
Different Mutations Providing Target Site Resistance to ALS- and ACCase-Inhibiting Herbicides in Echinochloa spp. from Rice Fields

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
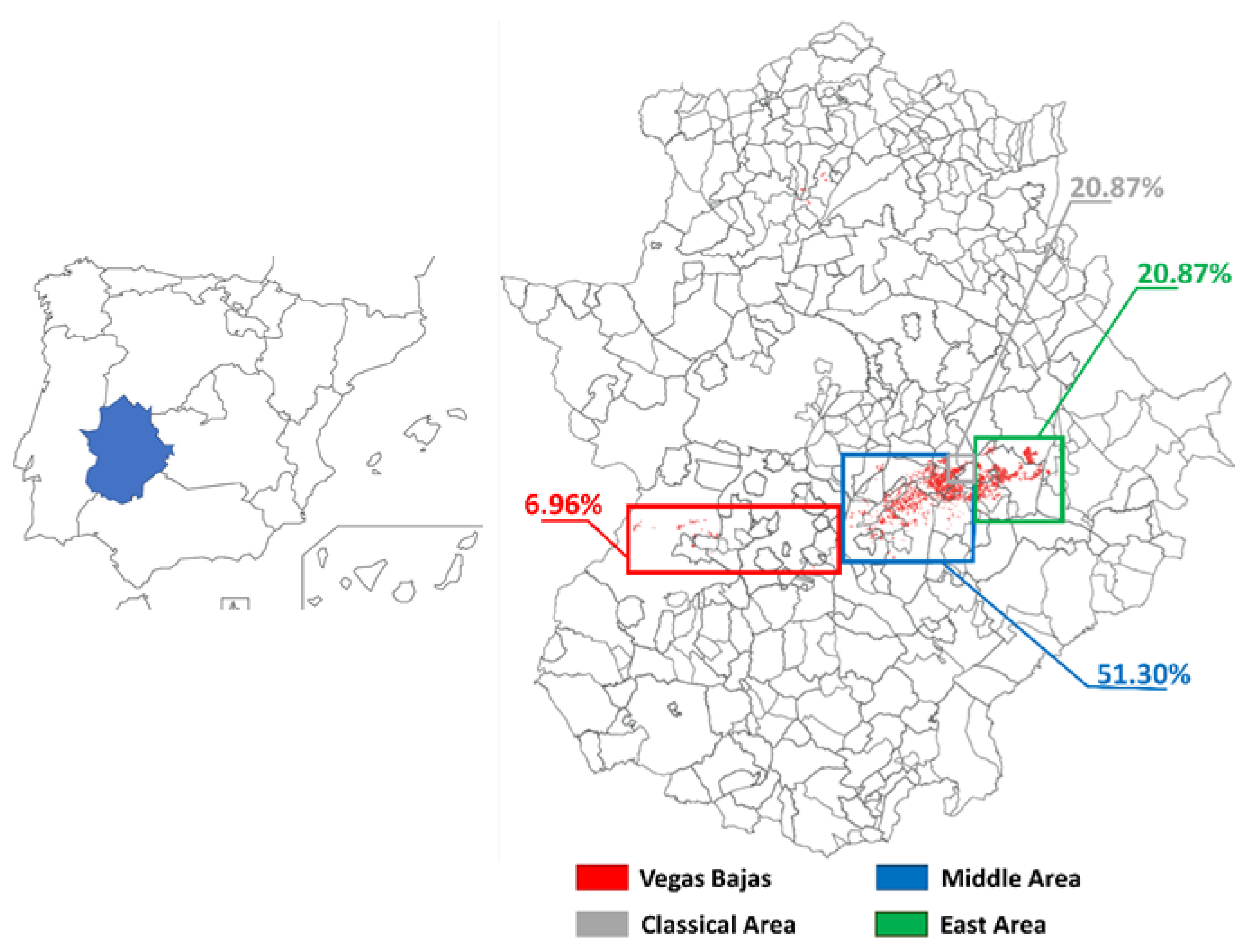
2.1. Surveys
2.2. DNA Extraction and Polymerase Chain Reaction
2.3. Molecular Characterization of Populations
2.4. ALS and ACCase Gene Sequencing, and Polymerase Chain Reaction (PCR)
2.5. Herbicides and Chemicals
2.6. Dose-Response Assays
3. Results
3.1. Surveys and Molecular Characterization
3.2. ALS and ACCase Gene Sequencing
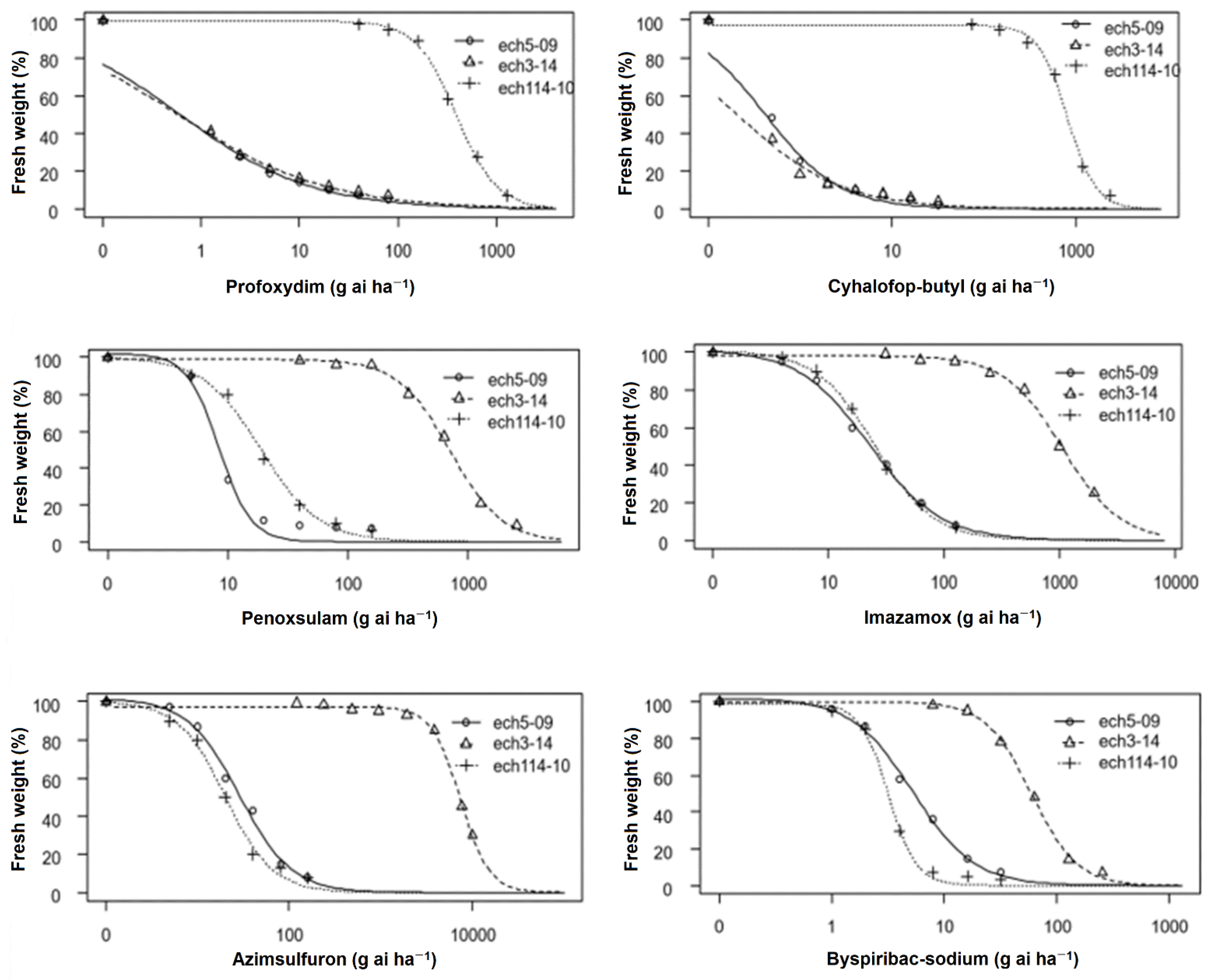
3.3. Dose-Response Assays with ACCase and ALS Inhibitors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MAPAMA—Ministerio de Agricultura y Pesca Alimentación y Medio Ambiente (Spanish Ministry of Agriculture, Fisheries, Food and Environment). Available online: https://www.mapa.gob.es/ (accessed on 21 February 2021).
- Panozzo, S.; Scarabel, L.; Tranel, P.; Sattin, M. Target-site resistance to ALS inhibitors in the polyploid species Echinochloa crus-galli. Pestic. Biochem. Physiol. 2013, 105, 93–101. [Google Scholar] [CrossRef]
- Altop, E.K.; Mennan, H. Genetic and morphologic diversity of Echinochloa crus-galli populations from different origins. Phytoparasitica 2010, 39, 93–102. [Google Scholar] [CrossRef]
- Carretero, J. El género Echinochloa Beauv. en el suroeste de Europa. An. Jard. Bot. Madrid 1981, 38, 91–108. [Google Scholar]
- Tabacchi, M.; Mantegazza, R.; Spada, A.; Ferrero, A. Morphological traits and molecular markers for classification of Echinochloa species from Italian rice fields. Weed Sci. 2006, 54, 1086–1093. [Google Scholar] [CrossRef]
- Fang, J.; He, Z.; Liu, T.; Li, J.; Dong, L. A novel mutation Asp-2078-Glu in ACCase confers resistance to ACCase herbicides in barnyardgrass (Echinochloa crus-galli). Pestic. Biochem. Physiol. 2020, 168, 104634. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org/Home.aspx (accessed on 21 February 2021).
- Shaner, D.L. Mechanisms of resistance to acetolactate synthase/acetohydroxyacid synthase inhibitors. In Proceedings of the Herbicide Resistance in Weeds and Crops; Elsevier: Amsterdam, The Netherlands, 1991; pp. 187–198. [Google Scholar]
- Socorro, R.J.M. Resistencia de Sinapis alba a Herbicidas Inhibidores de la ALS: Bases Agronómicas Bioquímicas y Moleculares. Ph.D. Thesis, University of Cordoba, Cordoba, Spain, 2011. [Google Scholar]
- Ray, T.B. Site of action of chlorsulfuron: Inhibition of valine and isoleucine biosynthesis in plants. Plant Physiol. 1984, 75, 827–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaner, D.L.; Anderson, P.C.; Stidham, M.A. Imidazolinones potent inhibitors of acetohydroxyacid synthase. Plant Physiol. 1984, 76, 545–546. [Google Scholar] [CrossRef] [Green Version]
- Gerwick, B.C.; Mireles, L.C.; Eilers, R.J. Rapid Diagnosis of Als/Ahas-Resistant Weeds. Weed Technol. 1993, 7, 519–524. [Google Scholar] [CrossRef]
- Takahashi, S.; Shigematsu, S.; Morita, A.; Nezu, M.; Claus, J.S.; Williams, C.S. KIH-2031, a new herbicide for cotton. In Proceedings of the Brighton Crop Protection Conference, Weeds (United Kingdom); BCPC: London, UK, 1991; Available online: https://www.yumpu.com/en/document/read/15429707/herbologia-anubih (accessed on 21 February 2021).
- Santel, H.J.; Bowden, B.A.; Sorensen, V.M.; Mueller, K.H. Flucarbazone-sodium—A new herbicide for the selective control of wild oat and green foxtail in wheat. In The 1999 Brighton Conference—Weeds, Proceedings of the International Conference Held at the Brighton Metropole Hotel, Brighton, UK, 15–18 November 1999; British Crop Protection Council: London, UK, 1999; pp. 23–28. [Google Scholar]
- Délye, C. Weed resistance to acetyl coenzyme A carboxylase inhibitors: An update. Weed Sci. 2005, 53, 728–746. [Google Scholar] [CrossRef]
- Muehlebach, M.; Boeger, M.; Cederbaum, F.; Cornes, D.; Friedmann, A.A.; Glock, J.; Niderman, T.; Stoller, A.; Wagner, T. Aryldiones incorporating a [1,4,5]oxadiazepane ring. Part I: Discovery of the novel cereal herbicide pinoxaden. Bioorg. Med. Chem. 2009, 17, 4241–4256. [Google Scholar] [CrossRef]
- Maroli, A.S.; Gaines, T.A.; Foley, M.E.; Duke, S.O.; Doğramacı, M.; Anderson, J.V.; Horvath, D.P.; Chao, W.S.; Tharayil, N. Omics in Weed Science: A Perspective from Genomics, Transcriptomics, and Metabolomics Approaches. Weed Sci. 2018, 66, 681–695. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, V.; Salas-Perez, R.A.; Bagavathiannan, M.V.; Lawton-Rauh, A.; Roma-Burgos, N. Target-site mutation ac-cumulation among ALS inhibitor-resistant Palmer amaranth. Pest Manag. Sci. 2019, 75, 1131–1139. [Google Scholar]
- Deng, W.; Di, Y.; Cai, J.; Chen, Y.; Yuan, S. Target-Site Resistance Mechanisms to Tribenuron-methyl and Cross-resistance Patterns to ALS-inhibiting Herbicides of Catchweed Bedstraw (Galium aparine) with Different ALS Mutations. Weed Sci. 2018, 67, 183–188. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Vargas, J.J.; Breeden, G.K.; Grier, L.; Aponte, R.A.; Tresch, S.; Laforest, M. A new amino acid substitution (Ala-205-Phe) in acetolactate synthase (ALS) confers broad spectrum resistance to ALS-inhibiting herbicides. Planta 2015, 243, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Jha, P. First report of Ser653Asn mutation endowing high-level resistance to imazamox in downy brome (Bromus tectorum L.). Pest Manag. Sci. 2017, 73, 2585–2591. [Google Scholar] [CrossRef] [Green Version]
- Laplante, J.; Rajcan, I.; Tardif, F.J. Multiple allelic forms of acetohydroxyacid synthase are responsible for herbicide resistance in Setaria viridis. Theor. Appl. Genet. 2009, 119, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bi, Y.; Liu, W.; Yuan, G.; Wang, J. Molecular basis for resistance to fenoxaprop-p-ethyl in American sloughgrass (Beckmannia syzigachne Steud.). Pestic. Biochem. Physiol. 2013, 105, 118–121. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Bailly, G.C.; Dale, R.P.; Hutchings, S.-J.; McIndoe, E. A novel W1999S mutation and non-target site resistance impact on acetyl-CoA carboxylase inhibiting herbicides to varying degrees in a UK Lolium multiflorum population. PLoS ONE 2013, 8, e58012. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhu, X.; Wang, H.; Li, J.; Dong, L. Mechanism of resistance to fenoxaprop in Japanese foxtail (Alopecurus japonicus) from China. Pestic. Biochem. Physiol. 2013, 107, 25–31. [Google Scholar] [CrossRef]
- Tang, H.; Li, J.; Dong, L.; Dong, A.; Lu, B.; Zhu, X. Molecular bases for resistance to acetyl-coenzyme A carboxylase inhibitor in Japanese foxtail (Alopecurus japonicus). Pest Manag. Sci. 2012, 68, 1241–1247. [Google Scholar] [CrossRef]
- Kaundun, S.S. An aspartate to glycine change in the carboxyl transferase domain of acetyl CoA carboxylase and non-target-site mechanism(s) confer resistance to Accase inhibitor herbicides in a Lolium multiflorum population. Pest Manag. Sci. 2010, 66, 1249–1256. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Hutchings, S.-J.; Dale, R.P.; McIndoe, E. Broad resistance to ACCase inhibiting herbicides in a ryegrass pop-ulation is due only to a cysteine to arginine mutation in the target enzyme. PLoS ONE 2012, 7, e39759. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Hipolito, H.; Domínguez-Valenzuela, J.A.; Osuna, M.D.; De Prado, R. Resistance mechanism to acetyl coenzyme A carboxylase inhibiting herbicides in Phalaris paradoxa collected in Mexican wheat fields. Plant Soil 2011, 355, 121–130. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Romano, Y.; Mendoza, F.; Palmerín, J.A.; Quiles, J.M.; Amaro-Blanco, I.; Osuna, M.D. Uso de marcadores moleculares para la caracterización de malas hierbas del cultivo del arroz en Extremadura: “Echinochloa spp.” y “Leptochloa spp.”. In Proceedings of the XVI Congreso de la Sociedad Española de Malherbología: Actas, Pamplona-Iruña, Spain, 25–27 Octubre 2017; Servicio de Publicaciones de la Universidad Pública de Navarra: Pamplona, Spain, 2017; pp. 337–342. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Romano-Garcia, Y.; Amaro-Blanco, I.; Quiles-Pecos, J.M.; Palmerín-Romero, J.A.; Torcea, L.; Osuna Ruiz, M.D. Problemática de las malas hierbas en el cultivo del arroz en Extremadura (campaña 2017). Agrícola Vergel Frutic. Hortic. Floric. 2017, 406, 378–381. [Google Scholar]
- Kaloumenos, N.S.; Chatzilazaridou, S.L.; Mylona, P.V.; Polidoros, A.N.; Eleftherohorinos, I.G. Target-site mutation associated with cross-resistance to ALS-inhibiting herbicides in late watergrass (Echinochloa oryzicola Vasing.). Pest Manag. Sci. 2012, 69, 865–873. [Google Scholar] [CrossRef]
- Yuan, J.S.; Tranel, P.J.; Stewart, C.N. Non-target-site herbicide resistance: A family business. Trends Plant Sci. 2007, 12, 6–13. [Google Scholar] [CrossRef]
- Ntoanidou, S.; Kaloumenos, N.; Diamantidis, G.; Madesis, P.; Eleftherohorinos, I. Molecular basis of Cyperus difformis cross-resistance to ALS-inhibiting herbicides. Pestic. Biochem. Physiol. 2016, 127, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Sada, Y.; Ikeda, H.; Kizawa, S. Resistance levels of sulfonylurea-resistantSchoenoplectus juncoides(Roxb.) Palla with various Pro197mutations in acetolactate synthase to imazosulfuron, bensulfuron-methyl, metsulfuron-methyl and imazaquin-ammonium. Weed Biol. Manag. 2013, 13, 53–61. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Powles, S.B. Mutations of the ALS gene endowing resistance to ALS-inhibiting herbicides in Lolium rigidum populations. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 1229–1236. [Google Scholar] [CrossRef]
- Xia, W.; Pan, L.; Li, J.; Wang, Q.; Feng, Y.; Dong, L. Molecular basis of ALS- and/or ACCase-inhibitor resistance in shortawn foxtail (Alopecurus aequalis Sobol.). Pestic. Biochem. Physiol. 2015, 122, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Iwakami, S.; Uchino, A.; Watanabe, H.; Yamasue, Y.; Inamura, T. Isolation and expression of genes for acetolactate synthase and acetyl-CoA carboxylase in Echinochloa phyllopogon, a polyploid weed species. Pest Manag. Sci. 2012, 68, 1098–1106. [Google Scholar] [CrossRef]
- Zagnitko, O.; Jelenska, J.; Tevzadze, G.; Haselkorn, R.; Gornicki, P. An isoleucine/leucine residue in the carboxyltransferase domain of acetyl-CoA carboxylase is critical for interaction with aryloxyphenoxypropionate and cyclohexanedione inhibitors. Proc. Natl. Acad. Sci. USA 2001, 98, 6617–6622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, I.A.; Li, R.; You, Z.; Li, Z. Japanese Foxtail (Alopecurus japonicus) Resistance to Fenoxaprop and Pinoxaden in China. Weed Sci. 2012, 60, 167–171. [Google Scholar] [CrossRef]
- Ruiz-Santaella, J.P.; De Prado, R.; Wagner, J.; Fischer, A.; Gerhards, R. Resistance mechanisms to cyhalofop-butyl in a biotype of Echinochloa phyllopogon (Stapf) Koss. from California. Z. Pflanzenkrankh. Pflanzenschutz-Sonderh. 2006, 20, 95. [Google Scholar]
- Iwakami, S.; Hashimoto, M.; Matsushima, K.-I.; Watanabe, H.; Hamamura, K.; Uchino, A. Multiple-herbicide resistance in Echinochloa crus-galli var. formosensis, an allohexaploid weed species, in dry-seeded rice. Pestic. Biochem. Physiol. 2015, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Osuna, M.D.; Okada, M.; Ahmad, R.; Fischer, A.J.; Jasieniuk, M. Genetic Diversity and Spread of Thiobencarb Resistant Early Watergrass (Echinochloa oryzoides) in California. Weed Sci. 2011, 59, 195–201. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primers | Restriction Enzyme | E. crus-galli/E. hispidula | E. oryzicola/E. oryzoides | E. colona |
|---|---|---|---|---|
| Trna/b1 | ---- | 449 | 481 | 449 |
| Eco RI | 178 + 221 | 481 | 449 | |
| Trnc/d | ---- | 620 | 620 | 620 |
| DraI | 120 + 500 | 620 | 120 + 500 | |
| AluI | 620 | 178 + 447 | 178 + 449 |
| Gene | Primer | Sequence 5′ to 3′ |
|---|---|---|
| ALS | BE1 | GTCTTGGGGCTATGGGATTT |
| BE2 | CGACAGAACAAGGGAGAACA | |
| CAD1F | CGACGTCTTCGCCTACCC | |
| CAD2R | ATCTGCTGCTGGATGTCCTT | |
| ACCase | CRUSS-F | GATTGGCATAGCCGATGAAG |
| CRUSS-R | TGGACAACACCATTGGTAGC | |
| AC-6F | AGCTTGGAGGAATCCCTGTT | |
| AC-6R | GGGTCAAGCCTACCCATACA |
| Herbicides | Chemical Group a | Population | Dose | Labeled Rate |
|---|---|---|---|---|
| g a.i. ha−1 | ||||
| ACCase-inhibiting herbicides | ||||
| Profoxydim b | CHD | ech5-09 | 0, 1.25, 2.5, 5, 10, 20, 40, 80 | 200 |
| ech3-14 | ||||
| ech114-10 | 0, 40, 80, 160, 320, 640, 1280 | |||
| Cyhalofop-butyl | APP | ech5-09 | 0, 0.5, 1, 2, 4, 8, 16, 32 | 300 |
| ech3-14 | ||||
| ech114-10 | 0, 75, 150, 300, 600, 1200, 2400 | |||
| ALS-inhibiting herbicides | ||||
| Penoxsulam | TP | ech5-09 | 0, 5, 10, 20, 40, 80, 160 | 40.8 |
| ech114-10 | ||||
| ech3-14 | 0, 40, 80, 160, 320, 640, 1280, 2560 | |||
| Imazamox | IMI | ech5-09 | 0, 4, 8, 16, 32,64, 128 | 35 |
| ech114-10 | ||||
| ech3-14 | 0, 32, 64, 128, 250, 500, 1000, 2000 | |||
| Azimsulfuron | SU | ech5-09 | 0, 5, 10, 20, 40, 80, 160 | 22.5 |
| ech114-10 | ||||
| ech3-14 | 0, 120, 240, 480, 960, 1920, 3840, 7680, 10,000 | |||
| Byspiribac-sodium | PB | ech5-09 | 0, 1, 2, 4, 8, 16, 32 | 25.5 |
| ech114-10 | ||||
| ech3-14 | 0, 8, 16, 32, 64, 128, 256 | |||
| Populations | The Amino Acid Position, Relative Sequence of Nucleotide and Derived Amino Acid | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 193 | 194 | 195 | 196 | 197 | 198 | 199 | 200 | 201 | 202 | 203 | |
| ech5-09 (S) | ACC | GGC | CAG | GTG | CCC | CGC | CGC | ATG | ATC | GGC | ACC |
| Thr | Gly | Gln | Val | Pro | Arg | Arg | Met | Ile | Gly | Thr | |
| ech3-14 (R) | ACC | GGC | CAG | GTG | TCC | CGC | CGC | ATG | ATC | GGC | ACC |
| Thr | Gly | Gln | Val | Ser | Arg | Arg | Met | Ile | Gly | Thr | |
| Populations | The Amino Acid Position, Relative Sequence of Nucleotide and Derived Amino Acid | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1777 | 1778 | 1779 | 1780 | 1781 | 1782 | 1783 | 1784 | 1785 | 1786 | |
| ech5-09 (S) | GGT | GTT | GAG | AAT | ATA | CAT | GGA | AGT | GCT | GCT |
| Gly | Val | Glu | Asn | Ile | His | Gly | Ser | Ala | Ala | |
| ech114-10 (R) | GGT | GTT | GAG | AAT | CTA | CAT | GGA | AGT | GCT | GCT |
| Gly | Val | Glu | Asn | Leu | His | Gly | Ser | Ala | Ala | |
| Herbicide | Population | d | b | ED50 (g ai ha−1) | RF | p-Value |
|---|---|---|---|---|---|---|
| Profoxydim | ech114-10 (ACCase) (R) | 99.49 | 2.11 | 392.29 ± 13.20 | 643.09 | 0.0001 |
| ech3-14 (ALS) (R) | 100.05 | 0.57 | 0.57 ± 0.10 | 0.93 | 0.0001 | |
| ech5-09 (S) | 100.05 | 0.65 | 0.61 ± 0.10 | - | - | |
| Cyhalofop-butyl | ech114-10 (ACCase) (R) | 97.28 | 2.79 | 817.22 ± 28.85 | 1993.21 | 0.0001 |
| ech3-14 (ALS) (R) | 100.11 | 0.76 | 0.20 ± 0.04 | 0.48 | 0.0001 | |
| ech5-09 (S) | 100.23 | 1.07 | 0.41 ± 0.04 | - | - | |
| Penoxsulam | ech114-10 (ACCase) (R) | 100.17 | 1.78 | 19.00 ± 1.19 | 2.24 | 0.0001 |
| ech3-14 (ALS) (R) | 98.79 | 1.99 | 713.42 ± 37.99 | 84.22 | 0.0001 | |
| ech5-09 (S) | 102.06 | 3.24 | 8.47 ± 0.34 | - | - | |
| Imazamox’ | ech114-10 (ACCase) (R) | 100.97 | 1.71 | 25.32 ± 1.11 | 1.10 | 0.0001 |
| ech3-14 (ALS) (R) | 98.25 | 1.73 | 1059.40 ± 42.95 | 46.22 | 0.0001 | |
| ech5-09 (S) | 101.43 | 1.42 | 22.92 ± 1.18 | - | - | |
| Azimsulfuron | ech114-10 (ACCase) (R) | 100.02 | 1.68 | 20.24 ± 1.02 | 0.69 | 0.0001 |
| ech3-14 (ALS) (R) | 97.46 | 2.75 | 7370.71. ± 209.04 | 254.33 | 0.0001 | |
| ech5-09 (S) | 101.60 | 1.53 | 28.98 ± 4 | - | - | |
| Byspiribac-sodium | ech114-10 (ACCase) (R) | 98.85 | 3.51 | 3.22 ± 0.10 | 0.66 | 0.0001 |
| ech3-14 (ALS) (R) | 99.80 | 2.13 | 59.98 ± 2.46 | 11.38 | 0.0001 | |
| ech5-09 (S) | 101.74 | 1.59 | 5.27 ± 0.27 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaro-Blanco, I.; Romano, Y.; Palmerin, J.A.; Gordo, R.; Palma-Bautista, C.; De Prado, R.; Osuna, M.D. Different Mutations Providing Target Site Resistance to ALS- and ACCase-Inhibiting Herbicides in Echinochloa spp. from Rice Fields. Agriculture 2021, 11, 382. https://doi.org/10.3390/agriculture11050382
Amaro-Blanco I, Romano Y, Palmerin JA, Gordo R, Palma-Bautista C, De Prado R, Osuna MD. Different Mutations Providing Target Site Resistance to ALS- and ACCase-Inhibiting Herbicides in Echinochloa spp. from Rice Fields. Agriculture. 2021; 11(5):382. https://doi.org/10.3390/agriculture11050382
Chicago/Turabian StyleAmaro-Blanco, Ignacio, Yolanda Romano, Jose Antonio Palmerin, Raquel Gordo, Candelario Palma-Bautista, Rafael De Prado, and María Dolores Osuna. 2021. "Different Mutations Providing Target Site Resistance to ALS- and ACCase-Inhibiting Herbicides in Echinochloa spp. from Rice Fields" Agriculture 11, no. 5: 382. https://doi.org/10.3390/agriculture11050382