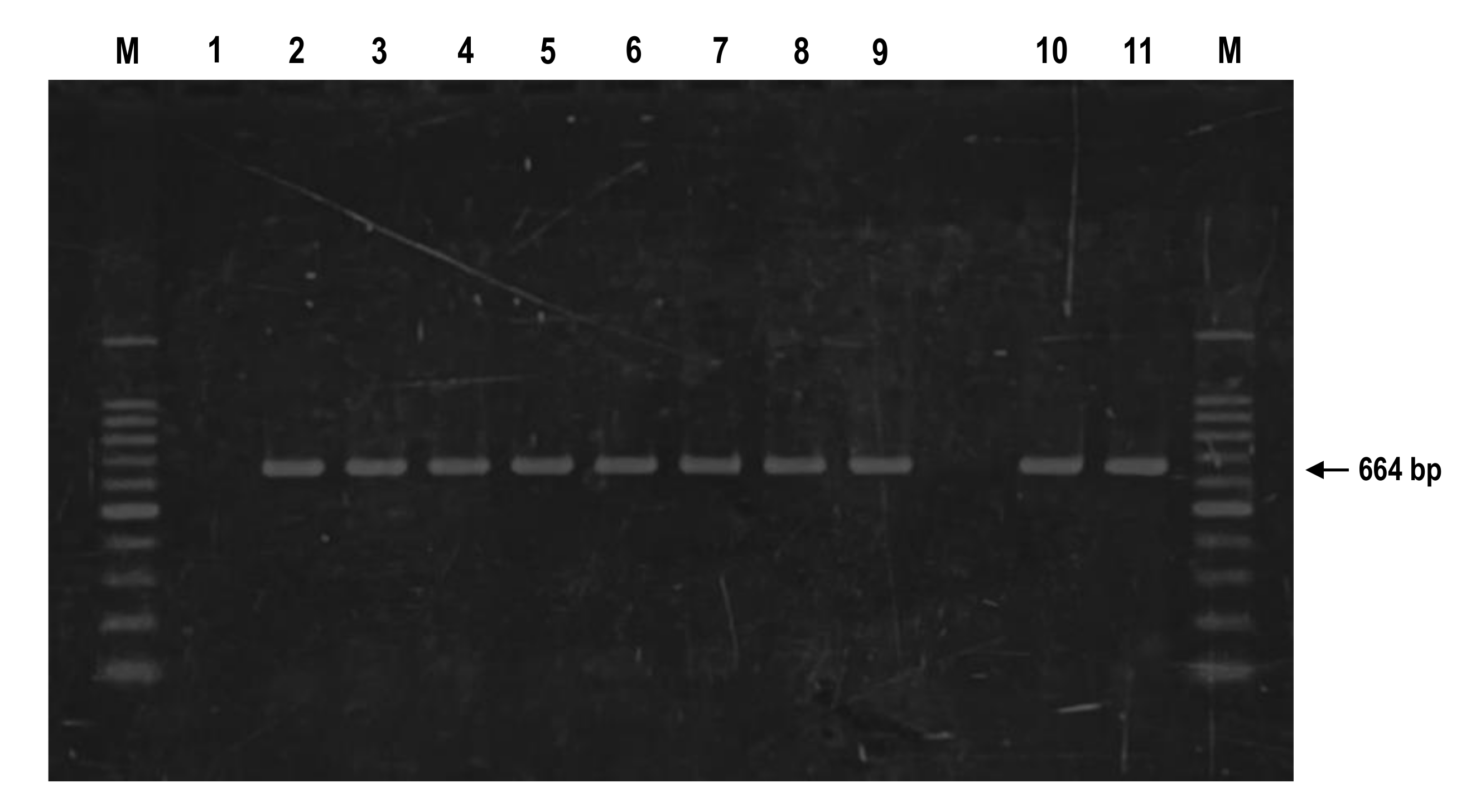
3.3. Amplification of Fungal 5.8S rDNA-ITS Sequences by PCR from Single P. neolycopersici-Inoculated Type I Trichome Cells
3.4. Transcript Detection of Plant- and Fungus-Derived Genes by suc-PCR (RT-PCR/N-PCR) from Single P. neolycopersici-Inoculated Type I Trichome Cells
3.5. Simultaneous Transcript Detection of Plant- and Fungus-Derived Genes by Multiplex RT-PCR/N-PCR from Single P. neolycopersici-Inoculated Type I Trichome Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, L.; Zhu, L.; Tu, L.; Guo, X.; Long, L.; Sun, L.; Gao, W.; Zhang, X. Differential gene expression in cotton defence response to Verticillium dahlia by SSH. J. Phytopathol. 2011, 159, 684–697. [Google Scholar] [CrossRef]
- Mohamed, I.A.; Shalby, N.; El-Badri, A.M.A.; Saleem, M.H.; Khan, M.N.; Nawaz, M.A.; Qin, M.; Agami, R.A.; Kuai, J.; Wang, B. Stomata and xylem vessels traits improved by melatonin application contribute to enhancing salt tolerance and fatty acidcomposition of Brassica napus L. plants. Agronomy 2020, 10, 1186. [Google Scholar] [CrossRef]
- Sudisha, J.; Sharathchandra, R.; Amruthesh, K.; Kumar, A.; Shetty, H.S. Pathogenesis related proteins in plant defense response. In Plant Defence: Biological Control; Springer: Berlin/Heidelberg, Germany, 2012; pp. 379–403. [Google Scholar]
- Munis, M.F.H.; Tu, L.; Deng, F.; Tan, J.; Xu, L.; Xu, S.; Long, L.; Zhang, X. A thaumatin-like protein gene involved in cotton fiber secondary cell wall development enhances resistance against Verticillium dahliae and other stresses in transgenic tobacco. Biochem. Biophys. Res. Commun. 2010, 393, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, S.; Usha, K.; Singh, B. Pathogenesis related (PR) proteins in plant defense mechanism. Sci. Against Microb. Pathog. 2011, 2, 1043–1054. [Google Scholar]
- Lawrence, C.B.; Singh, N.P.; Qiu, J.; Gardner, R.G.; Tuzun, S. Constitutive hydrolytic enzymes are associated with polygenic resistance of tomato to Alternaria solani and may function as an elicitor release mechanism. Physiol. Mol. Plant Pathol. 2000, 57, 211–220. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3-glucans are elicitors of defense response in tobacco. Plant Physiol. 2000, 124, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-Sommergruber, K. Plant allergens and pathogenesis-related proteins. Int. Arch. Allergy Immunol. 2000, 122, 155–166. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Ullah, A.; Gul, S.; Shaban, M.; Khan, A.H.; Ali, M.; Sani, S.G.A.S.; Chaudhary, H.J.; Munis, M.F.H. Expression analysis of defense related genes in wheat and maize against Bipolaris sorokiniana. Physiol. Mol. Plant Pathol. 2018, 103, 36–46. [Google Scholar] [CrossRef]
- Sunera; Amna; Saqib, S.; Uddin, S.; Zaman, W.; Ullah, F.; Ayaz, A.; Asghar, M.; ur Rehman, S.; Munis, M.F.H.; et al. Characterization and phytostimulatory activity of bacteria isolated from tomato (Lycopersicin esculentum Mill.) rhizosphere. Microb. Pathog. 2020, 140, 103966. [Google Scholar]
- Manghwar, H.; Hussain, A.; Ali, Q.; Saleem, M.H.; Abualreesh, M.H.; Alatawi, A.; Ali, S.; Munis, M.F.H. Disease severity, resistance analysis, and expression profiling of pathogenesis-related protein genes after the inoculation of Fusarium equiseti in wheat. Agronomy 2021, 11, 2124. [Google Scholar] [CrossRef]
- White, J.F., Jr.; Johnston, S.A.; Wang, C.L.; Chin, C.K. First report of powdery mildew in greenhouse-grown tomatoes in New Jersey. Plant Dis. 1997, 81, 227. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; Cook, R.T.A.; Saenz, G.S.; Cunnington, J.H.; Takamatsu, S.; Pascoe, I.; Bardin, M.; Nicot, P.C.; Sato, Y.; Rossman, A.Y. Identification of two powdery mildew fungi, Oidium neolycopersici sp. nov. and O. lycopersici, infecting tomato in different parts of the world. Mycol. Res. 2001, 105, 684–697. [Google Scholar] [CrossRef]
- Zheng, Z.; Nonomura, T.; Bóka, K.; Matsuda, Y.; Visser, R.G.F.; Toyoda, H.; Kiss, L.; Bai, Y. Detection and quantification of Leveillula taurica growth in pepper leaves. Phytopathology 2013, 103, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Braun, U.; Shin, H.D.; Takamatsu, S.; Meeboon, J.; Kiss, L.; Lebeda, A.; Kitner, M.; Götz, M. Phylogeny and taxonomy of Golovinomyces orontii revisited. Mycol. Prog. 2019, 18, 335–357. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kashimoto, K.; Takikawa, Y.; Aikami, R.; Nonomura, T.; Toyoda, H. Occurrence of new powdery mildew on greenhouse tomato cultivars. J. Gen. Plant Pathol. 2001, 67, 294–298. [Google Scholar] [CrossRef]
- Shimizu, K.; Matsuda, Y.; Nonomura, T.; Ikeda, H.; Tamura, N.; Kusakari, S.; Kimbara, J.; Toyoda, H. Dual protection of hydroponic tomatoes from rhizosphere pathogens Ralstonia solanacearum and Fusarium oxysporum f. sp. radicis-lycopersici and airborne conidia of Oidium neolycopersici with an ozone-generative electrostatic spore precipitator. Plant Pathol. 2007, 56, 987–997. [Google Scholar] [CrossRef]
- Nonomura, T.; Matsuda, Y.; Kakutani, K.; Takikawa, Y.; Toyoda, H. Physical control of powdery mildew (Oidium neolycopersici) on tomato leaves by exposure to corona discharge. Can. J. Plant Pathol. 2008, 30, 517–524. [Google Scholar] [CrossRef]
- Kashimoto, K.; Matsuda, Y.; Matsutani, K.; Sameshima, T.; Kakutani, K.; Nonomura, T.; Okada, K.; Kusakari, S.; Nakata, K.; Takamatsu, S.; et al. Morphological and molecular characterization for a Japanese isolate of tomato powdery mildew Oidium neolycopersici and its host range. J. Gen. Plant Pathol. 2003, 69, 176–185. [Google Scholar]
- Nonomura, T.; Nishitomi, A.; Matsuda, Y.; Soma, C.; Xu, L.; Kakutani, K.; Takikawa, Y.; Toyoda, H. Polymorphic change of appressoria by the tomato powdery mildew Oidium neolycopersici on host tomato leaves reflects multiple unsuccessful penetration attempts. Fungal Biol. 2010, 114, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, T.; Matsuda, Y.; Yamashita, S.; Akahoshi, H.; Takikawa, Y.; Kakutani, K.; Toyoda, H. Natural woody plant, Mallotus japonicus, as an ecological partner to transfer different pathotypic conidia of Oidium neolycopersici to greenhouse tomatoes. Plant Prot. Sci. 2013, 49, S33–S40. [Google Scholar] [CrossRef] [Green Version]
- Seifi, A.; Nonomura, T.; Matsuda, Y.; Toyoda, H.; Bai, Y. An avirulent tomato powdery mildew isolate induces localized acquired resistance to a virulent isolate in a spatiotemporal manner. Mol. Plant-Microbe Interact. 2012, 25, 372–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, H.; Whipps, J.M.; Gurr, S.J. The tomato powdery mildew fungus Oidium neolycopersici. Mol. Plant Pathol. 2001, 2, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Jacob, D.; David, D.R.; Sztjenberg, A.; Elad, Y. Conditions for development of powdery mildew of tomato caused by Oidium neolycopersici. Phytopathology 2008, 98, 270–281. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, G.G.; Sorenson, C.F. Role of glandular trichomes in the resistance of Lycopersicon hirsutum f. glabratum to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1985, 78, 547–551. [Google Scholar] [CrossRef]
- Duffey, S.S. Plant glandular trichomes: Their partial role in defense against insects. In Insects and the Plant Surface; Juniper, B.E., Southwood, T.E., Eds.; Arnold: London, UK, 1986; pp. 151–172. [Google Scholar]
- Peter, A.J.; Shanower, T.G. Plant glandular trichomes: Chemical factories with many potential uses. Resonance 1998, 3, 41–45. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Simmons, A.T.; Gurr, G.M. Trichomes of Lycopersicon species and their hybrids: Effects on pests and natural enemies. Agric. For. Entomol. 2005, 7, 265–276. [Google Scholar] [CrossRef]
- Nonomura, T.; Xu, L.; Wada, M.; Kawamura, S.; Miyajima, T.; Nishitomi, A.; Kakutani, K.; Takikawa, Y.; Matsuda, Y.; Toyoda, H. Trichome exudates of Lycopersicon pennellii form a chemical barrier to suppress leaf-surface germination of Oidium neolycopersici conidia. Plant Sci. 2009, 176, 31–37. [Google Scholar] [CrossRef]
- Kang, J.-H.; Liu, G.; Shi, F.; Jones, A.D.; Beaudry, R.M.; Howe, G.A. The tomato odorless-2 mutant is defective in trichome-based production of diverse specialized metabolites and broad-spectrum resistance to insect herbivores. Plant Physiol. 2010, 154, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Tooker, J.F.; Peiffer, M.; Luthe, D.S.; Felton, G.W. Trichomes as sensors detecting activity on the leaf surface. Plant Signal. Behav. 2010, 5, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Lemke, C.A.; Mutschler, M.A. Inheritance of glandular trichomes in crosses between Lycopersicon esculentum and L. pennellii. J. Am. Soc. Hort. Sci. 1984, 109, 592–596. [Google Scholar]
- Glover, B.J. Differentiation in plant epidermal cells. J. Exp. Bot. 2000, 51, 497–505. [Google Scholar] [CrossRef] [PubMed]
- McDowell, E.T.; Kapteyn, J.; Schmidt, A.; Li, C.; Kang, J.H.; Descour, A.; Shi, F.; Larson, M.; Schilmiller, A.; An, L.; et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol. 2011, 155, 524–539. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Murakami, T.; Takizumi, Y.; Ishimaru, H.; Kudo, D.; Takikawa, Y.; Matsuda, Y.; Kakutani, K.; Bai, Y.; Nonomura, T. Trichomes: Interaction sites of tomato leaves with biotrophic powdery mildew pathogens. Eur. J. Plant Pathol. 2018, 150, 115–125. [Google Scholar] [CrossRef]
- Suzuki, T.; Nakao, Y.; Ura, R.; Nakaba, M.; Nishiyama, T.; Takikawa, Y.; Matsuda, Y.; Kakutani, K.; Bai, Y.; Nonomura, T. Correlation between total hypha length and haustoria number of Pseudoidium neolycopersici in type I trichome cells of tomato leaves. Austral. Plant Pathol. 2020, 49, 451–460. [Google Scholar] [CrossRef]
- Fujita, K.; Matsuda, Y.; Wada, M.; Hirai, Y.; Mori, K.; Moriura, N.; Nonomura, T.; Kakutani, K.; Toyoda, H. Powdery mildew pathogens can suppress the chitinase gene expression induced in detached inner epidermis of barley coleoptile. Plant Cell Rep. 2004, 23, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Chandran, D.; Inada, N.; Hather, G.; Kleindt, C.K.; Wildermuth, M.C. Laser microdissection of Arabidopsis cells at the powdery mildew infection site reveals site-specific processes and regulators. Proc. Natl. Acad. Sci. USA 2010, 107, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular cloning and functional analysis of GmLACS2-3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Nonomura, T.; Matsuda, Y.; Xu, L.; Kakutani, K.; Takikawa, Y.; Toyoda, H. Collection of highly germinatve pseudochain conidia of Oidium neolycopersici from conidiophores by electrostatic attraction. Mycol. Res. 2009, 113, 364–372. [Google Scholar] [CrossRef]
- Sameshima, T.; Kashimoto, K.; Kida, K.; Matsuda, Y.; Nonomura, T.; Kakutani, K.; Nakata, K.; Kusakari, S.; Toyoda, H. Cytological events in tomato leaves inoculated with conidia of Blumeria graminis f. sp. hordei and Oidium neolycopersici KTP-01. J. Gen. Plant Pathol. 2004, 70, 7–10. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Wada, M.; Matsuda, Y.; Fujita, K.; Nanjo, A.; Nishimura, M.; Nonomura, T.; Kakutani, K.; Toyoda, H. RT-PCR amplification of mRNAs in nuclei or cytosol of single cells of tomato callus after micropipette extraction. Plant Cell Tissue Organ Cult. 2004, 79, 109–114. [Google Scholar] [CrossRef]
- Ewing, N.N.; Wimmers, L.E.; Meyer, D.J.; Chetelat, R.T.; Bennett, A.B. Molecular cloning of tomato plasma membrane H+-ATPase. Plant Physiol. 1990, 94, 1874–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, N.N.; Bennett, A.B. Assessment of the number and expression of P-type H+-ATPase genes in tomato. Plant Physiol. 1994, 106, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebeda, A.; Mieslerová, B.; Petřivalský, M.; Luhová, L.; Špundová, M.; Sedlářová, M.; Nožková-Hlaváčková, V.; Pink, D.A.C. Resistance mechanisms of wild tomato germplasm to infection of Oidium neolycopersici. Eur. J. Plant Pathol. 2014, 138, 569–596. [Google Scholar] [CrossRef] [Green Version]
- Dyki, B.; Staniaszek, M. Infection of tomato by Oidium lycopersicum (Cooke & Massee, emend. Noordeloos & Loerakker). Phytopathol. Pol. 1997, 13, 13–17. [Google Scholar]
- Whipps, J.M.; Budge, S.P.; Fenlon, J.S. Characteristics and host range of tomato powdery mildew. Plant Pathol. 1998, 47, 36–48. [Google Scholar] [CrossRef]
- LaMondia, J.A.; Smith, V.L.; Douglas, S.M. Host range of Oidium lycopersicum on selected Solanaceous species in Connecticut. Plant Dis. 1999, 83, 341–344. [Google Scholar] [CrossRef]
- Li, C.; Faino, L.; Dong, L.; Fan, J.; Kiss, L.; de Giovanni, C.; Lebeda, A.; Scott, J.; Matsuda, Y.; Toyoda, H.; et al. Characterization of polygenic resistance to powdery mildew in tomato at cytological, biochemical and gene expression level. Mol. Plant Pathol. 2012, 13, 148–159. [Google Scholar] [CrossRef]
- Dyki, B. Morphogenesis of pathogen causing powdery mildew in Polish cultivation of tomato. Veg. Crop. Res. Bull. 2003, 59, 131–138. [Google Scholar]
- Oichi, W.; Matsuda, Y.; Sameshima, T.; Nonomura, T.; Kakutani, K.; Nishimura, H.; Kusakari, S.; Toyoda, H. Consecutive monitoring for conidiogenesis by Oidium neolycopersici on tomato leaves with a high-fidelity digital microscope. J. Gen. Plant Pathol. 2004, 70, 329–332. [Google Scholar] [CrossRef]
- Oichi, W.; Matsuda, Y.; Nonomura, T.; Toyoda, H.; Xu, L.; Kusakari, S. Formation of conidial pseudochains by tomato powdery mildew Oidium neolycopersici. Plant. Dis. 2006, 90, 915–919. [Google Scholar] [CrossRef] [Green Version]
- Pepper, I.L.; Dowd, S.E. PCR applications for plant and soil microbes. In Manual of Environmental Microbiology; Hurst, C.J., Crawford, R.L., Knudsen, G.R., McInerney, M.J., Stetzenbach, L.D., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 573–582. [Google Scholar]
- Llop, P.; Bonaterra, A.; Penalver, J.; Lopez, M.M. Development of a high sensitive nested-PCR procedure using a single closed tube for detection of Erwinia amylovora in asymptomatic plant material. Appl. Environ. Microbiol. 2000, 66, 2071–2078. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Sameshima, T.; Moriura, N.; Inoue, K.; Nonomura, T.; Kakutani, K.; Nishimura, H.; Kusakari, S.; Takamatsu, S.; Toyoda, H. Identification of individual powdery mildew fungi infecting leaves and direct detection of gene expression by single conidium polymerase chain reaction. Phytopathology 2005, 95, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Kakutani, K.; Nishimura, M.; Wada, M.; Nonomura, T.; Toyoda, H. Direct RT-PCR amplification of mature mRNAs in single trichome cells of plant leaves. Recent Res. Dev. Cell Biol. 2003, 1, 145–153. [Google Scholar]
- Janke, C.; Magiera, M.M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. 2020, 21, 307–326. [Google Scholar] [CrossRef]
- Ellis, J. Insights into nonhost disease resistance: Can they assist disease control in agriculture? Plant Cell 2006, 18, 523–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, P. Nonhost resistance of plants to powdery mildew—New opportunities to unravel the mystery. Physiol. Mol. Plant Pathol. 2007, 70, 3–7. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Targets | Primers | Primer Sequences |
|---|---|---|
| rDNA-ITS | ITS-4 a | 5′-TCCTCCGCTTATTGATATGC-3′ |
| ITS-5 a | 5′-GGAAGTAAAAGTCGTAACAAGG-3′ | |
| LHA2 | lpRT-1 b | 5′-GCCAAAGGTGTTGACGCAGATAC-3′ |
| lpRT-2 b | 5′-CACTCCAAGATTCAAAGCCCTCCT-3′ | |
| lpN-1 b | 5′-AGAACCAGGATGCCATTGAC-3′ | |
| lpN-2 b | 5′-ACAAAGGCAGGAGACCAATG-3′ | |
| TUB2 | tbRT-1 | 5′-AACCAAATTGGGGCTGCTTTC-3′ |
| tbRT-2 | 5′-CCCAGTTGTTACCAGCCCC-3′ | |
| tbN-1 | 5′-GCCTTGACGGATCTGGTGT-3′ | |
| tbN-2 | 5′-GTCGAAAGAGCTGACCGAA-3′ |
| PCR Methods | Target Genes | Rates of Gene Expression Detection (%) x | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 y | ||
| PCR/N-PCR | 5.8S rDNA-ITS | ND z | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| RT-PCR/N-PCR | LHA2 | 100.0 | 100.0 | 100.0 | 100.0 | 95.0 | 100.0 | 95.0 | 90.0 | 100.0 | 95.0 | 100.0 |
| TUB2 | ND | 80.0 | 85.0 | 100.0 | 85.0 | 100.0 | 95.0 | 100.0 | 100.0 | 100.0 | 100.0 | |
| Multiplex RT-PCR/N-PCR | LHA2 | 100.0 | 100.0 | 100.0 | 100.0 | 95.0 | 100.0 | 95.0 | 95.0 | 100.0 | 95.0 | 100.0 |
| TUB2 | ND | 85.0 | 90.0 | 100.0 | 95.0 | 100.0 | 95.0 | 85.0 | 100.0 | 100.0 | 100.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwasaki, S.; Okada, N.; Kimura, Y.; Takikawa, Y.; Suzuki, T.; Kakutani, K.; Matsuda, Y.; Bai, Y.; Nonomura, T. Simultaneous Detection of Plant- and Fungus-Derived Genes Constitutively Expressed in Single Pseudoidium neolycopersici-Inoculated Type I Trichome Cells of Tomato Leaves via Multiplex RT-PCR and Nested PCR. Agriculture 2022, 12, 254. https://doi.org/10.3390/agriculture12020254
Iwasaki S, Okada N, Kimura Y, Takikawa Y, Suzuki T, Kakutani K, Matsuda Y, Bai Y, Nonomura T. Simultaneous Detection of Plant- and Fungus-Derived Genes Constitutively Expressed in Single Pseudoidium neolycopersici-Inoculated Type I Trichome Cells of Tomato Leaves via Multiplex RT-PCR and Nested PCR. Agriculture. 2022; 12(2):254. https://doi.org/10.3390/agriculture12020254
Chicago/Turabian StyleIwasaki, Shota, Naoko Okada, Yutaka Kimura, Yoshihiro Takikawa, Tomoko Suzuki, Koji Kakutani, Yoshinori Matsuda, Yuling Bai, and Teruo Nonomura. 2022. "Simultaneous Detection of Plant- and Fungus-Derived Genes Constitutively Expressed in Single Pseudoidium neolycopersici-Inoculated Type I Trichome Cells of Tomato Leaves via Multiplex RT-PCR and Nested PCR" Agriculture 12, no. 2: 254. https://doi.org/10.3390/agriculture12020254