
Survival of Salmonella Typhimurium, Listeria monocytogenes, and ESBL Carrying Escherichia coli in Stored Anaerobic Biogas Digestates in Relation to Different Biogas Input Materials and Storage Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Anaerobic Digestate Preparation
2.2. Pathogen Inoculum Preparation
2.3. Experimental Setup
2.4. Quantitative Microbiological Analyses during Storage Experiments
2.5. Data Analysis
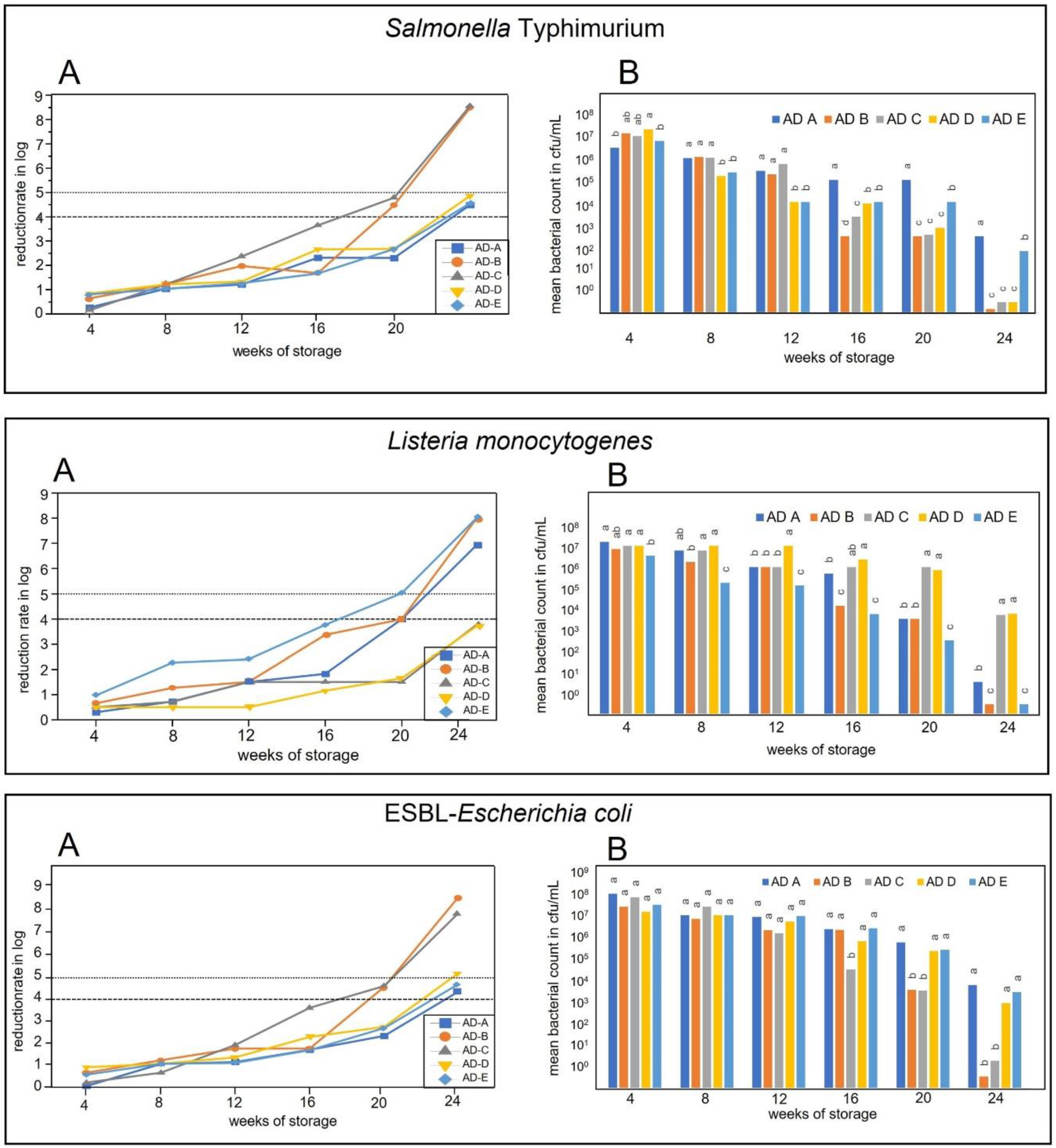
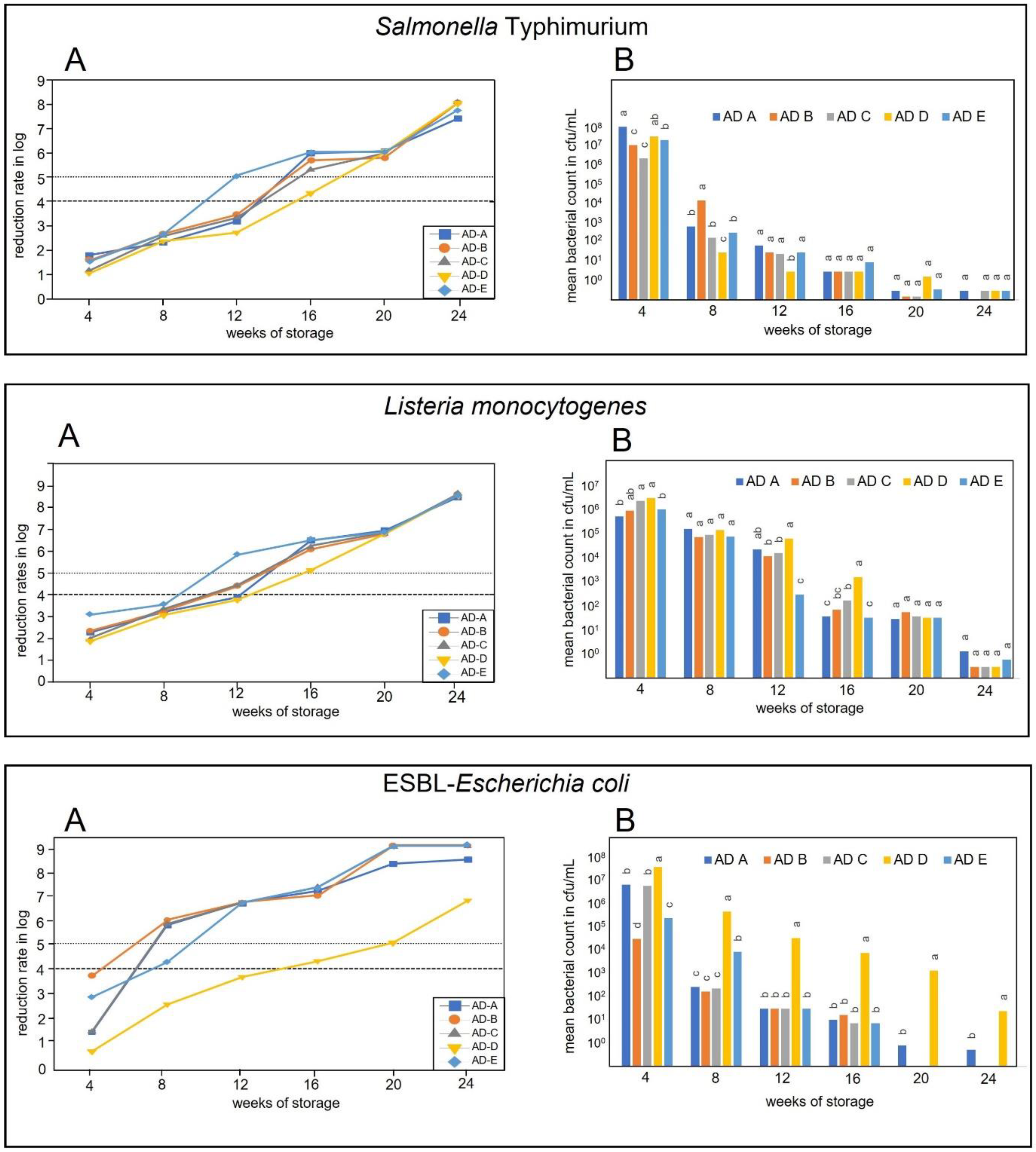
3. Results
3.1. Analysis of the Anaerobic Biogas Digestates
3.2. Survival of Salmonella Typhimurium, Listeria monocytogenes, and ESBL Carrying Escherichia coli
3.2.1. Inactivation during Storage in Temperature Profile 1 Conditions (January to June)
3.2.2. Inactivation during Storage in Temperature Profile 2 Conditions (April to September)
3.2.3. Inactivation during Storage in Temperature Profile 3 Conditions (July to December)
3.2.4. Inactivation during Storage in Temperature Profile 4 Conditions (October to March)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sahlström, L. A review of survival of pathogenic bacteria in organic waste used in biogas plants. Bioresour. Technol. 2003, 87, 161–166. [Google Scholar] [CrossRef]
- Hutchison, M.L.; Walters, L.D.; Avery, S.M.; Munro, F.; Moore, A. Analyses of livestock production, waste storage, and pathogen levels and prevalences in farm manures. Appl. Environ. Microbiol. 2005, 71, 1231–1236. [Google Scholar] [CrossRef] [Green Version]
- Gerba, C.P.; Smith, J.E., Jr. Sources of pathogenic microorganisms and their fate during land application of wastes. J. Environ. Qual. 2005, 34, 42–48. [Google Scholar]
- Sidhu, J.P.; Toze, S.G. Human pathogens and their indicators in biosolids: A literature review. Environ. Int. 2009, 35, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-spectrum β-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals: A threat for humans? Clin. Microbiol. Infect. 2017, 23, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Collis, R.M.; Burgess, S.A.; Biggs, P.J.; Midwinter, A.C.; French, N.P.; Toombs-Ruane, L.; Cookson, A.L. Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Dairy Farm Environments: A New Zealand Perspective. Foodborne Pathog. Dis. 2019, 16, 5–22. [Google Scholar] [CrossRef]
- Smith, S.R.; Lang, N.L.; Cheung, K.H.; Spanoudaki, K. Factors controlling pathogen destruction during anaerobic digestion of biowastes. Waste Manag. 2005, 25, 417–425. [Google Scholar] [CrossRef]
- Goberna, M.; Podmirseg, S.M.; Waldhuber, S.; Knapp, B.A.; García, C.; Insam, H. Pathogenic bacteria and mineral N in soils following the land spreading of biogas digestates and fresh manure. Appl. Soil Ecol. 2011, 49, 18–25. [Google Scholar] [CrossRef]
- Wagner, A.O.; Malin, C.; Gstraunthaler, G.; Illmer, P. Survival of selected pathogens in diluted sludge of a thermophilic waste treatment plant and in NaCl-solution under aerobic and anaerobic conditions. Waste Manag. 2009, 29, 425–429. [Google Scholar] [CrossRef]
- Wagner, A.O.; Gstraunthaler, G.; Illmer, P. Survival of bacterial pathogens during the thermophilic anaerobic digestion of biowaste: Laboratory experiments and in situ validation. Anaerobe 2008, 14, 181–183. [Google Scholar] [CrossRef]
- Avery, L.M.; Anchang, K.Y.; Tumwesige, V.; Strachan, N.; Goude, P.J. Potential for Pathogen reduction in anaerobic digestion and biogas generation in Sub-Saharan Africa. Biomass Bioenerg. 2014, 70, 112–124. [Google Scholar] [CrossRef]
- Fu, B.; Jiang, Q.; Liu, H.B.; Liu, H. Quantification of viable but nonculturable Salmonella spp. and Shigella spp. during sludge anaerobic digestion and their reactivation during cake storage. J. Appl. Microbiol. 2015, 119, 1138–1147. [Google Scholar] [CrossRef]
- Resende, J.A.; Diniz, C.G.; Silva, V.L.; Otenio, M.H.; Bonnafous, A.; Arcuri, P.B.; Godon, J.J. Dynamics of antibiotic resistance genes and presence of putative pathogens during ambient temperature anaerobic digestion. J. Appl. Microbiol. 2014, 117, 1689–1699. [Google Scholar] [CrossRef]
- Schauss, T.; Glaeser, S.P.; Gütschow, A.; Dott, W.; Kämpfer, P. Improved detection of extended spectrum beta-lactamase (ESBL)-producing Escherichia coli in input and output samples of German biogas plants by a selective pre-enrichment procedure. PLoS ONE 2015, 10, e0119791. [Google Scholar] [CrossRef]
- von Salviati, C.; Laube, H.; Guerra, B.; Roesler, U.; Friese, A. Emission of ESBL/AmpC-producing Escherichia coli from pig fattening farms to surrounding areas. Vet. Microbiol. 2015, 175, 77–84. [Google Scholar] [CrossRef]
- Alfa, M.I.; Adie, D.B.; Igboro, S.B.; Oranusi, U.S.; Dahunsi, S.O.; Akali, D.M. Assessment of biofertilizer quality and health implications of anaerobic digestion effluent of cow dung and chicken droppings. Renew. Energy 2014, 63, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Orzi, V.; Scaglia, B.; Lonati, S.; Riva, C.; Boccasile, G.; Alborali, G.L.; Adani, F. The role of biological processes in reducing both odor impact and pathogen content during mesophilic anaerobic digestion. Sci. Total Environ. 2015, 526, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Himathongkham, S.; Bahari, S.; Riemann, H.; Cliver, D. Survival of Escherichia coli O157:H7 and Salmonella typhimurium in cow manure and cow manure slurry. FEMS Microbiol. Lett. 1999, 178, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Kudva, I.T.; Blanch, K.; Hovde, C.J. Analysis of Escherichia coli O157:H7 survival in ovine or bovine manure and manure slurry. Appl. Environ. Microbiol. 1998, 64, 3166–3174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, F.A.; Groves, S.J.; Chambers, B.J. Pathogen survival during livestock manure storage and following land application. Bioresour. Technol. 2005, 96, 135–143. [Google Scholar] [CrossRef]
- You, Y.; Rankin, S.C.; Aceto, H.W.; Benson, C.E.; Toth, J.D.; Dou, Z. Survival of Salmonella enterica serovar Newport in manure and manure-amended soils. Appl. Environ. Microbiol. 2006, 72, 5777–5783. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.W.; Bew, J.; Burrows, M.R.; Matthews, P.R.; Collins, P. The occurrence of salmonellas, mycobacteria and pathogenic strains of Escherichia coli in pig slurry. Epidemiol. Infect. 1976, 77, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Doyle, M.P.; Shere, J.; Garber, L. Prevalence of enterohemorrhagic Escherichia coli O157:H7 in a survey of dairy herds. Appl. Environ. Microbiol. 1995, 61, 1290–1293. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Zhao, T.; Doyle, M.P. Fate of enterohemorrhagic Escherichia coli O157:H7 in bovine feces. Appl. Environ. Microbiol. 1996, 62, 2567–2570. [Google Scholar] [CrossRef] [Green Version]
- Stanley, K.N.; Wallace, J.S.; Currie, J.E.; Diggle, P.J.; Jones, K. The seasonal variation of thermophilic campylobacters in beef cattle, dairy cattle and calves. J. Appl. Microbiol. 1998, 85, 472–480. [Google Scholar] [CrossRef]
- Himathongkham, S.; Riemann, H. Destruction of Salmonella typhimurium, Escherichia coli O157:H7 and Listeria monocytogenes in chicken manure by drying and/or gassing with ammonia. FEMS Microbiol. Lett. 1999, 171, 179–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plachá, I.; Venglovský, J.; Sasáková, N.; Svoboda, I.F. The effect of summer and winter seasons on the survival of Salmonella typhimurium and indicator micro-organisms during the storage of solid fraction of pig slurry. J. Appl. Microbiol. 2001, 91, 1036–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piepho, H.-P. An Algorithm for a Letter-Based Representation of All-Pairwise Comparisons. J. Comput. Graph. Stat. 2004, 13, 456–466. [Google Scholar] [CrossRef]
- Piepho, H.-P. A SAS macro for generating letter displays of pairwise mean comparisons. Commun. Biom. Crop Sci. 2012, 7, 4–13. [Google Scholar]
- Fröschle, B.; Heiermann, M.; Lebuhn, M.; Messelhäusser, U.; Plöchl, M. Hygiene and Sanitation in Biogas Plants. Biogas Sci. Technol. 2015, 151, 63–99. [Google Scholar] [CrossRef]
- Le Maréchal, C.; Druilhe, C.; Repérant, E.; Boscher, E.; Rouxel, S.; Le Roux, S.; Poëzévara, T.; Ziebal, C.; Houdayer, C.; Nagard, B.; et al. Evaluation of the occurrence of sporulating and nonsporulating pathogenic bacteria in manure and in digestate of five agricultural biogas plants. Microbiologyopen 2019, 8, e872. [Google Scholar] [CrossRef]
- Costa, A.; Gusmara, C.; Gardoni, D.; Zaninelli, M.; Tambone, F.; Sala, V.; Guarino, M. The effect of anaerobic digestion and storage on indicator microorganisms in swine and dairy manure. Environ. Sci. Pollut. Res. Int. 2017, 24, 24135–24146. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, G.; Lawlor, P.G.; Gutierrez, M.; O’Sullivan, L.; Murphy, A.; Zhan, X.; Gardiner, G.E. An assessment of Salmonella survival in pig manure and its separated solid and liquid fractions during storage. J. Environ. Sci. Health. Part B 2015, 50, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Luo, F.; Jiang, X. Factors impacting the regrowth of Escherichia coli O157:H7 in dairy manure compost. J. Food Prot. 2009, 72, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shepherd, M.W., Jr.; Jiang, X. Evaluating the effect of environmental factors on pathogen regrowth in compost extract. Microb. Ecol. 2009, 58, 498–508. [Google Scholar] [CrossRef]
- Chen, Z.; Kim, J.; Jiang, X. Survival of Escherichia coli O157:H7 and Salmonella enterica in animal waste-based composts as influenced by compost type, storage condition and inoculum level. J. Appl. Microbiol. 2018, 124, 1311–1323. [Google Scholar] [CrossRef]
- Wang, H.; Dharmasena, M.; Chen, Z.; Jiang, X. Persistence of Non-O157 Shiga Toxin-Producing Escherichia coli in Dairy Compost during Storage. J. Food Prot. 2017, 80, 1999–2005. [Google Scholar] [CrossRef]
- Fukushima, H.; Hoshina, K.; Gomyoda, M. Long-term survival of shiga toxin-producing Escherichia coli O26, O111, and O157 in bovine feces. Appl. Environ. Microbiol. 1999, 65, 5177–5181. [Google Scholar] [CrossRef] [Green Version]
- Vinnerås, B. Comparison of composting, storage and urea treatment for sanitising of faecal matter and manure. Bioresour. Technol. 2007, 98, 3317–3321. [Google Scholar] [CrossRef]
- Ajariyakhajorn, C.; Goyal, S.M.; Robinson, R.A.; Johnston, L.J.; Clanton, C.A. The survival of Salmonella anatum, pseudorabies virus and porcine reproductive and respiratory syndrome virus in swine slurry. New Microbiol. 1997, 20, 365–369. [Google Scholar]
- Mawdsley, J.L.; Bardgett, R.D.; Merry, R.J.; Pain, B.F.; Theodorou, M.K. Pathogens in livestock waste, their potential for movement through soil and environmental pollution. Appl. Soil Ecol. 1995, 2, 1–15. [Google Scholar] [CrossRef]
- Arrus, K.M.; Holley, R.A.; Ominski, K.H.; Tenuta, M.; Blank, G. Influence of temperature on Salmonella survival in hog manure slurry and seasonal temperature profiles in farm manure storage reservoirs. Livest. Sci. 2006, 102, 226–236. [Google Scholar] [CrossRef]
- Semenov, A.V.; van Bruggen, A.H.; van Overbeek, L.; Termorshuizen, A.J.; Semenov, A.M. Influence of temperature fluctuations on Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in cow manure. FEMS Microbiol. Ecol. 2007, 60, 419–428. [Google Scholar] [CrossRef] [Green Version]
- García, R.; Baelum, J.; Fredslund, L.; Santorum, P.; Jacobsen, C.S. Influence of temperature and predation on survival of Salmonella enterica serovar Typhimurium and expression of invA in soil and manure-amended soil. Appl. Environ. Microbiol. 2010, 76, 5025–5031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, X.T.; Wolff, A.; Madsen, M.; Bang, D.D. Fate and Survival of Campylobacter coli in Swine Manure at Various Temperatures. Front. Microbiol. 2011, 2, 262. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Brown, L.; Ashbolt, N.J.; Stuetz, R.M.; Roser, D.J. Inactivation of indicators and pathogens in cattle feedlot manures and compost as determined by molecular and culture assays. FEMS Microbiol. Ecol. 2011, 77, 200–210. [Google Scholar] [CrossRef]
- Park, G.W.; Diez-Gonzalez, F. Utilization of carbonate and ammonia-based treatments to eliminate Escherichia coli O157:H7 and Salmonella Typhimurium DT104 from cattle manure. J. Appl. Microbiol. 2003, 94, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Lemunier, M.; Francou, C.; Rousseaux, S.; Houot, S.; Dantigny, P.; Piveteau, P.; Guzzo, J. Long-term survival of pathogenic and sanitation indicator bacteria in experimental biowaste composts. Appl. Environ. Microbiol. 2005, 71, 5779–5786. [Google Scholar] [CrossRef] [Green Version]
- Paniel, N.; Rousseaux, S.; Gourland, P.; Poitrenaud, M.; Guzzo, J. Assessment of survival of Listeria monocytogenes, Salmonella Infantis and Enterococcus faecalis artificially inoculated into experimental waste or compost. J. Appl. Microbiol. 2010, 108, 1797–1809. [Google Scholar] [CrossRef]
- Bonetta, S.; Ferretti, E.; Bonetta, S.; Fezia, G.; Carraro, E. Microbiological contamination of digested products from anaerobic co-digestion of bovine manure and agricultural by-products. Lett. Appl. Microbiol. 2011, 53, 552–557. [Google Scholar] [CrossRef]
- Maynaud, G.; Pourcher, A.M.; Ziebal, C.; Cuny, A.; Druilhe, C.; Steyer, J.P.; Wéry, N. Persistence and Potential Viable but Non-culturable State of Pathogenic Bacteria during Storage of Digestates from Agricultural Biogas Plants. Front. Microbiol. 2016, 7, 1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piveteau, P.; Depret, G.; Pivato, B.; Garmyn, D.; Hartmann, A. Changes in gene expression during adaptation of Listeria monocytogenes to the soil environment. PLoS ONE 2011, 6, e24881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivant, A.L.; Desneux, J.; Pourcher, A.M.; Piveteau, P. Transcriptomic Analysis of the Adaptation of Listeria monocytogenes to Lagoon and Soil Matrices Associated with a Piggery Environment: Comparison of Expression Profiles. Front. Microbiol. 2017, 8, 1811. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AD | Input Material of the Biogas Plant |
|---|---|
| A | 70% pig and cattle slurry 30% maize silage |
| B | 60% pig slurry 40% maize silage |
| C | 60% cattle slurry 40% horse, cattle and poultry manure and maize silage |
| D | 60% poultry slurry and poultry manure 40% maize silage |
| E | 60% cattle slurry, cattle, and horse manure 40% maize silage |
| Month | Jan | Feb | March | April | May | June | July | Aug | Sept | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | 1.1 | 1.9 | 5.5 | 10.2 | 14.3 | 17.8 | 19.1 | 18.8 | 14.5 | 10.3 | 5.5 | 1.8 |
| profile 1 (winter-spring) 1 |  | |||||||||||
| profile 2 (spring-summer) | | |||||||||||
| profile 3 (summer-autumn) | | |||||||||||
| profile 4 (autumn-winter) |  | | ||||||||||
| Before Storage | After 24 Months of Storage | |||||
|---|---|---|---|---|---|---|
| Anaerobic Digestate | Temperature Profile 1 | Temperature Profile 2 | Temperature Profile 3 | Temperature Profile 4 | ||
| total N 1 % | 0.46 | 0.415 | 0.392 | 0.456 | 0.444 | |
| AD-A | NH4-N 2 % | 0.21 | 0.217 | 0.196 | 0.246 | 0.243 |
| pH | 7.7 | 7.7 | 7.8 | 7.5 | 8.4 | |
| total N % | 0.69 | 0.694 | 0.696 | 0.694 | 0.94 | |
| AD-B | NH4-N % | 0.43 | 0.496 | 0.476 | 0.477 | 0.491 |
| pH | 7.9 | 7.9 | 7.7 | 7.8 | 7.8 | |
| total N % | 0.71 | 0.705 | 0.706 | 0.707 | 0.699 | |
| AD-C | NH4-N % | 0.36 | 0.405 | 0.396 | 0.407 | 0.392 |
| pH | 8.3 | 8.8 | 8.0 | 7.7 | 8.8 | |
| total N % | 0.76 | 0.761 | 0.750 | 0.748 | 0.756 | |
| AD-D | NH4-N % | 0.40 | 0.374 | 0.43 | 0.428 | 0.415 |
| pH | 8.1 | 7.9 | 8.3 | 7.9 | 8.3 | |
| total N % | 0.44 | 0.433 | 0.415 | 0.231 | 0.399 | |
| AD-E | NH4-N % | 0.12 | 0.131 | 0.147 | 0.016 | 0.124 |
| pH | 7.7 | 8.2 | 7.4 | 8.1 | 7.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schilling, T.; Hoelzle, K.; Philipp, W.; Hoelzle, L.E. Survival of Salmonella Typhimurium, Listeria monocytogenes, and ESBL Carrying Escherichia coli in Stored Anaerobic Biogas Digestates in Relation to Different Biogas Input Materials and Storage Temperatures. Agriculture 2022, 12, 67. https://doi.org/10.3390/agriculture12010067
Schilling T, Hoelzle K, Philipp W, Hoelzle LE. Survival of Salmonella Typhimurium, Listeria monocytogenes, and ESBL Carrying Escherichia coli in Stored Anaerobic Biogas Digestates in Relation to Different Biogas Input Materials and Storage Temperatures. Agriculture. 2022; 12(1):67. https://doi.org/10.3390/agriculture12010067
Chicago/Turabian StyleSchilling, Thorben, Katharina Hoelzle, Werner Philipp, and Ludwig E. Hoelzle. 2022. "Survival of Salmonella Typhimurium, Listeria monocytogenes, and ESBL Carrying Escherichia coli in Stored Anaerobic Biogas Digestates in Relation to Different Biogas Input Materials and Storage Temperatures" Agriculture 12, no. 1: 67. https://doi.org/10.3390/agriculture12010067