
Control of Apple Replant Disease Using Mixed Cropping with Brassica juncea or Allium fistulosum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Treatment
2.2. Fungal Pathogen Isolation, Identification and Pathogenicity Testing
2.3. DNA Extraction and PCR Amplification
2.4. Terminal-Restriction Fragment Length Polymorphism (T-RFLP) Analysis
2.5. Sequence Analysis of Clone Libraries
2.6. Quantification of F. proliferatum in Replanted Soil by Real-Time Quantitative PCR (qPCR)
2.7. Statistical Analysis
3. Results
3.1. Effect of Mixed Cropping with A. fistulosum and B. juncea on the Growth of Replanted Seedlings
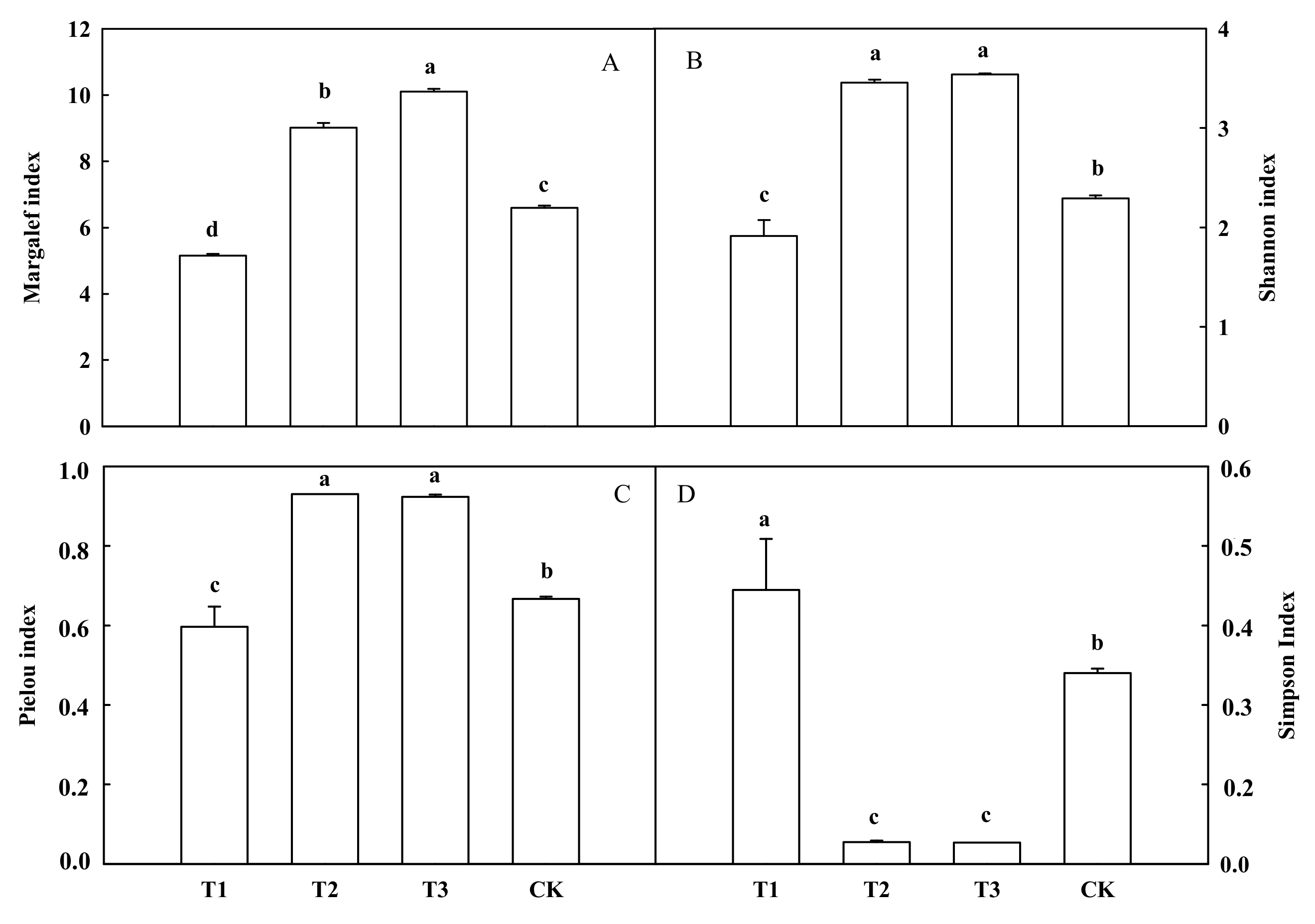
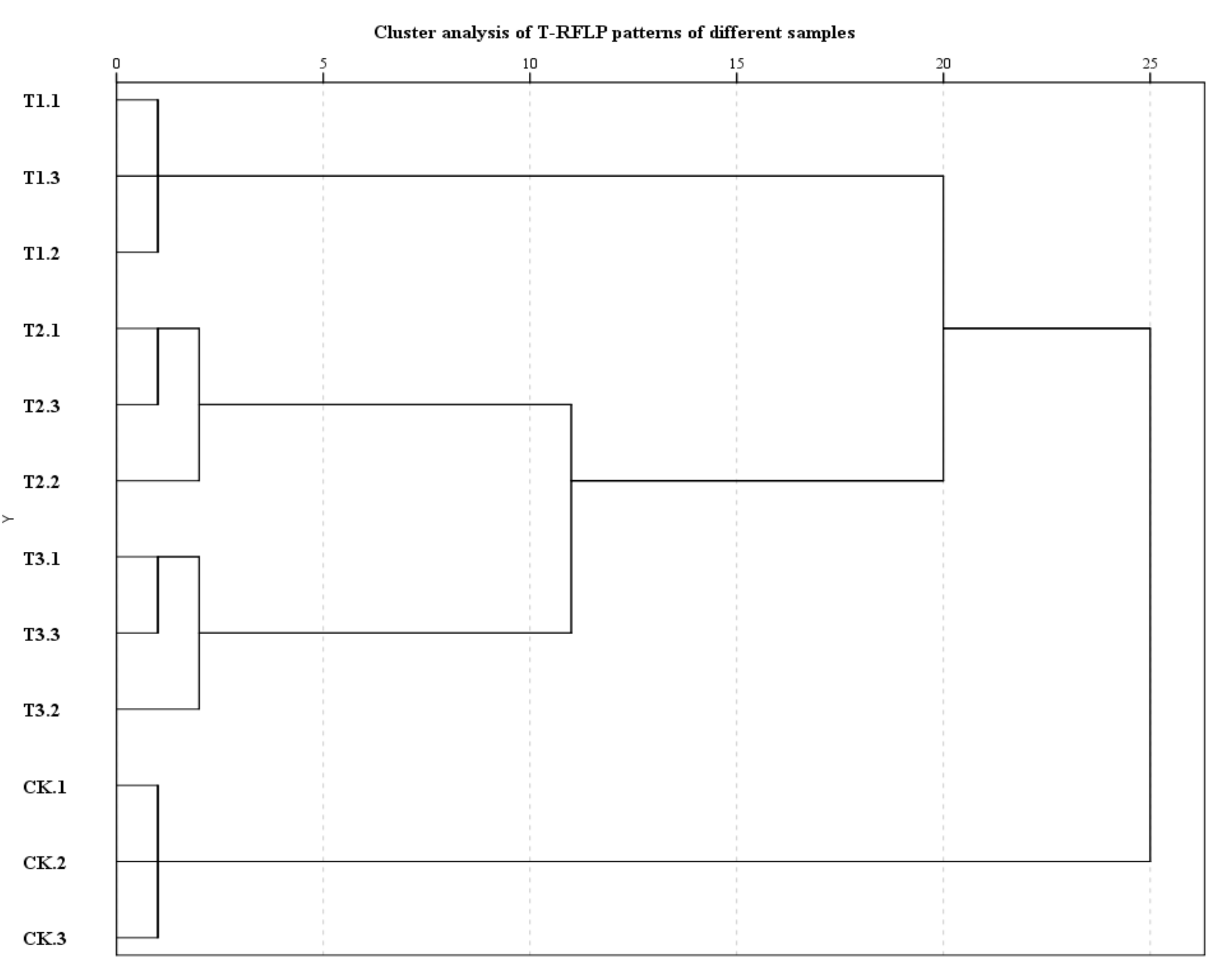
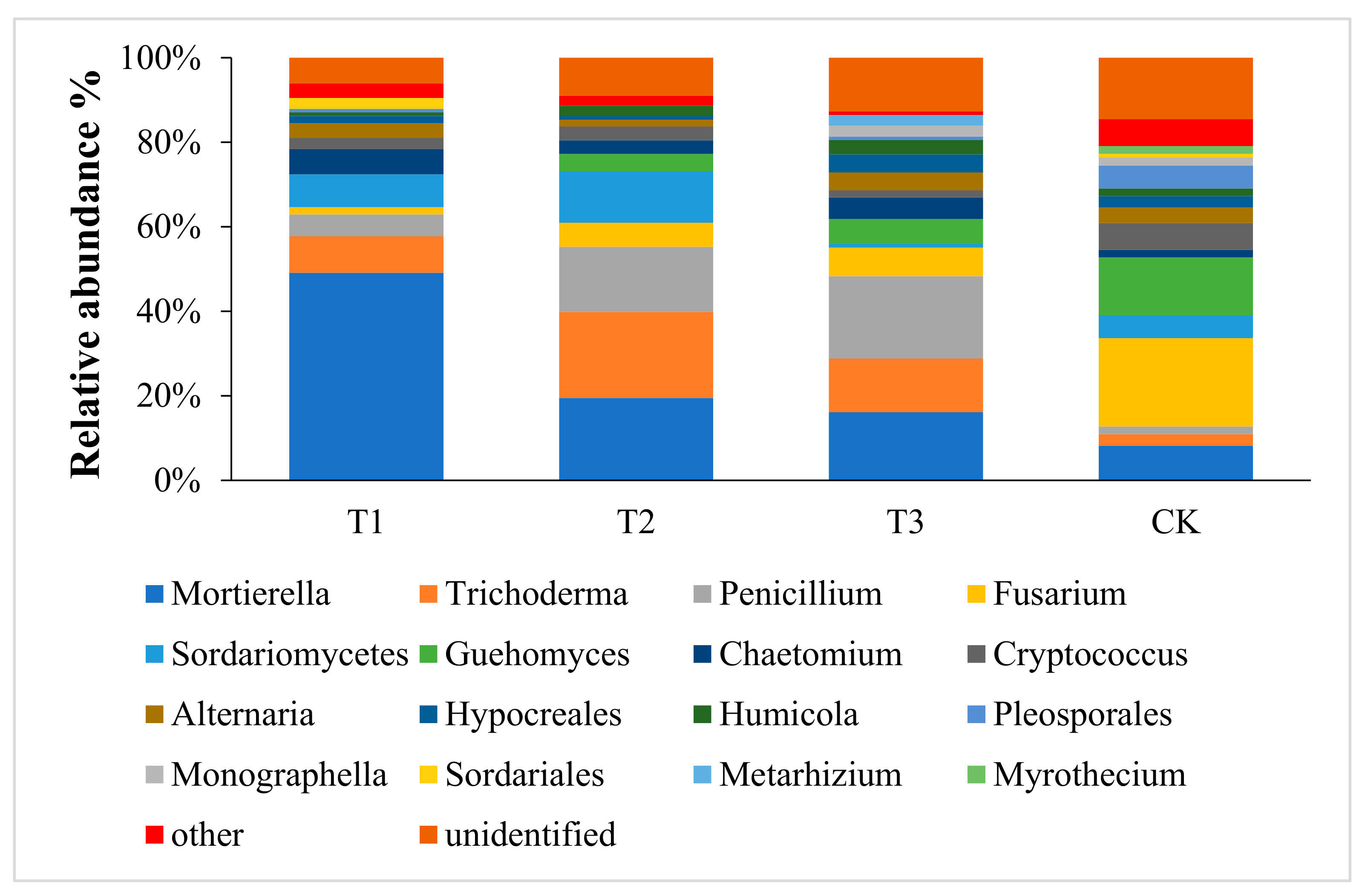
3.2. Effects of Mixed Cropping with A. fistulosum and B. juncea on the Fungal Community
3.3. Pathogenicity Testing of F. proliferatum and Real-Time Quantitative PCR (qPCR) Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mazzola, M.; Manici, L.M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Laurent, A.S.; Merwin, I.A.; Thies, J.E. Long-term orchard groundcover management systems affect soil microbial communities and apple replant disease severity. Plant Soil 2008, 304, 209–225. [Google Scholar] [CrossRef]
- Stirling, G.R.; Dullahide, S.R.; Nikulin, A. Management of lesion nematode (Pratylenchus jordanensis) on replanted apple trees. Anim. Prod. Sci 1995, 35, 247–258. [Google Scholar] [CrossRef]
- Zhang, H.; Mallik, A.; Zeng, R.S. Control of Panama disease of banana by rotating and intercropping with Chinese chive (Allium tuberosum Rottler): Role of plant volatiles. J. Chem. Ecol. 2013, 39, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Manici, L.M.; Kelderer, M.; Frankewhittle, I.H.; Rühmer, T.; Baab, G.; Nicoletti, F.; Caputo, F.; Topp, A.; Insam, H.; Naef, A. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe. Appl. Soil Ecol. 2013, 72, 207–214. [Google Scholar] [CrossRef]
- Yim, B.; Smalla, K.; Winkelmann, T. Evaluation of apple replant problems based on different soil disinfection treatments—Links to soil microbial community structure? Plant Soil 2013, 366, 617–631. [Google Scholar] [CrossRef]
- Weerakoon, D.M.N.; Reardon, C.L.; Paulitz, T.C.; Izzo, A.D.; Mazzola, M. Long-term suppression of Pythium abappressorium induced by Brassica juncea seed meal amendment is biologically mediated. Soil Biol. Biochem. 2012, 51, 44–52. [Google Scholar] [CrossRef]
- Manici, L.M.; Ciavatta, C.; Kelderer, M.; Erschbaumer, G. Replant problems in South Tyrol: Role of fungal pathogens and microbial population in conventional and organic apple orchards. Plant Soil 2003, 256, 315–324. [Google Scholar] [CrossRef]
- Mazzola, M. Elucidation of the microbial complex having a causal role in the development of apple replant disease in Washington. Phytopathology 1998, 88, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke-Whittle, I.H.; Manici, L.M.; Insam, H.; Stres, B. Rhizosphere bacteria and fungi associated with plant growth in soils of three replanted apple orchards. Plant Soil 2015, 395, 317–333. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Botha, W.J.; Spes, C.F.; Mcleod, A. Characterization of fungi (Fusarium and Rhizoctonia) and oomycetes (Phytophthora and Pythium) associated with apple orchards in South Africa. Eur. J. Plant Pathol. 2011, 130, 215–229. [Google Scholar] [CrossRef]
- Manici, L.M.; Kelderer, M.; Caputo, F.; Mazzola, M. Auxin-mediated relationships between apple plants and root inhabiting fungi: Impact on root pathogens and potentialities of growth-promoting populations. Plant Pathol. 2015, 64, 843–851. [Google Scholar] [CrossRef]
- Atucha, A.; Merwin, I.A.; Brown, M.G. Long-term effects of four groundcover management systems in an apple orchard. Hortscience 2011, 46, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Schoor, L.V.; Denman, S.; Cook, N.C. Characterisation of apple replant disease under South African conditions and potential biological management strategies. Sci. Hortic. 2009, 119, 153–162. [Google Scholar] [CrossRef]
- Rodríguez-Kábana, R.; Canullo, G.H. Cropping systems for the management of phytonematodes. Phytoparasitica 1992, 20, 211–224. [Google Scholar] [CrossRef]
- Mazzola, M.; Zhao, X.W. Brassica juncea seed meal particle size influences chemistry but not soil biology-based suppression of individual agents inciting apple replant disease. Plant Soil 2010, 337, 313–324. [Google Scholar] [CrossRef]
- Nelson, M.K.; Eric, B. Compost-induced suppression of Pythium damping-off is mediated by fatty-acid-metabolizing seed-colonizing microbial communities. Appl. Environ. Microbiol. 2003, 69, 452–460. [Google Scholar]
- Scheuerell, S.J.; Sullivan, D.M.; Mahaffee, W.F. Suppression of seedling damping-off caused by Pythium ultimum, P. irregulare, and Rhizoctonia solani in container media amended with a diverse range of pacific northwest compost sources. Phytopathology 2005, 95, 306–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, N.; Gowen, S.R.; El-Hassan, S.A.; Inam-Ul-Haq, M.; Shahina, F.; Pembroke, B. Efficacy of neem (Azadirachta indica) formulations on biology of root-knot nematodes (Meloidogyne javanica) on tomato. Crop Prot. 2008, 27, 36–43. [Google Scholar] [CrossRef]
- Natarajan, N.; Cork, A.; Boomathi, N.; Pandi, R.; Velavan, S.; Dhakshnamoorthy, G. Cold aqueous extracts of African marigold, Tagetes erecta for control tomato root knot nematode, Meloidogyne incognita. Crop Prot. 2006, 25, 1210–1213. [Google Scholar] [CrossRef]
- Angus, J.F.; Gardner, P.A.; Kirkegaard, J.A.; Desmarchelier, J.M. Biofumigation: Isothiocyanates released from Brassica roots inhibit growth of the take-all fungus. Plant Soil 1994, 162, 107–112. [Google Scholar] [CrossRef]
- Lewis, J.A.; Papavizas, G.C. Effect of sulfur-containing volatile compounds and vapors from cabbage decomposition on Aphanomyces euteiches. Phytopathology 1971, 61, 208–214. [Google Scholar] [CrossRef]
- Smolinska, U.; Morra, M.J.; Knudsen, G.R.; James, R.L. Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum. Plant Dis. 2003, 87, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Mau, J.L.; Chen, C.P.; Hsieh, P.C. Antimicrobial effect of extracts from Chinese Chive, Cinnamon, and Corni Fructus. J. Agric. Food Chem. 2001, 49, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Potential of Chinese chive oil as a natural antimicrobial for controlling Flavobacterium columnare infection in Nile tilapia Oreochromis niloticus. Fish Sci. 2009, 75, 1431–1437. [Google Scholar] [CrossRef]
- Seo, K.I.; Moon, Y.H.; Sang, U.C.; Park, K.H. Antibacterial Activity of S-Methyl Methanethiosulfinate and S-Methyl 2-Propene-1-thiosulfinate from Chinese Chive toward Escherichia coli O157:H7. Biosci. Biotech. Biochem. 2001, 65, 966–968. [Google Scholar] [CrossRef]
- Arnault, I.; Fleurance, C.; Vey, F.; Fretay, G.D.; Auger, J. Use of Alliaceae residues to control soil-borne pathogens. Ind. Crops Prod. 2013, 49, 265–272. [Google Scholar] [CrossRef]
- Mazzola, M. Transformation of soil microbial community structure and rhizoctonia-suppressive potential in response to apple roots. Phytopathology 1999, 89, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Buchan, A.; Newell, S.Y.; Butler, M.; Biers, E.J.; Hollibaugh, J.T.; Moran, M.A. Dynamics of bacterial and fungal communities on decaying salt marsh grass. Appl. Environ. Microbiol. 2003, 69, 6676–6687. [Google Scholar] [CrossRef] [Green Version]
- Ngala, B.M.; Woods, S.R.; Back, M.A. Sinigrin degradation and G. pallida suppression in soil cultivated with brassicas under controlled environmental conditions. Appl. Soil Ecol. 2015, 95, 9–14. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, J.; Liu, S.Z.; Wei, Q.P. Manure refinement affects apple rhizosphere bacterial community structure: A study in sandy soil. PLoS ONE 2013, 8, e76937. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Cheng, Z.; Meng, H.; Liu, L.; Li, H.; Dong, Y. Intercropping of Green Garlic (Allium sativum L.) Induces Nutrient Concentration Changes in the Soil and Plants in Continuously Cropped Cucumber (Cucumis sativus L.) in a Plastic Tunnel. PLoS ONE 2013, 8, e62173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.P.; Yin, D.H.; Chen, S.J.; Xia, F.; Yang, J.; Li, Q.; Wang, W. Effect of biocontrol agent Pseudomonas fluorescens 2P24 on soil fungal community in cucumber rhizosphere using T-RFLP and DGGE. PLoS ONE 2012, 7, e31806. [Google Scholar]
- Wang, Y.; Fu, F.; Li, J.; Wang, G.; Wu, M.; Zhan, J.; Chen, X.; Mao, Z. Effects of seaweed fertilizer on the growth of Malus hupehensis rehd. seedlings, soil enzyme activities and fungal communities under replant condition. Eur. J. Soil Biol. 2016, 75, 1–7. [Google Scholar] [CrossRef]
- Ghanbarzadeh, B.; Safaie, N.; Goltapeh, E.M. Antagonistic activity and hyphal interactions of Trichoderma spp. against Fusarium proliferatum and F. oxysporum in vitro. Arch. Phytopathol. Pflanzenschutz 2014, 47, 1979–1987. [Google Scholar] [CrossRef]
- Sharma, P. Complexity of trichoderma-fusarium interaction and manifestation of biological control. Aust. J. Crop. Sci. 2011, 5, 1027–1038. [Google Scholar]
- Zhang, X.F.; Xiang, L.; Wang, Y.F.; Wang, G.S.; Liu, H.X.; Sun, G.W.; Shen, X.; Chen, X.S.; Zhou, H.; Yin, C.M. Identification of Penicillium oxalicum A1 strain and antagonistic effects on four species of Fusarium pathogen of apple. Acta Hort. 2016, 43, 841–852. [Google Scholar]
- Li, R.; Shen, Z.Z.; Sun, L.; Zhang, R.F.; Fu, L.; Deng, X.H.; Shen, Q.R. Novel soil fumigation method for suppressing cucumber Fusarium wilt disease associated with soil microflora alterations. Appl. Soil Ecol. 2016, 101, 28–36. [Google Scholar] [CrossRef]
- Melo, I.S.; Santos, S.N.; Rosa, L.H.; Parma, M.M.; Silva, L.J.; Queiroz, S.C.N.; Pellizari, V.H. Isolation and biological activities of an endophytic Mortierella alpina strain from the Antarctic moss Schistidium antarctici. Extremophiles 2014, 18, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.M.; Wang, G.S.; Li, Y.Y.; Chen, X.S.; Wu, S.J.; Mao, Z.Q. Assessment of fungal diversity in apple replanted orchard soils by T-RFLP analysis. Acta Ecol. Sin. 2014, 34, 837–846. [Google Scholar]
- Chang, K.F.; Hwang, S.F.; Conner, R.L.; Ahmed, H.U.; Zhou, Q.; Turnbull, G.D.; Strelkov, S.E.; Mclaren, D.L.; Gossen, B.D. First report of Fusarium proliferatum causing root rot in soybean (Glycine max L.) in Canada. Crop Prot. 2015, 67, 52–58. [Google Scholar] [CrossRef]
- Zou, Q.J.; Wang, S.T.; Liang, K.J.; Wang, Y.N.; Tong-Le, H.U.; Han, Z.Q.; Cao, K.Q. Suspected pathogenic Fusarium spp. isolated from apple orchard soils in Hebei Province. Mycosystema 2014, 33, 976–983. [Google Scholar]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Mazzola, M.; Reardon, C.L.; Brown, J. Initial Pythium species composition and Brassicaceae seed meal type influence extent of Pythium-induced plant growth suppression in soil. Soil Biol. Biochem. 2012, 48, 20–27. [Google Scholar] [CrossRef]
- Arnault, I.; Huchette, O.; Auger, J.; Neffati, M.; Belgacem, A.O.; Elmourid, M. Characterisation of aroma "type" in Allium species according to their S-alk(en)yl cysteine sulfoxides and γ-glutamyl dipeptides contents. Acta Hort. 2010, 53, 171–182. [Google Scholar] [CrossRef]
- Gerik, J.S. Evaluation of soil fumigants applied by drip irrigation for liatris production. Plant Dis. 2007, 89, 883–887. [Google Scholar] [CrossRef]
- Fritsch, J. Dimethyl disulfide as a new chemical potential alternative to methyl bromide in soil disinfestation in France. Acta Hort. 2005, 698, 71–76. [Google Scholar] [CrossRef]
- Mazzola, M.; Granatstein, D.M.; Elfving, D.C.; Mullinix, K. Suppression of specific apple root pathogens by Brassica napus seed meal amendment regardless of glucosinolate content. Phytopathology 2001, 91, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.F.; Yamasaki, H.; Mazzola, M. Brassica napus seed meal soil amendment modifies microbial community structure, nitric oxide production and incidence of Rhizoctonia root rot. Soil Biol. Biochem. 2005, 37, 1215–1227. [Google Scholar] [CrossRef]
- Larkin, R.P.; Honeycutt, C.W. Effects of different 3-yr cropping systems on soil microbial communities and soilborne disease of potato. Phytopathology 2006, 96, 68–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolinska, U. Survival of Sclerotium cepivorum and Fusarium oxysporum chlamydospores in soil amended with cruciferous residues. J. Phytopathol. 2000, 148, 343–349. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Date | Treatment | Plant Height (cm) | Trunk Diameter (mm) | Total Length of Branch (cm) | Dry Weight (g) |
|---|---|---|---|---|---|
| 15 October 2015 | CK | 152.00 ± 2.31c | 19.91 ± 0.35 c | 312.67 ± 15.17 c | 540.24 ± 25.55 c |
| T1 | 204.67 ± 6.33 a | 26.08 ± 1.40 a | 589.67 ± 32.11 a | 890.07 ± 69.07 a | |
| T2 | 171.33 ± 3.53 b | 22.98 ± 0.47 b | 477.00 ± 47.35 b | 669.14 ± 38.41 b | |
| T3 | 170.33 ± 3.76 b | 22.83 ± 0.30 b | 536.00 ± 19.70 ab | 682.28 ± 34.27 b | |
| 15 October 2016 | CK | 184.67 ± 3.48 c | 123.33 ± 1.20 c | 455.67 ± 34.36 c | 834.67 ± 11.85 c |
| T1 | 218.33 ± 0.88 a | 143.33 ± 3.18 a | 678.00 ± 18.00 a | 1167.00 ± 34.49 a | |
| T2 | 196.67 ± 2.91 b | 134.67 ± 0.67 b | 574.33 ± 34.84 b | 1037.33 ± 32.44 b | |
| T3 | 197.67 ± 1.67 b | 133.00 ± 3.06 b | 612.33 ± 20.22 ab | 1098.67 ± 20.18 b |
| Plant Height (cm) | Dry Weight (cm) | Death Rate (%) | |
|---|---|---|---|
| Inoculated | 8.43 ± 0.18 b | 2.80 ± 0.57 b | 56.67% |
| Uninoculated | 14.74 ± 0.15 a | 4.56 ± 0.07 a | NF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Wang, G.; Liu, X.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. Control of Apple Replant Disease Using Mixed Cropping with Brassica juncea or Allium fistulosum. Agriculture 2022, 12, 68. https://doi.org/10.3390/agriculture12010068
Zhao L, Wang G, Liu X, Chen X, Shen X, Yin C, Mao Z. Control of Apple Replant Disease Using Mixed Cropping with Brassica juncea or Allium fistulosum. Agriculture. 2022; 12(1):68. https://doi.org/10.3390/agriculture12010068
Chicago/Turabian StyleZhao, Lei, Gongshuai Wang, Xin Liu, Xuesen Chen, Xiang Shen, Chengmiao Yin, and Zhiquan Mao. 2022. "Control of Apple Replant Disease Using Mixed Cropping with Brassica juncea or Allium fistulosum" Agriculture 12, no. 1: 68. https://doi.org/10.3390/agriculture12010068