
Consequences of the Long-Term Fertilization System Use on Physical and Microbiological Soil Status in the Western Polissia of Ukraine
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area and the Characteristics of Experimental Plots
- Soil without fertilization (C);
- O1FS—organic fertilization system: soil + manure;
- MFS—mineral fertilization system: soil + NPK;
- O1MFS—organic-mineral fertilization system: soil + manure + NPK;
- O2MFS—organic-mineral fertilization system: soil + siderate + NPK.
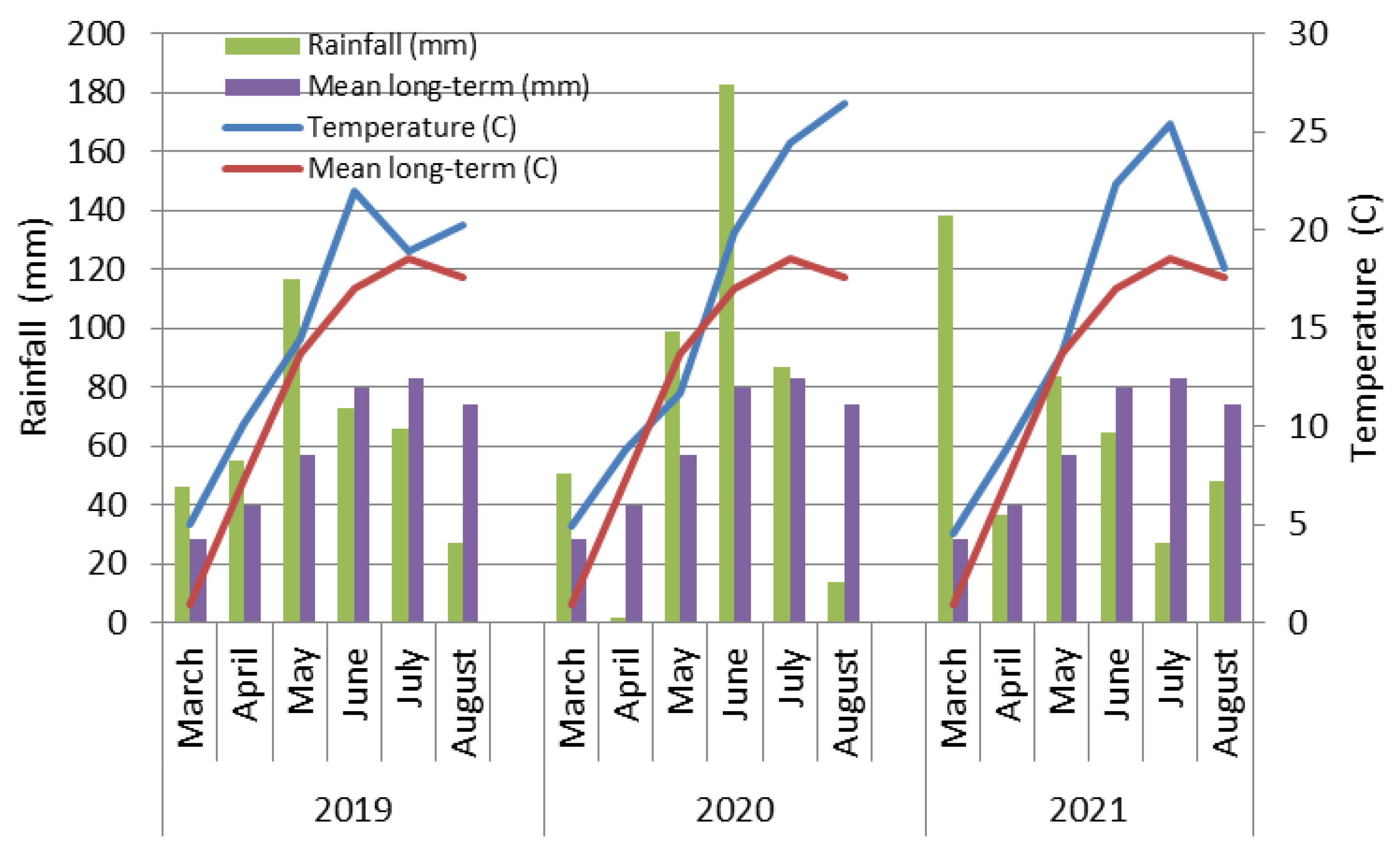
2.2. Meteorological Conditions
2.3. Soil Sampling
2.4. Chemical Analyses
2.5. Microbiological Analyses
2.6. Statistical Analysis
3. Results and Discussion
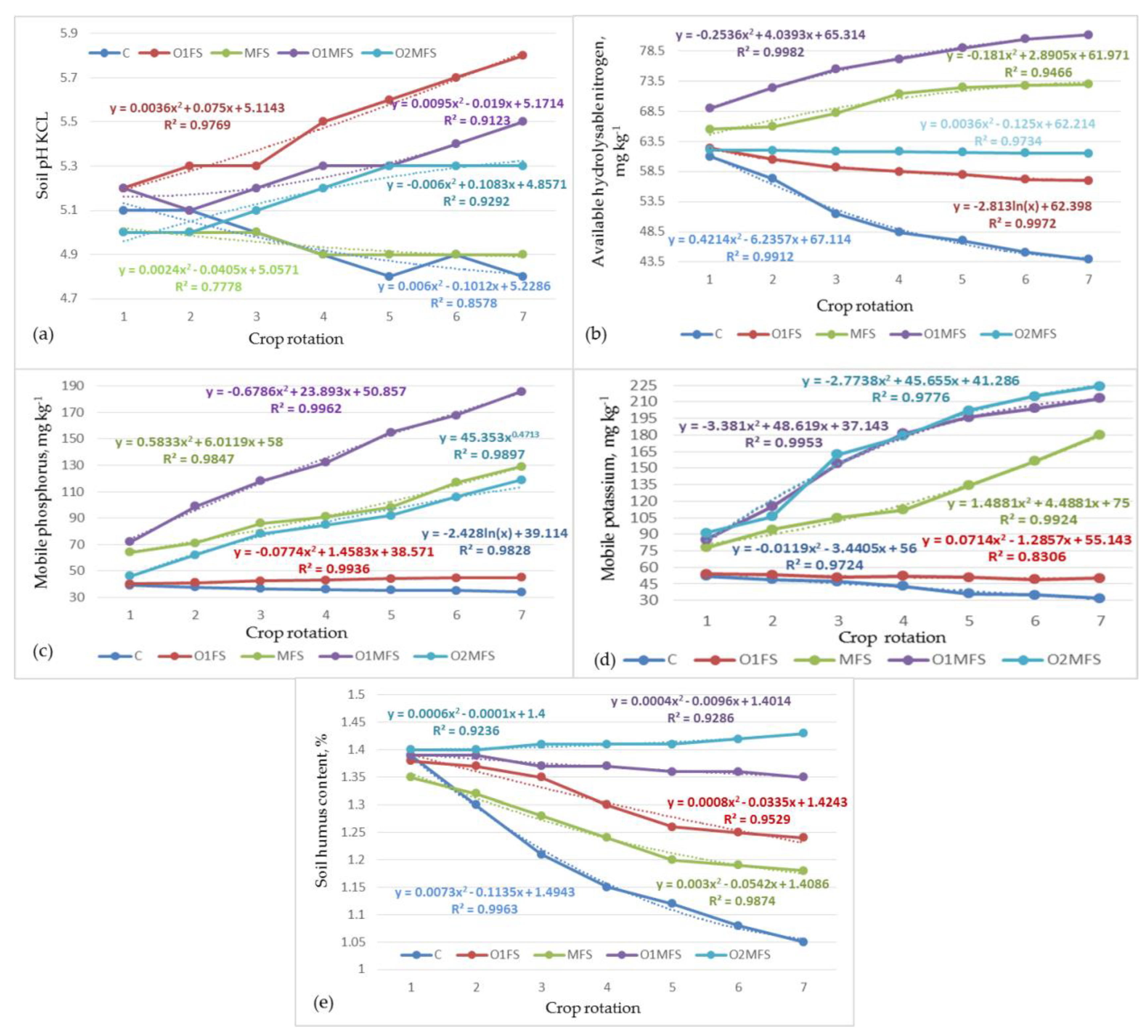
3.1. Soil Physical and Chemical Properties after Long-Term Fertilization Systems
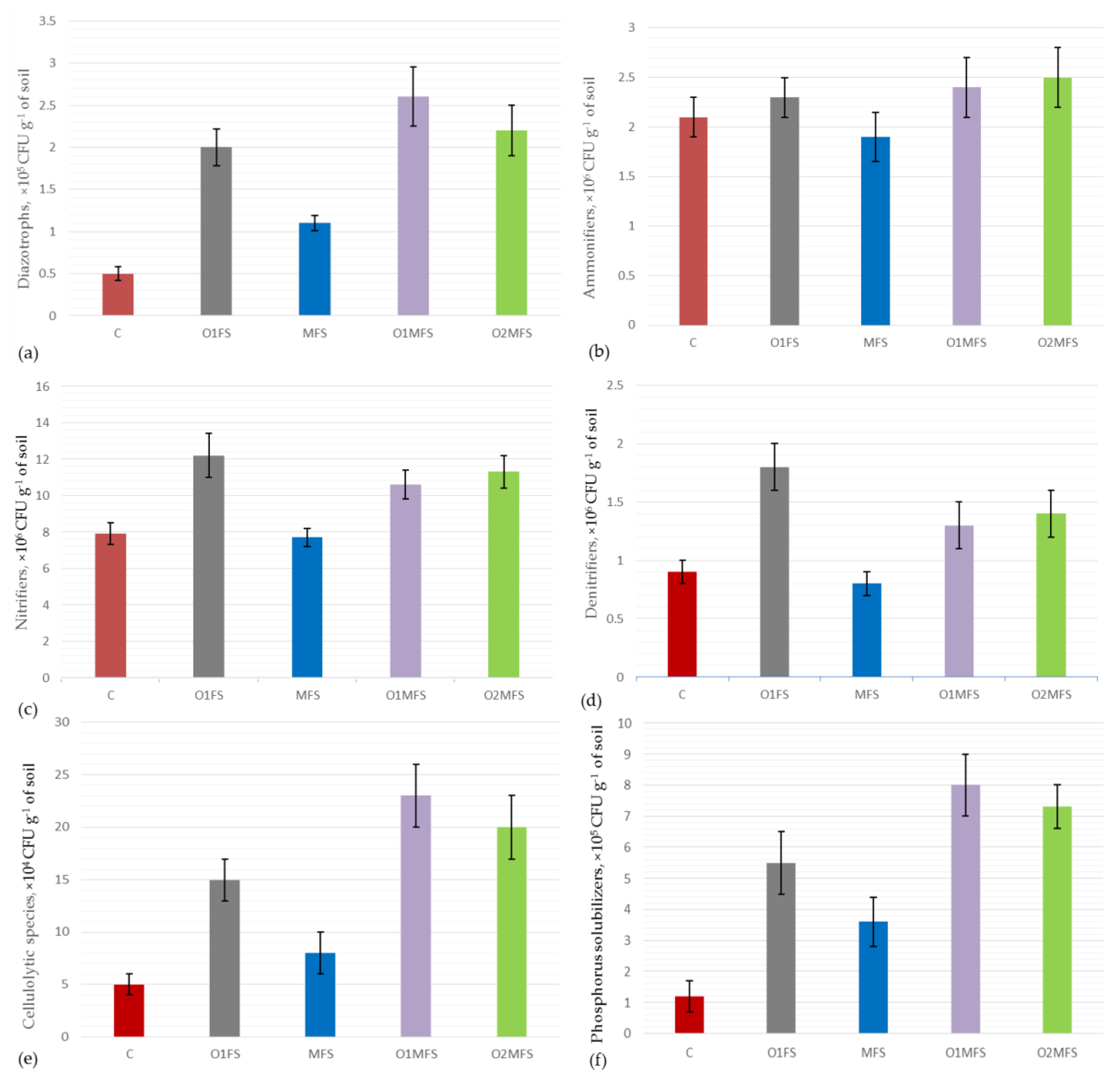
3.2. The Number of Microorganisms of the Main Taxonomic Groups
3.3. The Number of Different Physiological Groups of Soil Microorganisms
3.3.1. Atmospheric Nitrogen-Fixing Bacteria (Diazotrophs)
3.3.2. Ammonifiers (Organotrophic Bacteria)
3.3.3. Nitrifiers (Mineral Nitrogen Assimilating Bacteria)
3.3.4. Denitrifiers
3.3.5. Cellulolytic Microorganisms
3.3.6. Phosphorus-Solubilizing Microorganisms
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Trasar-Cepeda, C.; Sotres, F.G.; Bello, D. Use of enzyme activities to monitor pollution of agricultural land. EQA-Int. J. Environ. Qual. 2016, 22, 15–24. [Google Scholar] [CrossRef]
- Sabir, M.S.; Shahzadi, F.; Ali, F.; Shakeela, Q.; Niaz, Z.; Ahmed, S. Comparative Effect of Fertilization Practices on Soil Microbial Diversity and Activity: An Overview. Curr. Microbiol. 2021, 78, 3644–3655. [Google Scholar] [CrossRef] [PubMed]
- Joniec, J.; Kwiatkowska, E.; Kwiatkowski, C.A. Assessment of the Effects of Soil Fertilization with Spent Mushroom Substrate in the Context of Microbial Nitrogen Transformations and the Potential Risk of Exacerbating the Greenhouse Effect. Agriculture 2022, 12, 1190. [Google Scholar] [CrossRef]
- Poulton, P.R.; Johnston, A.E. Can Long-Term Experiments Help Us Understand, and Manage, the Wider Landscape—Examples from Rothamsted, England. Exploring and Optimizing Agricultural Landscapes; Innovations in Landscape Research; Mueller, L., Sychev, V.G., Dronin, N.M., Eulenstein, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 233–252. [Google Scholar] [CrossRef]
- Van der Bom, F.; Nunes, I.; Raymond, N.S.; Hansen, V.; Bonnichsen, L.; Magid, J.; Nybroe, O.; Jensen, L.S. Long-term fertilisation form, level and duration affect the diversity, structure and functioning of soil microbial communities in the field. Soil Biol. Biochem. 2018, 122, 91–103. [Google Scholar] [CrossRef]
- Li, C.-X.; Ma, S.-C.; Shao, Y.; Ma, S.-T.; Zhang, L.-L. Effects of long-term organic fertilization on soil microbiologic characteristics, yield and sustainable production of winter wheat. J. Integr. Agric. 2018, 17, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Kibblewhite, M.G.; Ritz, K.; Swift, M.J. Soil health in agricultural systems. Phil. Trans. R. Soc. 2008, 363, 685–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Delgado-Baquerizo, M.; Zhu, Y.; Han, X.; Han, X.; Xin, X.; Li, W.; Guo, Z.; Dang, T.; Li, C. Responses of Soil Bacterial Diversity to Fertilization are Driven by Local Environmental Context Across China. Engineering 2022, 12, 164–170. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Miller, R.M.; Jastrow, J.D. Mycorrhizal Fungi Influence Soil Structure. In Arbuscular Mycorrhizas: Physiology and Function; Kapulnik, Y., Douds, D.D., Eds.; Springer: Dordrecht, Germany, 2000. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Megyes, M.; Borsodi, A.K.; Árendás, T.; Márialigeti, K. Variations in the diversity of soil bacterial and archaeal communities in response to different long-term fertilization regimes in maize fields. Appl. Soil Ecol. 2021, 168, 104120. [Google Scholar] [CrossRef]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility. A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Ninh, H.T.; Grandy, A.S.; Wickings, K.; Snapp, S.S.; Kirk, W.; Hao, J. Organic amendment effects on potato productivity and quality are related to soil microbial activity. Plant Soil 2015, 386, 223–236. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Chen, R.; Hu, J.; Zhao, F.; Wang, J.; Chu, H.; Zhang, J.; Dolfing, J.; Lin, X. Bacillus asahii comes to the fore in organic manure fertilized alkaline soil. Soil Biol. Biochem. 2015, 81, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Ye, G.; Luo, J.; Di, H.J.; Lindsey, S.; Fan, J.; Liu, D.; Ding, W. Long-term organic fertilization regulates the abundance of major nitrogen-cycling-related genes in aggregates from an acidic Ultisol. Appl. Soil Ecol. 2021, 165, 104014. [Google Scholar] [CrossRef]
- Li, X.; Su, Y.; Ahmed, T.; Ren, H.; Javed, M.R.; Yao, Y.; An, Q.; Yan, J.; Li, B. Effects of Different Organic Fertilizers on Improving Soil from Newly Reclaimed Land to Crop Soil. Agriculture 2021, 11, 560. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, H.; He, X.; Thomas, B.W.; Lupwayi, N.Z.; Hao, X.; Thomas, M.C.; Shi, X. Fertilization Shapes Bacterial Community Structure by Alteration of Soil pH. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Bardgett, R.D.; Vitousek, P.M.; Maestre, F.T.; Williams, M.A.; Eldridge, D.J.; Lambers, H.; Neuhauser, S.; Gallardo, A.; García-Velázquez, L.; et al. Changes in belowground biodiversity during ecosystem development. Proc. Natl. Acad. Sci. USA 2019, 116, 6891–6896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Wall, D.H.; Nielsen, U.N.; Six, J. Soil biodiversity and human health. Nature. 2015, 528, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Leff, W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayer, J.; Cassman, K.G. Agricultural innovation to protect the environment. Proc. Natl. Acad. Sci. USA 2013, 110, 8345–8348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Zhalnina, K.; Dias, R.; de Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH determines microbial diversity and composition in the park grass experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.M.; Kim, M.; Tateno, R.; Kim, W.; Wang, J.J.; Lai-Hoe, A.; Ab Shukor, N.A.; Rahim, R.A.; Go, R.; Adams, J.M. Soil pH and biome are both key determinants of soil archaeal community structure. Soil Biol. Biochem. 2015, 88, 1–8. [Google Scholar] [CrossRef]
- Tripathi, B.M.; Stegen, J.C.; Kim, M.; Dong, K.; Adams, J.M.; Lee, Y.K. Soil pH mediates the balance between stochastic and deterministic assembly of bacteria. ISME J. 2018, 12, 1072–1083. [Google Scholar] [CrossRef]
- Tripathi, B.M.; Kim, M.; Singh, D.; Lee-Cruz, L.; Lai-Hoe, A.; Ainuddin, A.N.; Go, R.; Rahim, R.A.; Husni, M.H.A.; Chun, J. Tropical soil bacterial communities in Malaysia: pH dominates in the equatorial tropics too. Microb. Ecol. 2012, 64, 474–484. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Lupatini, M.; Korthals, G.W.; de Hollander, M.; Janssens, T.K.; Kuramae, E.E. Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, C.; Tu, C.; Hoyt, G.D.; DeForest, J.L.; Hu, S. Long-term no-tillage and organic input management enhanced the diversity and stability of soil microbial community. Sci. Total Environ. 2017, 609, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.-C.; Schlaeppi, K. Farming practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Harkes, P.; Suleiman, A.K.A.; van den Elsen, S.J.J.; de Haan, J.J.; Holterman, M.; Kuramae, E.E.; Helder, J. Conventional and organic soil management as divergent drivers of resident and active fractions of major soil food web constituents. Nature Res. Sci. Rep. 2019, 9, 13521. [Google Scholar] [CrossRef] [Green Version]
- Peltoniemi, K.; Velmala, S.; Fritze, H.; Lemola, R.; Pennanen, T. Long-term impacts of organic and conventional farming on the soil microbiome in boreal arable soil. Eur. J. Soil Sci. 2021, 104, 103314. [Google Scholar] [CrossRef]
- Ndubuisi-Nnaji, U.U.; Adegoke, A.A.; Ogbu, H.I.; Ezenobi, N.O.; Okoh, A.I. Effect of long-term organic fertilizer application on soil microbial dynamics. Afr. J. Biotechnol. 2011, 10, 556–559. [Google Scholar] [CrossRef]
- Ye, G.; Banerjee, S.; He, I.-Z.; Fan, J.; Wang, Z.; Wei, X.; Hu, H.-W.; Zheng, Y.; Duan, C.; Wan, S.; et al. Manure application increases microbiome complexity in soil aggregate fractions: Results of an 18-year field experiment. Agric. Ecosyst. Environ. 2021, 307, 91–103. [Google Scholar] [CrossRef]
- Gautam, A.; Sekaran, U.; Guzman, J.; Kovács, P.; Hernandez, J.L.G.; Kumar, S. Responses of soil microbial community structure and enzymatic activities to long-term application of mineral fertilizer and beef manure. Environ. Sustain. Indic. 2020, 8, 100073. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Schwenke, G.D.; Van Zwieten, L. Impact of agricultural inputs on soil organisms—A review. Aust. J. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.G.; Spaner, D. Cropping Systems Management, Soil Microbial Communities, and Soil Biological Fertility. In Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming; Lichtfouse, E., Ed.; Springer: Heidelberg, Germany, 2010; pp. 217–242. [Google Scholar] [CrossRef]
- Smith, P.; Fallon, P.D.; Smith, J.U.; Powlson, D.S. Soil Organic Matter Network (SOMNET): 2001 Model and Experimental Metadata, 2nd ed.; GCTE Report No. 7. A Core Project of the IGBP; GCTE: Wallingford, UK, 2001; p. 115. [Google Scholar]
- World Reference Base for Soil Resources 2014. Update 2015; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; pp. 171–172.
- Zviahyntsev, D.H. Methods of Soil Microbiology and Biochemistry, 2nd ed.; Moscow State University: Moscow, Russia, 1991; p. 303. (In Russian) [Google Scholar]
- Feder, F. Effects of Fertilisation Using Organic Waste Products with Mineral Complementation on Sugarcane Yields and Soil Properties in a 4 Year Field Experiment. Agriculture 2021, 11, 985. [Google Scholar] [CrossRef]
- Radulov, I.; Berbecea, A.; Sala, F.; Crista, F.; Lato, A. Mineral fertilization influence on soil pH, cationic exchange capacity and nutrient content. Res. J. Agric. Sci. 2011, 43, 160–165. [Google Scholar]
- Wen, Y.C.; Li, H.Y.; Lin, Z.A.; Zhao, B.Q.; Sun, Z.B.; Yuan, L.; Xu, J.K.; Li, Y.Q. Long-term fertilization alters soil properties and fungal community composition in fluvo-aquic soil of the North China Plain. Sci. Rep. 2020, 10, 7198. [Google Scholar] [CrossRef]
- Schroder, J.L.; Zhang, H.; Girma, K.; Raun, W.R.; Penn, C.J.; Payton, M.E. Soil Acidification from Long-Term Use of Nitrogen Fertilizers on Winter Wheat. Soil Sci. Soc. Am. J. 2011, 75, 957–964. [Google Scholar] [CrossRef]
- Zhao, B.-Q.; Li, X.-Y.; Li, X.-P.; Shi, X.-J.; Huang, S.-M.; Wang, B.-R.; Zhu, P.; Yang, X.-Y.; Liu, H.; Chen, Y.; et al. Long-Term Fertilizer Experiment Network in China: Crop Yields and Soil Nutrient Trends. Agron J. 2010, 102, 216–230. [Google Scholar] [CrossRef]
- Hati, K.M.; Swarup, A.; Dwivedi, A.K.; Misra, A.K.; Bandyopadhyay, K.K. Changes in soil physical properties and organic carbon status at the topsoil horizon of a vertisol of central India after 28 years of continuous cropping, fertilization and manuring. Agr. Ecosyst. Environ. 2007, 119, 127–134. [Google Scholar] [CrossRef]
- Neina, D. The Role of Soil pH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K.; Barbosa Filho, M.B. Influence of pH on Productivity, Nutrient Use Efficiency by Dry Bean, and Soil Phosphorus Availability in a No-Tillage System. Commun. Soil Sci. Plant Anal. 2008, 39, 1016–1025. [Google Scholar] [CrossRef]
- Wang, R.; Zou, R.; Liu, J.; Liu, L.; Hu, Y. Spatial Distribution of Soil Nutrients in Farmland in a Hilly Region of the Pearl River Delta in China Based on Geostatistics and the Inverse Distance Weighting Method. Agriculture 2021, 11, 50. [Google Scholar] [CrossRef]
- Paradelo, R.; Eden, M.; Martínez, I.; Keller, T.; Houot, S. Soil physical properties of a Luvisol developed on loess after 15 years of amendment with compost. Soil Till. Res. 2019, 191, 207–215. [Google Scholar] [CrossRef]
- Janzen, H.H. Beyond carbon sequestration: Soil as conduit of solar energy. Eur. J. Soil Sci. 2015, 66, 19–32. [Google Scholar] [CrossRef]
- Wang, X.; Yan, J.; Zhang, X.; Zhang, S.; Chen, Y. Organic manure input improves soil water and nutrients use for sustainable maize (Zea mays. L) productivity on the Loess Plateau. PLoS ONE 2020, 15, e0238042. [Google Scholar] [CrossRef]
- Oueriemmi, H.; Kidd, P.S.; Trasar-Cepeda, C.; Rodríguez-Garrido, B.; Zoghlami, R.I.; Ardhaoui, K.; Prieto-Fernández, Á.; Moussa, M. Evaluation of Composted Organic Wastes and Farmyard Manure for Improving Fertility of Poor Sandy Soils in Arid Regions. Agriculture 2021, 11, 415. [Google Scholar] [CrossRef]
- Parham, J.; Deng, S.; Raun, W.R.; Johnson, G. Long-term cattle manure application in soil. Biol. Fertil. Soils 2002, 35, 328–337. [Google Scholar] [CrossRef]
- Obriot, F.; Stauffer, M.; Goubard, Y.; Cheviron, N.; Peres, G.; Eden, M.; Revallier, A.; Vieuble Gonod, L.; Houot, S. Multi-criteria indices to evaluate the effects of repeated organic amendment applications on soil and crop quality. Agric. Ecosyst. Environ. 2016, 232, 165–178. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, C.; Du, L.; Chen, X.; Li, J. Effects of long-term localized fertilization on soil humus combining form in sheltered vegetable field. Chin. J. Appl. Ecol. 2006, 17, 831–834, In Chinese. [Google Scholar] [PubMed]
- Poulton, P.; Johnston, J.; Macdonald, A.; White, R.; Powlson, D. Major limitations to achieving “4 per 1000” increases in soil organic carbon stock in temperate regions: Evidence from long-term experiments at Rothamsted Research, United Kingdom. Glob. Change Biol. 2018, 24, 2563–2584. [Google Scholar] [CrossRef] [Green Version]
- Powlson, D.S.; Bhogal, A.; Chambers, B.J.; Coleman, K.; Macdonald, A.J.; Goulding, W.T.; Whitmore, A.P. The potential to increase soil carbon stocks through reduced tillage or organic material additions in England and Wales: A case study. Agric. Ecosyst. Environ. 2012, 146, 23–33. [Google Scholar] [CrossRef]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Aislabie, J.; Deslippe, J. Soil Microbes and Their Contribution to Soil Services. In Ecosystems Services in New Zealand Conditions and Trends; Dymond, J.R., Ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2013; pp. 143–161. [Google Scholar]
- Langley, J.A.; Hungate, B.A. Plant community feedbacks and long-term ecosystem responses to multi-factored global change. AoB Plants 2014, 6, 24–27. [Google Scholar] [CrossRef]
- Demyanyuk, O.; Shatsman, D.; Symochko, L. Structure and Dynamics of Soil Microbial Communities of Natural and Transformed Ecosystems. Environ. Res. Eng. Manag. 2020, 76, 97–105. [Google Scholar] [CrossRef]
- Lammel, D.R.; Barth, G.; Ovaskainen, O.; Cruz, L.M.; Zanatta, J.A.; Ryo, M.; Souza, E.M.; Pedrosa, F.O. Direct and indirect effects of a pH gradient bring insights into the mechanisms driving prokaryotic community structures. Microbiome 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Li, Y.; Yao, H. Fertilization with inorganic and organic nutrients changes diazotroph community composition and N-fixation rates. J. Soils Sediments 2018, 18, 1076–1086. [Google Scholar] [CrossRef]
- Sivojiene, D.; Kacergius, A.; Baksiene, E.; Maseviciene, A.; Zickiene, L. The Influence of Organic Fertilizers on the Abundance of Soil Microorganism Communities, Agrochemical Indicators, and Yield in East Lithuanian Light Soils. Plants 2021, 10, 2648. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Lazcano, C.; Zhu-Barker, X.; Decock, C. Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review. Microorganisms 2021, 9, 983. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Fan, F.; Song, A.; Cui, P.; Li, T.; Liang, Y. Denitrification potential under different fertilization regimes is closely coupled with changes in the denitrifying community in a black soil. Appl. Microbiol. Biotechnol. 2015, 99, 5719–5729. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Wierzchowski, P.S.; Stępień, W.; Górska, E.B. The Reaction of Cellulolytic and Potentially Cellulolytic Spore-Forming Bacteria to Various Types of Crop Management and Farmyard Manure Fertilization in Bulk Soil. Agronomy 2021, 11, 772. [Google Scholar] [CrossRef]
- Tang, H.; Li, C.; Xu, Y.; Cheng, K.; Shi, L.; Wen, L.; Li, W.; Xiao, X. Effects of fertilizer practice on fungal and actinobacterial cellulolytic community with different humified particle-size fractions in double-cropping field. Sci. Rep. 2021, 11, 18441. [Google Scholar] [CrossRef]
- Samaddar, S.; Chatterjee, P.; Truu, J.; Anandham, R.; Kim, S.; Sa, T. Long-term phosphorus limitation changes the bacterial community structure and functioning in paddy soils. Appl. Soil Ecol. 2019, 134, 111–115. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Azziz, G.; Bajsa, N.; Haghjou, T.; Taulé, C.; Valverde, A.; Igual, J.; Arias, A. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop–pasture rotations in a no-tillage regime in Uruguay. Appl. Soil Ecol. 2012, 61, 320–326. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Unit | Value |
|---|---|---|
| Exchange acidity, pH KCL | 1 mol KCl | 5.1 |
| Hydrolytic acidity | meq 100 g | 2.34 |
| Cation exchange capacity, S | meq 100 g | 2.6 |
| Degree of bases saturation, V | % | 52.6 |
| Humus content | % | 1.39 |
| Easily hydrolyzed nitrogen | mg kg−1 | 61 |
| Mobile phosphorus | mg kg−1 | 39 |
| Exchangeable potassium | mg kg−1 | 52 |
| Copper, Cu | mg kg−1 | 2.05 |
| Zinc, Zn | mg kg−1 | 11.7 |
| Boron, B | mg kg−1 | 0.08 |
| Cobalt, Co | mg kg−1 | 1.39 |
| Nickel, Ni | mg kg−1 | 5.9 |
| Lead, Pb | mg kg−1 | 4.7 |
| Cadmium, Cd | mg kg−1 | 0.06 |
| Manganese, Mn | mg kg−1 | 98.0 |
| Medium | Media Composition for 1000 mL H2O |
|---|---|
| Meat-peptone agar medium (MPA) | Dry fermented peptone—10.0 g; meat extract—11.0 g; NaCl—5.0 g; agar—15.0 g. |
| Starch-ammonia agar medium (CAA) | KH2PO4—1.0 g; (NH4)2SO4—2.0 g; MgSO4—1.0 g; NaCl—1.0 g; CaCO3—3.0 g; soluble starch—10.0 g; agar—20.0 g |
| Chapek’s medium | KH2PO4—1.0 g; MgSO4—0.5 g; NaNO3—3.0 g; KCl—0.5 g; sucrose—30 g; FeSO4—0.01 g; agar—20 g. Before pouring into Petri dishes, streptomycin was added to a flask with a hot medium. |
| Giltaya’s medium | Two solutions were prepared, which were later combined. Solution 1: KNO3—2.1 g; asparagine—1.0 g; distilled water—250 mL. Solution 2: sodium citric acid—5.0 g; KH2PO4—2.0 g; MgSO4—2.0 g; CaCl2—2.0 g; FeCl3—traces; distilled water—500 mL. |
| Hutchinson’s medium | K2HPO4—1.0 g; CaCl2—0.1 g; MgSO4—0.3 g; NaCl—0.1 g; FeCl3—0.01 g; NaNO3—2.50 g; CaCO3—10 g; agar—20 g. After solidification of the medium in Petri dishes, before sowing, sterile filter paper was placed on the surface of the medium as a source of cellulose. |
| Muromtsev’s medium | Glucose—10.0 g; asparagine—1.0 g; K2SO4—0.2 g; MgSO4 • 7H2O—0.4 g; yeast autolysate—0.5 g; agar—17.0 g; KH2PO4—1.0 g; MgSO4—0.5 g; NaNO3—3.0 g; KCl—0.5 g; sucrose—30 g; FeSO4—0.01 g; agar—20 g. Ca3(PO4)2 was added before pouring the medium into Petri dishes. |
| Vinohradsky’s liquid medium | K2HPO4—0.5 g; MgSO4—0.5 g; glucose—15 g; NaCl—traces; FeSO4—traces; MnSO4—traces. Before pouring the medium, CaCO3 was added to each tube at the tip of the spatula. |
| Experiment Variants | Bacteria, ×106 CFU g−1 of Soil | Micromycetes, ×103 CFU g−1 of Soil |
|---|---|---|
| C | 11.0 ± 0.2 | 44 ± 1.2 |
| O1FS | 14.5 ± 0.3 | 24 ± 0.5 |
| MFS | 9.6 ± 0.3 | 38 ± 1.1 |
| O1MFS | 13.0 ± 0.4 | 32 ± 0.8 |
| O2MFS | 13.8 ± 0.3 | 28 ± 0.5 |
| Taxonomic Groups | Regression Equation y = ax ± c | Coefficients of | |
|---|---|---|---|
| Correlation, R | Determination, R2 | ||
| pH KCl | |||
| Bacteria | y = 1.04x + 9.26 | 0.88 | 0.65 |
| Micromycetes | y = −4.6x + 47 | −0.91 | 0.84 |
| Humus, % | |||
| Bacteria | y = 1.04x + 9.26 | 0.75 | 0.65 |
| Micromycetes | y = −3.8x + 44.6 | −0.74 | 0.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puzniak, O.; Hrynchyshyn, N.; Datsko, T.; Andruszczak, S.; Hulko, B. Consequences of the Long-Term Fertilization System Use on Physical and Microbiological Soil Status in the Western Polissia of Ukraine. Agriculture 2022, 12, 1955. https://doi.org/10.3390/agriculture12111955
Puzniak O, Hrynchyshyn N, Datsko T, Andruszczak S, Hulko B. Consequences of the Long-Term Fertilization System Use on Physical and Microbiological Soil Status in the Western Polissia of Ukraine. Agriculture. 2022; 12(11):1955. https://doi.org/10.3390/agriculture12111955
Chicago/Turabian StylePuzniak, Oksana, Natalia Hrynchyshyn, Tetiana Datsko, Sylwia Andruszczak, and Bohdan Hulko. 2022. "Consequences of the Long-Term Fertilization System Use on Physical and Microbiological Soil Status in the Western Polissia of Ukraine" Agriculture 12, no. 11: 1955. https://doi.org/10.3390/agriculture12111955