
Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits


Abstract
:1. Introduction
2. Materials and Methods
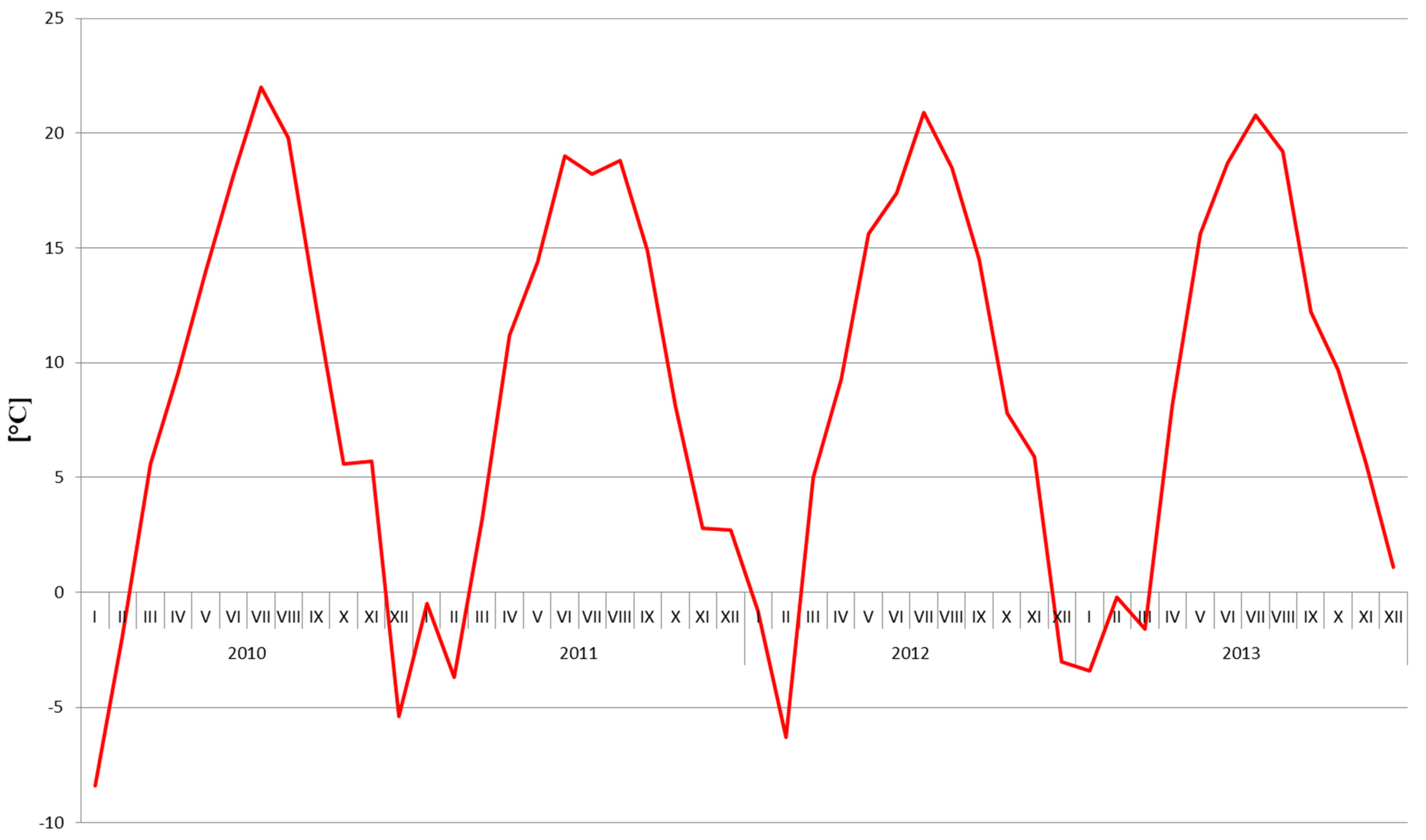
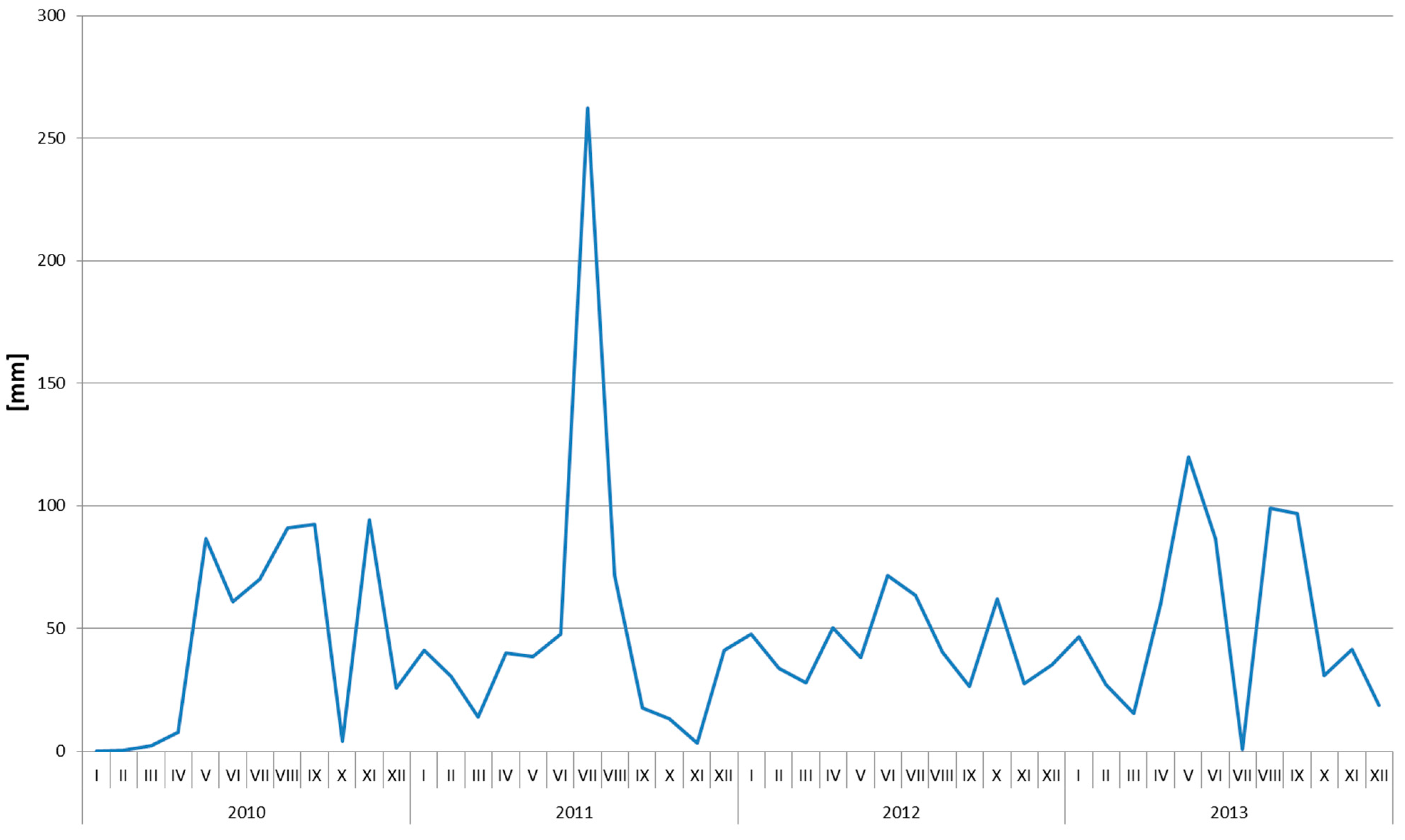
2.1. Soil Nitrogen Content
- (1)
- Before applying nitrogen fertilizer (April);
- (2)
- Five weeks after applying nitrogen fertilizer (May);
- (3)
- Nine weeks after applying nitrogen fertilizer (June).
2.2. Yielding of the Apple Trees
2.3. Nutritional Status of Leaves and Fruits
2.4. Statistical Analysis
3. Results
3.1. Mineral Nitrogen Content in the Soil
3.2. Leaf Nutrient Status
3.3. Yielding of Apple Trees
3.4. Fruit Nutrient Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal nitrogen and water use in horticulture: Effects on quality and content of selected nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Dong, S.; Neilsen, D.; Neilsen, G.; Fuchigami, L.H. Foliar N application reduced soil NO3-N leaching loss in apple orchard. Plant Soil 2005, 268, 357–366. [Google Scholar] [CrossRef]
- Tagliavini, M.; Millard, P. Fluxes of nitrogen within deciduous fruit trees. Acta Sci. Pol. Hortorum Cultus 2005, 4, 21–30. [Google Scholar]
- Raese, J.T.; Drake, S.R.; Curry, E.A. Nitrogen fertilizer influences fruit quality, soil nutrition and cover crops, leaf colour and nitrogen content, biennial bearing and cold hardiness of ‘Golden Delicious’. J. Plant Nutr. 2007, 30, 1585–1604. [Google Scholar] [CrossRef]
- Bokszczanin, K.Ł.; Wrona, D.; Przybyłko, S. The effect of microbial inoculation under various nitrogen regimes on the uptake of nutrients by apple trees. Agronomy 2021, 11, 2348. [Google Scholar] [CrossRef]
- Marschner, P. Marschner's Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2005. [Google Scholar]
- Fernandes, M.S.; Rosiello, R.O.P. Mineral nitrogen in plant physiology and plant nutrition. Crit. Rev. Plant Sci. 1995, 14, 111–148. [Google Scholar] [CrossRef]
- Cruz-Alvarez, O.; Hernández-Rodríguez, O.A.; Jacobo-Cuellar, J.L.; Ávila-Quezada, G.; Morales-Maldonado, E.; Parra-Quezada, R.A.; Robles-Hernandez, L.; Ojeda-Barrios, D.L. Nitrogen fertilization in pecan and its effect on leaf nutrient concentration, yield and nut quality. Rev. Chapingo Ser. Hortic. 2020, 26, 163–173. [Google Scholar] [CrossRef]
- Lawlor, D.W. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot. 2002, 53, 773–787. [Google Scholar] [CrossRef]
- Tagliavini, M.; Quartieri, M.; Millard, P. Remobilised nitrogen and root uptake of nitrate for leaf growth, flowers and developing fruits of pear (P. communis) trees. Plant Soil 1997, 195, 137–142. [Google Scholar] [CrossRef]
- Menino, M.R.; Carranca, C.; de Varennes, A. Distribution and remobilization of nitrogen in young non-bearing orange trees grown under Mediterranean conditions. J. Plant Nutr. 2007, 30, 1083–1096. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Environmental Defense Fund. The Case for Reforming Farm Programs to Preserve the Environment and Help Family Farmers, Ranchers, and Foresters; Environmental Defense Fund: Washington, DC, USA, 2001. [Google Scholar]
- Elser, J.J.; Bennett, E. A broken biogeochemical cycle. Nat. Cell Biol. 2011, 478, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Tubiello, F.N.; Salvatore, M.; Ferrara, A.F.; House, J.; Federici, S.; Rossi, S.; Biancalani, R.; Golec, R.D.C.; Jacobs, H.; Flammini, A.; et al. The contribution of agriculture, forestry and other land use activities to global warming, 1990–2012. Glob. Change Biol. 2015, 21, 2655–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, S.E.; Tsigaridis, K.; Miller, R. Significant atmospheric aerosol pollution caused by world food cultivation. Geophys. Res. Lett. 2016, 43, 5394–5400. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Clark, M.; Williams, D.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nat. Cell Biol. 2017, 546, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kramer, S.B.; Reganold, J.P.; Glover, J.D.; Bohannan, B.J.M.; Mooney, H.A. Reduced nitrate leaching and enhanced denitrifier activity and efficiency in organically fertilized soils. Proc. Natl. Acad. Sci. USA 2006, 103, 4522–4527. [Google Scholar] [CrossRef] [Green Version]
- Mulvaney, R.L.; Khan, S.A.; Ellsworth, T.R. Synthetic nitrogen fertilizers deplete soil nitrogen: A global dilemma for sustainable cereal production. J. Environ. Qual. 2009, 38, 2295–2314. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.E.; Cambardella, C.A.; Laird, D.A.; Jaynes, D.B.; Meek, D.W. Nitrogen fertilizer effects on soil carbon balances in Midwestern U.S. agricultural systems. Ecol. Appl. 2009, 19, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Ernani, P.R.; Dias, J. Soil nitrogen application in the spring did not increase apple yield. Cienc. Rural 1999, 29, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Wrona, D. Effects on growth, cropping and fruit quality of ‘Šampion’ apple trees during 9 years of planting. Folia Hortic. 2004, 16, 55–60. [Google Scholar]
- Wrona, D. The influence of nitrogen fertilization on growth, yield and fruit size of ‘Jonagored’ apple trees. Acta Sci. Pol. Hortorum Cultus 2011, 10, 3–10. [Google Scholar]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Anal. Bioanal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Paltineanu, C.; Iancu, M.; Chatzoulakis, K.S. Orchard groundcover management effects on nitrate leaching in an irrigated heavy-clay soil. Acta Hort. 1997, 449, 739–745. [Google Scholar] [CrossRef]
- Thalheimer, M.; Paoli, N. Effects of time of nitrogen fertilization of apple trees on fruit quality and soil nitrogen content. Italus Hortus 2006, 13, 80–84. [Google Scholar]
- Jadczyszyn, T.; Kowalczyk, J.; Lipiński, W. Fertilization Recommendations for Field Crops and Permanent Grasslands; Institute of Soil Science; Plant Cultivation-State Research Institute: Puławy, Poland, 2010. [Google Scholar]
- Wrona, D.; Sadowski, A. Effects of nitrogen fertilisation and soil management on soil mineral nitrogen in the apple orchard. J. Fruit Ornam. Plant Res. 2004, 12, 191–199. [Google Scholar]
- Engel, A.; Blanke, M.; Lenz, F. Nutrient translocation from the grass alleyway to the tree strip with mulching in a fruit orchard. Erwerbs-Obstbau 2009, 51, 151–161. [Google Scholar] [CrossRef]
- Komosa, A. Changes in some chemical properties of the soil under grass sward and herbicide strips in apple orchard. Acta Hortic. 1990, 274, 223–230. [Google Scholar] [CrossRef]
- Deckers, T.; Schoofs, H.; Daemen, E.; Missotten, C. Effect of long term soil and leaf nitrogen applications to apple cv. Jonagold and Boskoop on N-min in the soil and on leaf and fruit quality. Acta Hortic. 2001, 564, 269–278. [Google Scholar] [CrossRef]
- Rupp, D. Nitrogen fertilization in apple orchards-relationships between available nitrogen in soil samples, nitrates in water and leaching of nitrogen. Acta Hortic. 1995, 383, 401–409. [Google Scholar] [CrossRef]
- Unuk, T.; Hribar, J.; Tojnko, S.; Simcic, M.; Pozrl, T.; Plestenjak, A.; Vidrih, R. Effect of nitrogen application and crop load on external and internal fruit quality parameters of apples. Dtsch. Lebensmitt. Rundsch. 2008, 104, 127–135. [Google Scholar]
- Tahir, I.I.; Svensson, S.-E.; Hansson, D. Floor management systems in an organic apple orchard affect fruit quality and storage life. HortScience 2015, 50, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Żelazny, W.R.; Licznar-Małańczuk, M. Soil quality and tree status in a twelve-year-old apple orchard under three mulch-based floor management systems. Soil Tillage Res. 2018, 180, 250–258. [Google Scholar] [CrossRef]
- Granatstein, D.; Sánchez, E. Research knowledge and needs for orchard floor management in organic tree fruit systems. Int. J. Fruit Sci. 2009, 9, 257–281. [Google Scholar] [CrossRef]
- Fallahi, E.; Fallahi, B.; Neilsen, G.H.; Neilsen, D.; Peryea, F.J. Effects of mineral nutrition on fruit quality and nutritional disorders in apples. Acta Hortic. 2010, 868, 49–60. [Google Scholar] [CrossRef]
- TerAvest, D.; Smith, J.L.; Boggs, L.C.; Hoagland, L.; Granatstein, D.; Reganold, J.P. Influence of orchard floor management and compost application timing on nitrogen partitioning in apple trees. HortScience 2010, 45, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Wrona, D.; Sadowski, A. Effects of nitrogen fertilisation in young apple orchard. Acta Hortic. 1997, 448, 481–486. [Google Scholar] [CrossRef]
- Marcelle, R.D. Mineral nutrition and fruit quality. Acta Hortic. 1995, 383, 219–226. [Google Scholar] [CrossRef]
- Fallahi, E.; Colt, W.M.; Baird, C.R.; Fallahi, B.; Chun, I.-J. Influence of nitrogen and bagging on fruit quality and mineral concentrations of ‘BC-2 Fuji’ apple. Horttechnology 2001, 13, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Pacholak, E.; Zachwieja, M.; Zydlik, Z. Effect of nitrogen fertilization on the content of mineral components in soil, leaves and fruits of ‘Šampion’ apple trees. Acta Sci. Pol. Hortorum Cultus 2004, 3, 207–2017. [Google Scholar]
- Raese, J.T.; Drake, S.R. Nitrogen fertilization and elemental composition affects fruit quality of 'Fuji' apples. J. Plant Nutr. 1997, 20, 1797–1809. [Google Scholar] [CrossRef]
- Kühn, B.F.; Bertelsen, M.; Sørensen, L. Optimising quality-parameters of apple cv. ‘Pigeon’ by adjustment of nitrogen. Sci. Hortic. 2011, 129, 369–375. [Google Scholar] [CrossRef]
- Ernani, P.R.; Rogeri, D.A.; Proença, M.M.; Dias, J. Addition of nitrogen had no effect on yield and quality of apples in an high density orchard carrying a dwarf rootstock. Rev. Bras. Frutic. 2008, 30, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Fallahi, E.; Mohan, K.S. Influence of nitrogen and rootstock on tree growth precocity, fruit quality, leaf mineral nutrients and fire blight in ‘Scarlet Gala’ apple. HortTechnology 2000, 10, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Rietra, R.P.J.J.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of nutrient antagonism and synergism on yield and fertilizer use efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef] [Green Version]
- Meheriuk, M.; Neilsen, G.M.; Hogue, E.J. The influence of nitrogen fertilization, season of application and orchard floor management on fruit quality and leaf mineral content of ‘Golden Delicious’ apple trees. Fruit Var. J. 1992, 46, 71–75. [Google Scholar]
- Rupp, D.; Hübner, H. Impact of nitrogen fertilization in apple orchards. Results of a long year fertilizing trial. Erwerbs-Obstbau 1995, 37, 29–31. [Google Scholar]
- Sotiropoulos, T.E.; Therios, I.N.; Dimassi, K.N.; Tsirakoglou, V. Effects of applications of a complex and Na-Ca fertilizer on leaf and fruit nutrient concentrations and some fruit quality parameter in two apple cultivars. Hortic. Sci. 2005, 43, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Cmelik, Z.; Tojnko, S.; Unuk, T. Fruit quality of ‘Fuji’ apple as affected by crop load and rates of nitrogen. Acta Hortic. 2006, 721, 147–152. [Google Scholar] [CrossRef]
- Fallahi, E.; Fallahi, B.; Retamales, J.B.; Valdes, C.; Tabatabei, S.J. Prediction of apple fruit quality using preharvest mineral nutrients. Acta Hortic. 2006, 721, 259–264. [Google Scholar] [CrossRef]
- Tahir, I.I.; Johansson, E.; Olsson, M.E. Improving the productivity, quality and storability of ‘Katja’ apple by better orchard management procedures. Hortic. Sci. 2008, 43, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Awad, M.A.; de Jager, A. Relationships between fruit nutrients and concentrations of flavonoids and chlorogenic acid in `Elstar' apple skin. Sci. Hortic. 2002, 92, 265–276. [Google Scholar] [CrossRef]
- Ludders, P. The effect of nitrogen nutrition on bitter pit in apples. Commun. Soil Sci. Plant Anal. 1979, 10, 401–415. [Google Scholar] [CrossRef]
- Perring, M. Effects of area of herbicide and grass sward treatments and amount of nitrogen fertilizer on the mineral composition during development. J. Hortic. Sci. 1984, 62, 17–25. [Google Scholar]
- Cheng, L.; Raba, R. Accumulation of macro- and micronutrients and nitrogen demand-supply relationship of ‘Gala’/‘Malling 26’ apple trees grown in sand culture. J. Am. Soc. Hortic. Sci. 2009, 134, 3–13. [Google Scholar] [CrossRef]
- Uysal, E. Influence of different nitrogen applications on mineral content of fruits in apple. Tarim Bilim. Derg. 2015, 8, 50–53. [Google Scholar]



{kind=link}
{kind=link}
| Year | Sampling Place | Sampling Time | Nitrogen Fertilization | Soil Layer [cm] | ||
|---|---|---|---|---|---|---|
| 0–30 | 30–60 | 60–90 | ||||
| 2011 | herbicyde strips | April | N-0 | 29.80 ± 0.70 | 12.32 ± 4.93 | 6.72 ± 2.98 |
| N-50 | 33.10 ± 6.14 | 24.03 ± 10.98 | 16.56 ± 9.19 | |||
| N-100 | 26.46 ± 4.53 | 24.06 ± 8.04 | 33.76 ± 13.16 | |||
| N-100G | 41.62 ± 2.89 | 24.13 ± 7.53 | 14.78 ± 9.19 | |||
| May | N-0 | 26.73 ± 5.64 | 20.83 ± 5.12 | 11.86 ± 5.50 | ||
| N-50 | 66.97 ± 15.24 | 26.56 ± 8.00 | 19.96 ± 7.80 | |||
| N-100 | 55.87 ± 10.90 | 49.97 ± 18.36 | 35.90 ± 10.24 | |||
| N-100G | 30.24 ± 5.84 | 25.27 ± 7.58 | 13.74 ± 12.33 | |||
| June | N-0 | 22.97 ± 8.73 | 23.80 ± 2.04 | 19.70 ± 5.54 | ||
| N-50 | 35.73 ± 2.09 | 33.93 ± 7.50 | 31.40 ± 15.99 | |||
| N-100 | 67.93 ± 16.69 | 70.47 ± 24.06 | 72.93 ± 16.90 | |||
| N-100G | 28.63 ± 7.52 | 42.20 ± 28.54 | 31.50 ± 15.39 | |||
| grass alleyways | April | N-0 | 30.46 ± 12.05 | 17.23 ± 4.40 | 7.11 ± 3.02 | |
| N-50 | 34.77 ± 5.07 | 17.40 ± 5.16 | 14.29 ± 8.66 | |||
| N-100 | 38.80 ± 3.42 | 20.20 ± 1.91 | 15.50 ± 4.61 | |||
| N-100G | 23.00 ± 9.86 | 14.43 ± 0.61 | 11.95 ± 3.86 | |||
| May | N-0 | 34.80 ± 12.45 | 22.57 ± 11.16 | 11.59 ± 5.72 | ||
| N-50 | 26.33 ± 6.15 | 24.93 ± 5.13 | 19.97 ± 4.67 | |||
| N-100 | 79.30 ± 14.26 | 33.03 ± 12.55 | 21.47 ± 9.23 | |||
| N-100G | 71.93 ± 16.08 | 35.80 ± 6.50 | 43.30 ± 13.88 | |||
| June | N-0 | 22.33 ± 7.31 | 22.27 ± 11.98 | 16.43 ± 4.78 | ||
| N-50 | 25.87 ± 8.56 | 18.96 ± 2.15 | 14.80 ± 6.82 | |||
| N-100 | 63.23 ± 39.69 | 34.43 ± 2.99 | 37.03 ± 23.70 | |||
| N-100G | 32.77 ± 6.23 | 25.63 ± 7.11 | 14.66 ± 4.55 | |||
| 2012 | herbicyde strips | April | N-0 | 35.80 ± 6.93 | 17.63 ± 8.31 | 10.49 ± 2.84 |
| N-50 | 48.40 ± 9.17 | 28.30 ± 13.81 | 16.58 ± 6.32 | |||
| N-100 | 56.00 ± 17.15 | 94.40 ± 7.03 | 69.03 ± 24.18 | |||
| N-100G | 39.87 ± 10.92 | 37.80 ± 5.12 | 27.17 ± 8.77 | |||
| May | N-0 | 38.50 ± 3.62 | 26.66 ± 6.77 | 20.63 ± 11.91 | ||
| N-50 | 99.67 ± 4.93 | 38.03 ± 22.26 | 15.53 ± 0.40 | |||
| N-100 | 98.90 ± 22.67 | 66.20 ± 16.00 | 57.10 ± 17.23 | |||
| N-100G | 38.56 ± 12.26 | 29.26 ± 0.80 | 9.85 ± 2.59 | |||
| June | N-0 | 34.43 ± 16.00 | 49.63 ± 31.26 | 45.93 ± 4.37 | ||
| N-50 | 53.93 ± 37.70 | 18.27 ± 8.60 | 18.37 ± 9.66 | |||
| N-100 | 160.70 ± 59.88 | 105.97 ± 8.59 | 90.33 ± 10.03 | |||
| N-100G | 65.80 ± 24.11 | 26.73 ± 6.32 | 11.53 ± 5.23 | |||
| grass alleyways | April | N-0 | 26.00 ± 9.72 | 11.56 ± 3.11 | 8.68 ± 1.58 | |
| N-50 | 47.47 ± 13.42 | 21.40 ± 1.13 | 21.17 ± 13.96 | |||
| N-100 | 30.93 ± 9.01 | 25.67 ± 11.57 | 12.29 ± 6.30 | |||
| N-100G | 28.33 ± 11.35 | 19.46 ± 7.46 | 20.45 ± 7.57 | |||
| May | N-0 | 32.30 ± 10.14 | 14.06 ± 9.09 | 10.90 ± 4.62 | ||
| N-50 | 36.40 ± 7.76 | 19.23 ± 3.95 | 8.46 ± 1.99 | |||
| N-100 | 67.20 ± 0.00 | 44.50 ± 26.44 | 26.60 ± 20.90 | |||
| N-100G | 79.23 ± 2.56 | 39.96 ± 25.13 | 19.67 ± 5.61 | |||
| June | N-0 | 52.17 ± 26.55 | 21.53 ± 13.01 | 25.53 ± 22.68 | ||
| N-50 | 55.47 ± 26.32 | 23.63 ± 3.21 | 22.97 ± 15.47 | |||
| N-100 | 74.87 ± 18.50 | 35.00 ± 0.10 | 7.91 ± 3.84 | |||
| N-100G | 21.07 ± 6.73 | 29.47 ± 22.76 | 15.00 ± 3.83 | |||
| Year | p = 0.002 | p = 0.039 | p = 0.475 | |||
| Sampling place | p = 0.095 | p < 0.0001 | p < 0.0001 | |||
| Time of the sampling | p = 0.003 | p = 0.047 | p = 0.020 | |||
| Nitrogen fertilization | p < 0.0001 | p < 0.0001 | p < 0.0001 | |||
| Year × sampling place | p = 0.003 | p = 0.071 | p = 0.14 | |||
| Year × time of the sampling | p = 0.14 | p = 0.058 | p = 0.44 | |||
| Year × nitrogen fertilization | p = 0.298 | p = 0.024 | p = 0.277 | |||
| Sampling place × time of the sampling | p = 0.544 | p = 0.254 | p = 0.061 | |||
| Sampling place × nitrogen fertilization | p = 0.12 | p < 0.0001 | p < 0.0001 | |||
| Time of the sampling × nitrogen fertilization | p = 0.001 | p = 0.556 | p = 0.250 | |||
| Year × sampling place × time of sampling | p = 0.828 | p = 0.601 | p = 0.870 | |||
| Year × sampling place × nitrogen fertilization | p = 0.049 | p = 0.020 | p = 0.013 | |||
| Year × sampling place × nitrogen fertilization × time of sampling | p = 0.176 | p = 0.065 | p = 0.10 | |||
| Year | Sampling Place | Sampling Time | Soil Layer [cm] | Nitrogen Fertilization | |||
|---|---|---|---|---|---|---|---|
| N-0 | N-50 | N-100 | N-100G | ||||
| 2011 | herbicyde strips | April | 0–30 | 29.80 ± 0.70 | 33.10 ± 6.14 | 26.46 ± 4.53 | 41.62 ± 2.89 |
| 30–60 | 12.32 ± 4.93 | 24.03 ± 10.98 | 24.06 ± 8.04 | 24.13 ± 7.53 | |||
| 60–90 | 6.72 ± 2.98 | 16.56 ± 9.19 | 33.76 ± 13.16 | 14.78 ± 9.19 | |||
| May | 0–30 | 26.73 ± 5.64 | 66.97 ± 15.24 | 55.87 ± 10.90 | 30.24 ± 5.84 | ||
| 30–60 | 20.83 ± 5.12 | 26.56 ± 8.00 | 49.97 ± 18.36 | 25.27 ± 7.58 | |||
| 60–90 | 11.86 ± 5.50 | 19.96 ± 7.80 | 35.90 ± 10.24 | 13.74 ± 12.33 | |||
| June | 0–30 | 22.97 ± 8.73 | 35.73 ± 2.09 | 67.93 ± 16.69 | 28.63 ± 7.52 | ||
| 30–60 | 23.80 ± 2.04 | 33.93 ± 7.50 | 70.47 ± 24.06 | 42.20 ± 28.54 | |||
| 60–90 | 19.70 ± 5.54 | 31.40 ± 15.99 | 72.93 ± 16.90 | 31.50 ± 15.39 | |||
| grass alleyways | April | 0–30 | 30.46 ± 12.05 | 34.77 ± 5.07 | 38.80 ± 3.42 | 23.00 ± 9.86 | |
| 30–60 | 17.23 ± 4.40 | 17.40 ± 5.16 | 20.20 ± 1.91 | 14.43 ± 0.61 | |||
| 60–90 | 7.11 ± 3.02 | 14.29 ± 8.66 | 15.50 ± 4.61 | 11.95 ± 3.86 | |||
| May | 0–30 | 34.80 ± 12.45 | 26.33 ± 6.15 | 79.30 ± 14.26 | 71.93 ± 16.08 | ||
| 30–60 | 22.57 ± 11.16 | 24.93 ± 5.13 | 33.03 ± 12.55 | 35.80 ± 6.50 | |||
| 60–90 | 11.59 ± 5.72 | 19.97 ± 4.67 | 21.47 ± 9.23 | 43.30 ± 13.88 | |||
| June | 0–30 | 22.33 ± 7.31 | 25.87 ± 8.56 | 63.23 ± 39.69 | 32.77 ± 6.23 | ||
| 30–60 | 22.27 ± 11.98 | 18.96 ± 2.15 | 34.43 ± 2.99 | 25.63 ± 7.11 | |||
| 60–90 | 16.43 ± 4.78 | 14.80 ± 6.82 | 37.03 ± 23.70 | 14.66 ± 4.55 | |||
| 2012 | herbicyde strips | April | 0–30 | 35.80 ± 6.93 | 48.40 ± 9.17 | 56.00 ± 17.15 | 39.87 ± 10.92 |
| 30–60 | 17.63 ± 8.31 | 28.30 ± 13.81 | 94.40 ± 7.03 | 37.80 ± 5.12 | |||
| 60–90 | 10.49 ± 2.84 | 16.58 ± 6.32 | 69.03 ± 24.18 | 27.17 ± 8.77 | |||
| May | 0–30 | 38.50 ± 3.62 | 99.67 ± 4.93 | 98.90 ± 22.67 | 38.56 ± 12.26 | ||
| 30–60 | 26.66 ± 6.77 | 38.03 ± 22.26 | 66.20 ± 16.00 | 29.26 ± 0.80 | |||
| 60–90 | 20.63 ± 11.91 | 15.53 ± 0.40 | 57.10 ± 17.23 | 9.85 ± 2.59 | |||
| June | 0–30 | 34.43 ± 16.00 | 53.93 ± 37.70 | 160.70 ± 59.88 | 65.80 ± 24.11 | ||
| 30–60 | 49.63 ± 31.26 | 18.27 ± 8.60 | 105.97 ± 8.59 | 26.73 ± 6.32 | |||
| 60–90 | 45.93 ± 4.37 | 18.37 ± 9.66 | 90.33 ± 10.03 | 11.53 ± 5.23 | |||
| grass alleyways | April | 0–30 | 26.00 ± 9.72 | 47.47 ± 13.42 | 30.93 ± 9.01 | 28.33 ± 11.35 | |
| 30–60 | 11.56 ± 3.11 | 21.40 ± 1.13 | 25.67 ± 11.57 | 19.46 ± 7.46 | |||
| 60–90 | 8.68 ± 1.58 | 21.17 ± 13.96 | 12.29 ± 6.30 | 20.45 ± 7.57 | |||
| May | 0–30 | 32.30 ± 10.14 | 36.40 ± 7.76 | 67.20 ± 0.00 | 79.23 ± 2.56 | ||
| 30–60 | 14.06 ± 9.09 | 19.23 ± 3.95 | 44.50 ± 26.44 | 39.96 ± 25.13 | |||
| 60–90 | 10.90 ± 4.62 | 8.46 ± 1.99 | 26.60 ± 20.90 | 19.67 ± 5.61 | |||
| June | 0–30 | 52.17 ± 26.55 | 55.47 ± 26.32 | 74.87 ± 18.50 | 21.07 ± 6.73 | ||
| 30–60 | 21.53 ± 13.01 | 23.63 ± 3.21 | 35.00 ± 0.10 | 29.47 ± 22.76 | |||
| 60–90 | 25.53 ± 22.68 | 22.97 ± 15.47 | 7.91 ± 3.84 | 15.00 ± 3.83 | |||
| Year | p = 0.006 | p = 0.18 | p = 0.004 | p = 0.719 | |||
| Sampling place | p = 0.192 | p = 0.013 | p < 0.0001 | p = 0.881 | |||
| Time of the sampling | p = 0.002 | p = 0.395 | p = 0.001 | p = 0.052 | |||
| Soil layer | p < 0.0001 | p < 0.0001 | p = 0.002 | p < 0.0001 | |||
| Year × sampling place | p = 0.079 | p = 0.876 | p < 0.0001 | p = 0.484 | |||
| Year × time of the sampling | p = 0.018 | p = 0.979 | p = 0.851 | p = 0.639 | |||
| Year × soil layer | p = 0.652 | p = 0.005 | p = 0.425 | p = 0.358 | |||
| Sampling place × time of the sampling | p = 0.018 | p = 0.979 | p = 0.851 | p = 0.639 | |||
| Sampling place × soil layer | p = 0.322 | p = 0.079 | p = 0.259 | p = 0.598 | |||
| Time of the sampling × soil layer | p = 0.156 | p = 0.183 | p = 0.185 | p = 0.206 | |||
| Year × sampling place × time of sampling | p = 0.907 | p = 0.196 | p = 0.424 | p = 0.941 | |||
| Year × sampling place × soil layer | p = 0.437 | p = 0.680 | p = 0.524 | p = 0.430 | |||
| Year × sampling place × soil layer × time of sampling | p = 0.271 | p = 0.996 | p = 0.871 | p = 0.0006 | |||
| Macroelement | Year | Nitrogen Fertilization | |||
|---|---|---|---|---|---|
| N-0 | N-50 | N-100 | N-100G | ||
| N | 2010 | a 1.91 ± 0.11 A | a 2.11 ± 0.07 B | a 2.15 ± 0.10 B | a 2.11 ± 0.03 B |
| 2011 | a 1.92 ± 0.29 A | b 2.32 ± 0.09 B | b 2.37 ± 0.08 B | a 2.28 ± 0.11 B | |
| 2012 | a 1.81 ± 0.07 A | ab 2.23 ± 0.06 B | ab 2.26 ± 0.07 B | a 2.21 ± 0.08 B | |
| P | 2010 | a 0.29 ± 0.001 B | a 0.16 ± 0.01 A | a 0.15 ± 0.002 A | a 0.17 ± 0.01 A |
| 2011 | a 0.30 ± 0.07 B | a 0.18 ± 0.01 A | a 0.16 ± 0.01 A | a 0.17 ± 0.01 A | |
| 2012 | a 0.29 ± 0.03 B | a 0.17 ± 0.01 A | a 0.16 ± 0.002 A | a 0.17 ± 0.008 A | |
| K | 2010 | a 1.54 ± 0.04 B | b 1.22 ± 0.10 A | b 1.31 ± 0.11 A | a 1.34 ± 0.18 AB |
| 2011 | a 1.56 ± 0.17 B | a 1.06 ± 0.08 A | a 1.24 ± 0.03 A | a 1.14 ± 0.17 A | |
| 2012 | a 1.55 ± 0.09 B | ab 1.15 ± 0.06 A | ab 1.28 ± 0.07 A | a 1.24 ± 0.17 A | |
| Mg | 2010 | a 0.18 ± 0.01 A | a 0.27 ± 0.01 B | a 0.24 ± 0.02 B | a 0.24 ± 0.03 B |
| 2011 | b 0.25 ± 0.02 A | b 0.30 ± 0.02 B | b 0.29 ± 0.02 B | b 0.30 ± 0.03 B | |
| 2012 | ab 0.22 ± 0.02 A | ab 0.28 ± 0.01 B | ab 0.27 ± 0.01 B | ab 0.27 ± 0.02 B | |
| Ca | 2010 | a 2.14 ± 0.04 A | a 2.14 ± 0.03 A | a 2.14 ± 0.06 A | a 2.12 ± 0.03 A |
| 2011 | a 2.18 ± 0.05 A | a 2.15 ± 0.05 A | a 2.13 ± 0.04 A | a 2.16 ± 0.06 A | |
| 2012 | a 2.17 ± 0.02 A | a 2.14 ± 0.01 A | a 2.15 ± 0.01 A | a 2.15 ± 0.02 A | |
| Year | Nitrogen Fertilization | |||
|---|---|---|---|---|
| N-0 | N-50 | N-100 | N-100G | |
| 2010 | a 9.52 ± 2.61 A | a 12.01 ± 1.62 A | a 13.25 ± 2.23 A | a 11.31 ± 1.41 A |
| 2011 | ab 16.77 ± 6.97 A | ab 20.13 ± 8.48 A | b 18.27 ± 3.13 A | b 18.21 ± 6.51 A |
| 2012 | b 24.60 ± 5.62 A | b28.25 ± 3.90 A | c 30.79 ± 2.84 A | c 28.46 ± 3.22 A |
| Macroelement | Year | Nitrogen Fertilization | |||
|---|---|---|---|---|---|
| N-0 | N-50 | N-100 | N-100G | ||
| N | 2010 | a 0.27 ± 0.044 A | a 0.32 ± 0.024 BC | a 0.33 ± 0.0095 C | a 0.28 ± 0.011 AB |
| 2011 | a 0.21 ± 0.069 A | a 0.32 ± 0.032 B | a 0.34 ± 0.024 B | a 0.28 ± 0.017 AB | |
| 2012 | a 0.24 ± 0.055 A | a 0.32 ± 0.029 B | a 0.33 ± 0.011 B | a 0.32 ± 0.010 B | |
| P | 2010 | b 0.10 ± 0.0047 A | a 0.094 ± 0.0067 A | b 0.096 ± 0.0084 A | b 0.095 ± 0.0046 A |
| 2011 | a 0.086 ± 0.0043 A | a 0.085 ± 0.0065 A | a 0.080 ± 0.0026 A | a 0.083 ± 0.0064 A | |
| 2012 | b 0.098 ± 0.0029 B | a 0.089 ± 0.0061 AB | ab 0.089 ± 0.0042 A | ab 0.090 ± 0.054 AB | |
| K | 2010 | a 0.78 ± 0.071 A | a 0.68 ± 0.047 A | a 0.71 ± 0.066 A | a 0.67 ± 0.016 A |
| 2011 | a 0.74 ± 0.031 A | b 0.75 ± 0.011 A | a 0.69 ± 0.016 A | a 0.72 ± 0.075 A | |
| 2012 | a 0.76 ± 0.022 B | ab 0.72 ± 0.019 AB | a 0.70 ± 0.039 A | a 0.69 ± 0.035 A | |
| Mg | 2010 | a 0.033 ± 0.0022 A | a 0.032 ± 0.0019 A | a 0.036 ± 0.0079 A | a 0.031 ± 0.0013 A |
| 2011 | a 0.032 ± 0.0030 A | a 0.035 ± 0.00033 A | a 0.033 ± 0.002 A | a 0.034 ± 0.0021 A | |
| 2012 | a 0.033 ± 0.0043 A | a 0.032 ± 0.00016 A | a 0.035 ± 0.0064 A | a 0.032 ± 0.0011 A | |
| Ca | 2010 | a 0.043 ± 0.0012 A | a 0.036 ± 0.0067 A | a 0.037 ± 0.0024 A | a 0.035 ± 0.0065 A |
| 2011 | a 0.035 ± 0.0074 A | a 0.031 ± 0.0037 A | a 0.032 ± 0.0059 A | a 0.032 ± 0.0087 A | |
| 2012 | a 0.041 ± 0.0035 A | a 0.034 ± 0.0010 A | a 0.036 ± 0.0029 A | a 0.033 ± 0.0042 A | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, W.; Wrona, D.; Przybyłko, S. Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits. Agriculture 2022, 12, 2169. https://doi.org/10.3390/agriculture12122169
Kowalczyk W, Wrona D, Przybyłko S. Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits. Agriculture. 2022; 12(12):2169. https://doi.org/10.3390/agriculture12122169
Chicago/Turabian StyleKowalczyk, Wojciech, Dariusz Wrona, and Sebastian Przybyłko. 2022. "Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits" Agriculture 12, no. 12: 2169. https://doi.org/10.3390/agriculture12122169