Identification of a Novel QTL for Chlorate Resistance in Rice (Oryza sativa L.)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mapping Population and Potassium Chlorate Treatment
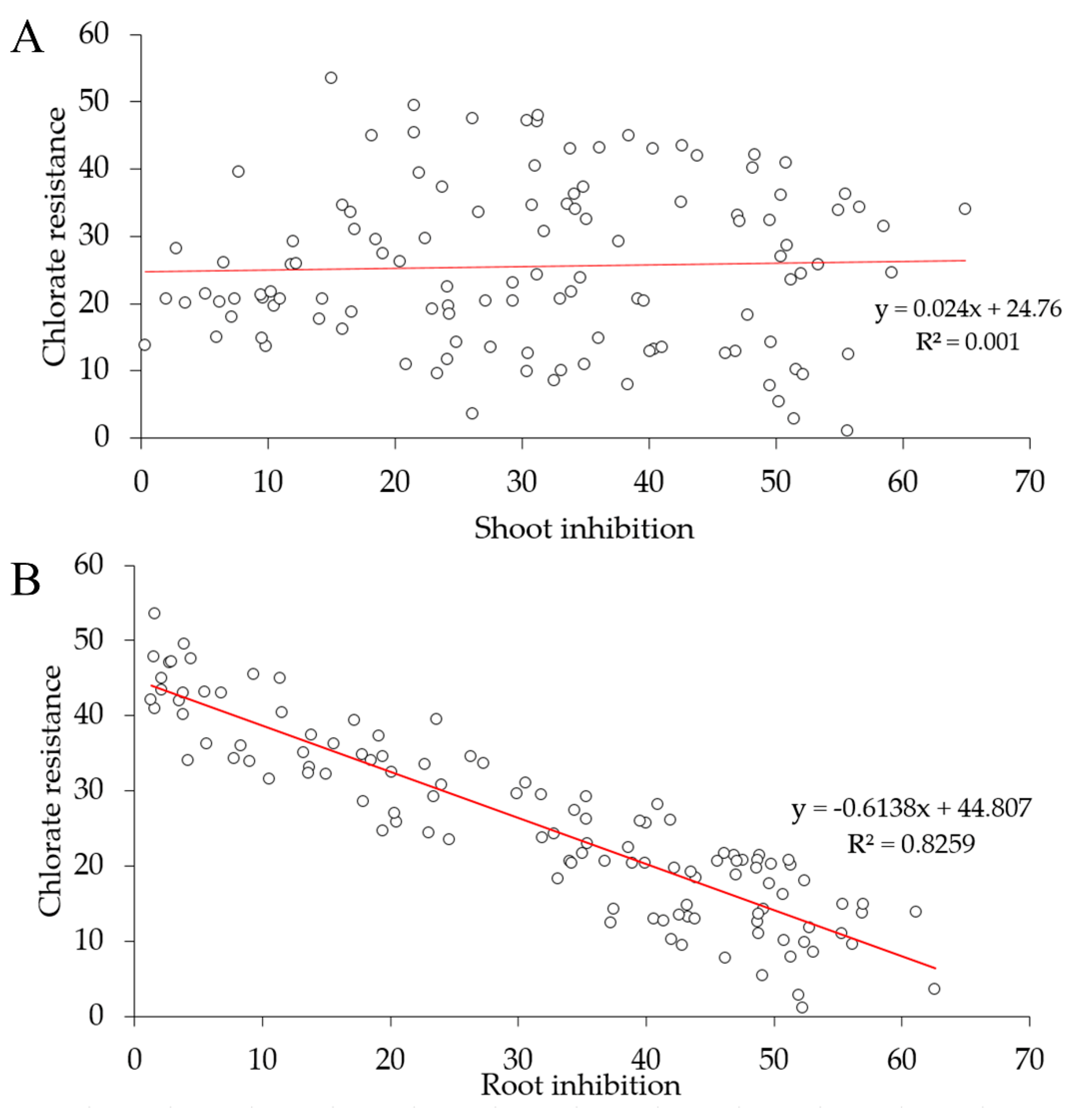
2.2. Q–Q Plot, Frequency Distribution, Correlation Analysis, and Principal Component Analysis
2.3. Construction of Linkage Map and QTL Analysis
2.4. Genomic DNA Extraction and Genotyping
2.5. Comparative Alignment of Coding Sequences (CDS) of Candidate Genes between Indica and Japonica Subspecies
3. Results
3.1. Differential Phenotypic Response of Parental Genotypes and DH Lines
3.2. Relatedness and Principal Component Analysis (PCA)
3.3. Novel QTL for Chlorate Resistance Detected on Chromosome 3 in Rice
3.4. InDel Markers Amplified Polymorphic Bands between Parental Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, M.T.; Bandillo, N.; Al Shiblawi, F.R.A.; Sharma, S.; Liu, K.; Du, Q.; Schmitz, A.J.; Zhang, C.; Véry, A.-A.; Lorenz, A.J. Allelic variants of OsHKT1; 1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content. PLoS Genet. 2017, 13, e1006823. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Motohashi, R.; Tsuchimoto, S.; Fukuta, Y.; Ohtsubo, H.; Ohtsubo, E. Polyphyletic origin of cultivated rice: Based on the interspersion pattern of SINEs. Mol. Biol. Evol. 2003, 20, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, T.; Yu, S.; Li, Y.; Huang, Y.; Huang, T.; Zhang, L.; Zhu, J.; Zhao, Q.; Fan, D. A collection of 10,096 indica rice full-length cDNAs reveals highly expressed sequence divergence between Oryza sativa indica and japonica subspecies. Plant Mol. Biol. 2007, 65, 403–415. [Google Scholar] [CrossRef]
- Ni, J.; Colowit, P.M.; Mackill, D.J. Evaluation of genetic diversity in rice subspecies using microsatellite markers. Crop Sci. 2002, 42, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Ashikari, M. Rice genomics, genetics and breeding; Springer: Berlin/Heidlberg, Germany, 2018. [Google Scholar]
- Teng, S.; Tian, C.; Chen, M.; Zeng, D.; Guo, L.; Zhu, L.; Han, B.; Qian, Q.J.E. QTLs and candidate genes for chlorate resistance in rice (Oryzasativa L.). Euphytica 2006, 152, 141–148. [Google Scholar] [CrossRef]
- Reflinur, B.K.; Lestari, P.; Akter, M.B.; Koh, H.-J. Identification of QTLs Associated with indica-japonica Differentiation-Related Traits in Rice (Oryza sativa L.). Plant Breed. Biotechnol. 2018, 6, 193–205. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, C. Nitrogen-use divergence between indica and japonica rice: Variation at nitrate assimilation. Mol. Plant 2020, 13, 6–7. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Wang, D.; Ruan, B.; Liu, C.; Jiang, H. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Duan, D.; Zhang, H. A single SNP in NRT1. 1B has a major impact on nitrogen use efficiency in rice. Sci. China Life Sci. 2015, 58, 827. [Google Scholar] [CrossRef] [Green Version]
- Oka, H. Intervarietal variation and classification of cultivated rice. Ind. J. Genet. Plant Breed. 1958, 18, 79–89. [Google Scholar]
- Mészáros, A.; Pauk, J. Chlorate resistance as a tool to study the effect of nitrate reductase antisense gene in wheat. Cereal Res. Commun. 2002, 30, 245–252. [Google Scholar] [CrossRef]
- Nakagawa, H.; Yamashita, N. Chlorate reducing activity of spinach nitrate reductase. Agric. Biol. Chem. 1986, 50, 1893–1894. [Google Scholar]
- Solomonson, L.; Vennesland, B. Nitrate reductase and chlorate toxicity in Chlorella vulgaris Beijerinck. Plant Physiol. 1972, 50, 421–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roldan, M.; Reyes, F.; Moreno-Vivian, C.; Castillo, F. Chlorate and nitrate reduction in the phototrophic bacteriaRhodobacter capsulatus andRhodobacter sphaeroides. Curr. Microbiol. 1994, 29, 241–245. [Google Scholar] [CrossRef]
- LaBrie, S.T.; Wilkinson, J.Q.; Crawford, N.M. Effect of chlorate treatment on nitrate reductase and nitrite reductase gene expression in Arabidopsis thaliana. Plant Physiol. 1991, 97, 873–879. [Google Scholar] [CrossRef] [Green Version]
- Rusmana, I.; Nedwell, D.B. Use of chlorate as a selective inhibitor to distinguish membrane-bound nitrate reductase (Nar) and periplasmic nitrate reductase (Nap) of dissimilative nitrate reducing bacteria in sediment. FEMS Microbiol. Ecol. 2004, 48, 379–386. [Google Scholar] [CrossRef]
- Li, H.-S.Z.X.-Y.; Zeng, X.-Y.; Nie, C.-R. Toxic effects of potassium chlorate on peanut growth. J. Plant Ecol. 2006, 30, 124–131. [Google Scholar]
- Lu, J.; Yang, R.; Wang, H.; Huang, X. Stress effects of chlorate on longan (Dimocarpus longan Lour.) trees: Changes in nitrogen and carbon nutrition. Hortic. Plant J. 2017, 3, 237–246. [Google Scholar] [CrossRef]
- Clark, I.C.; Melnyk, R.A.; Iavarone, A.T.; Novichkov, P.S.; Coates, J.D. Chlorate reduction in S hewanella algae ACDC is a recently acquired metabolism characterized by gene loss, suboptimal regulation and oxidative stress. Mol. Microbiol. 2014, 94, 107–125. [Google Scholar] [CrossRef]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-responsive mechanisms in plant leaves revealed by proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, W.S.; Moore, J.H. Genome-Wide Association Studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhorn, J.N.; Daly, M.J. Genome-Wide Association Studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Famoso, A.N.; Zhao, K.; Clark, R.T.; Tung, C.-W.; Wright, M.H.; Bustamante, C.; Kochian, L.V.; McCouch, S.R. Genetic architecture of aluminum tolerance in rice (Oryza sativa) determined through genome-wide association analysis and QTL mapping. PLoS Genet. 2011, 7, e1002221. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Rai, V. Plant Omics and Crop Breeding; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kim, S.; Tang, H.; Mardis, E.R. Genome sequencing technology and algorithms; Artech House, Inc.: Norwook, MA, USA, 2007. [Google Scholar]
- Kasahara, M.; Morishita, S. Large-scale genome sequence processing; Imperial College Press: London, UK, 2006. [Google Scholar]
- Kovach, M.J.; Sweeney, M.T.; McCouch, S.R. New insights into the history of rice domestication. Trends Genet. 2007, 23, 578–587. [Google Scholar] [CrossRef]
- Gondro, C.; Van der Werf, J.; Hayes, B. Genome-wide association studies and genomic prediction; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Agrawal, G.K.; Iwahashi, H.; Rakwal, R. Rice MAPKs. Biochem. Biophys. Res. Commun. 2003, 302, 171–180. [Google Scholar] [CrossRef]
- Garrick, D.J.; Fernando, R.L. Implementing a QTL detection study (GWAS) using genomic prediction methodology. In Genome-Wide Association Studies and Genomic Prediction; Springer: Berlin/Heidelberg, Germany, 2013; pp. 275–298. [Google Scholar]
- Lee, S.-M.; Kang, J.-W.; Lee, J.-Y.; Seo, J.; Shin, D.; Cho, J.-H.; Jo, S.; Song, Y.-C.; Park, D.-S.; Ko, J.-M. QTL Analysis for Fe and Zn Concentrations in Rice Grains Using a Doubled Haploid Population Derived from a Cross Between Rice (Oryza sativa) Cultivar 93-11 and Milyang 352. Plant Breed. Biotechnol. 2020, 8, 69–76. [Google Scholar] [CrossRef]
- Mutinda, Y.; Muthomi, J.; Kimani, J.; George Cheminigw’wa, G.; Olubayo, F. Viability and dormancy of rice seeds after storage and pre-treatment with dry heat and chemical agents. J. Agric. Sci. 2017, 9, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, R. Multiple group principal component analysis and population differentiation. J. Zool. 1988, 216, 37–40. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Cheon, K.-S.; Baek, J.; Cho, Y.-i.; Jeong, Y.-M.; Lee, Y.-Y.; Oh, J.; Won, Y.J.; Kang, D.-Y.; Oh, H.; Kim, S.L. Single nucleotide polymorphism (SNP) discovery and kompetitive allele-specific PCR (KASP) marker development with Korean japonica rice varieties. Plant Breed. Biotechnol. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Van Ooijen, J.W.J.H. LOD significance thresholds for QTL analysis in experimental populations of diploid species. Heredity 1999, 83, 613–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keb-Llanes, M.; González, G.; Chi-Manzanero, B.; Infante, D. A rapid and simple method for small-scale DNA extraction in Agavaceae and other tropical plants. Plant Mol. Biol. Rep. 2002, 20, 299–300. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Jackson, M. Hormones from roots as signals for the shoots of stressed plants. Trends Plant Sci. 1997, 2, 22–28. [Google Scholar] [CrossRef]
- Asano, T.; Wakayama, M.; Aoki, N.; Komatsu, S.; Ichikawa, H.; Hirochika, H.; Ohsugi, R. Overexpression of a calcium-dependent protein kinase gene enhances growth of rice under low-nitrogen conditions. Plant Biotechnol. 2010, 27, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Moudry, P.; Lukas, C.; Macurek, L.; Hanzlikova, H.; Hodny, Z.; Lukas, J.; Bartek, J. Ubiquitin-activating enzyme UBA1 is required for cellular response to DNA damage. Cell Cycle 2012, 11, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Maekawa, S.; Yasuda, S.; Yamaguchi, J. Carbon and nitrogen metabolism regulated by the ubiquitin-proteasome system. Plant Signal. Behav. 2011, 6, 1465–1468. [Google Scholar] [CrossRef] [Green Version]
- SZAL, B.; Podgorska, A. The role of mitochondria in leaf nitrogen metabolism. Plant Cell Environ. 2012, 35, 1756–1768. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, F.; Bao, A.; You, Q.; Li, Z.; Chen, J.; Cheng, Y.; Zhao, W.; Shen, X.; Zhou, X. The soybean Rhg1 amino acid transporter gene alters glutamate homeostasis and jasmonic acid-induced resistance to soybean cyst nematode. Mol. Plant Pathol. 2019, 20, 270–286. [Google Scholar] [CrossRef] [Green Version]
- Mittl, P.R.; Schneider-Brachert, W. Sel1-like repeat proteins in signal transduction. Cell. Signal. 2007, 19, 20–31. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Carrayol, E.; Valadier, M.-H. The two nitrogen mobilisation-and senescence-associated GS1 and GDH genes are controlled by C and N metabolites. Planta 2005, 221, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Feller, U.; Fischer, A. Nitrogen metabolism in senescing leaves. Crit. Rev. Plant Sci. 1994, 13, 241–273. [Google Scholar] [CrossRef]
- Fan, P.; Feng, J.; Jiang, P.; Chen, X.; Bao, H.; Nie, L.; Jiang, D.; Lv, S.; Kuang, T.; Li, Y. Coordination of carbon fixation and nitrogen metabolism in Salicornia europaea under salinity: Comparative proteomic analysis on chloroplast proteins. Proteomics 2011, 11, 4346–4367. [Google Scholar] [CrossRef] [PubMed]
- Herbik, A.; Koch, G.; Mock, H.P.; Dushkov, D.; Czihal, A.; Thielmann, J.; Stephan, U.; Bäumlein, H. Isolation, characterization and cDNA cloning of nicotianamine synthase from barley: A key enzyme for iron homeostasis in plants. Eur. J. Biochem. 1999, 265, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Hakoyama, T.; Watanabe, H.; Tomita, J.; Yamamoto, A.; Sato, S.; Mori, Y.; Kouchi, H.; Suganuma, N. Nicotianamine synthase specifically expressed in root nodules of Lotus japonicus. Planta 2009, 230, 309. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.; Hannam, C.; Gu, H.; Bi, Y.M.; Rothstein, S.J. A mutation in NLA, which encodes a RING-type ubiquitin ligase, disrupts the adaptability of Arabidopsis to nitrogen limitation. Plant J. 2007, 50, 320–337. [Google Scholar] [CrossRef]
- Podzimska-Sroka, D.; O’Shea, C.; Gregersen, P.L.; Skriver, K. NAC transcription factors in senescence: From molecular structure to function in crops. Plants 2015, 4, 412–448. [Google Scholar] [CrossRef] [Green Version]
- Sablowski, R.W.; Meyerowitz, E.M. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Harling, H.; Czaja, I.; Schell, J.; Walden, R. A plant cation–chloride co-transporter promoting auxin-independent tobacco protoplast division. EMBO J. 1997, 16, 5855–5866. [Google Scholar] [CrossRef] [Green Version]
- Walden, R.; Hayashi, H.; Lubenow, H.; Czaja, I.; Schell, J. Auxin inducibility and developmental expression of axi 1: A gene directing auxin independent growth in tobacco protoplasts. EMBO J. 1999, 18, 2908. [Google Scholar] [CrossRef]
- Commichau, F.M.; Forchhammer, K.; Stülke, J. Regulatory links between carbon and nitrogen metabolism. Curr. Opin. Microbiol. 2006, 9, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Burkovski, A. 14 Nitrogen Metabolism and Its Regulation. In Handbook of Corynebacterium glutamicum.; CRC Press: Boca Raton, FL, USA, 2005; p. 333. [Google Scholar]
- Zhang, J.; Liu, Y.-X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, Y.; Yan, P.; Zhang, X.; Guo, X. NRT1. 1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef]
- Sun, P.; Liu, F.; Tan, L.; Zhu, Z.; Fu, Y.; Sun, C.; Cai, H.J.I.J.G.P.B. Quantitative trait loci (QTLs) for potassium chlorate resistance and low temperature tolerance in seedling stage in rice (Oryza sativa L.). Indian J. Genet. Plant Breed. 2012, 72, 405–414. [Google Scholar]
- Wallsgrove, R.M.; Keys, A.J.; Lea, P.J.; Miflin, B.J. Photosynthesis, photorespiration and nitrogen metabolism. Plant Cell Environ. 1983, 6, 301–309. [Google Scholar]
- Steward, F.; Bidwell, R. Nitrogen metabolism, respiration, and growth of cultured plant tissue: PART IV. The impact of growth on protein metabolism and respiration of carrot tissue explants. General discussion of results. J. Exp. Bot. 1958, 285–305. [Google Scholar] [CrossRef]
- Weger, H.G.; Turpin, D.H. Mitochondrial respiration Can support NO3− and NO2− reduction during photosynthesis: Interactions between photosynthesis, respiration, and N assimilation in the N-limited green alga Selenastrum minutum. Plant Physiol. 1989, 89, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G.; Hodges, M. Respiration and nitrogen assimilation: Targeting mitochondria-associated metabolism as a means to enhance nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1467–1482. [Google Scholar] [CrossRef] [Green Version]
- Maier, R.J. Nitrogen fixation and respiration: Two processes linked by the energetic demands of Nitrogenase. In Respiration in Archaea and Bacteria; Springer: Berlin/Heidelberg, Germany, 2004; pp. 101–120. [Google Scholar]
- Huppe, H.; Turpin, D. Integration of carbon and nitrogen metabolism in plant and algal cells. Annu. Rev. Plant Biol. 1994, 45, 577–607. [Google Scholar] [CrossRef]
- Saïd, H.; El Shishiny, E. Respiration and nitrogen metabolism of whole and sliced radish roots with reference to the effect of alternation of air and nitrogen atmospheres. Plant Physiol. 1947, 22, 452. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | QTL (a) | Chr (b) | Position (cM) (c) | Left Marker (d) | Right Marker (e) | LOD (f) | PVE (%) (g) | Add (h) |
|---|---|---|---|---|---|---|---|---|
| Chlorate resistance | qCHR-3 | 3 | 136 | ah03001094 | id3005168 | 4.1 | 14.92 | −11.975 |
| MSU ID (Nipponbare) | Indica Database (Ensembl) | Description | Remarks | Similar Papers |
|---|---|---|---|---|
| LOC_Os03g18170 | BGIOSGA012400 | Protein kinase domain-containing protein, expressed | 162 bp deletion in indica | [43] |
| LOC_Os03g18380 | BGIOSGA010930 | Ubiquitin-activating enzyme E1 | 357 bp deletion in japonica | [44,45] |
| LOC_Os03g18500 | BGIOSGA010922 | Mitochondrial import inner membrane translocase subunit tim22 | 100% similar | [46] |
| LOC_Os03g18550 | BGIOSGA012419 | Mitochondrial carrier protein domain-containing protein | 100% similar | [46] |
| LOC_Os03g18630 | BGIOSGA012425 | Leucine-rich repeat family protein, receptor-like kinase RHG1, putative, expressed | 100% similar | [47] |
| LOC_Os03g18690 | BGIOSGA012432 | 26S protease regulatory subunit 4 homolog (26S proteasome subunit AtRPT2a) | 72 bp deletion in indica and 102 bp deletion in japonica | [45] |
| LOC_Os03g18700 | BGIOSGA010916 | Sel1-like domain containing protein (Sel1-like repeats); these represent a subfamily of TPR (tetratricopeptide repeat) sequences. | 306 bp deletion in indica, and 30 bp deletion in japonica | [48] |
| LOC_Os03g18790 | BGIOSGA010911 | Senescence-associated gene 20 (SAG20) putative, expressed. Cys-rich family protein (DUF2985) and PLAC8 family domains. This family includes the placenta-specific gene 8 protein. | 100% similar | [49] |
| LOC_Os03g19290 | BGIOSGA012456 | Mitochondrial import inner membrane translocase, subunit Tim17/22 family protein, putative, expressed | 27 bp deletion in japonica | [46] |
| LOC_Os03g19380 | BGIOSGA012463 | Calvin cycle protein CP12, putative, expressed. A chloroplast protein that regulates the Calvin cycle responsible for CO2 assimilation. | 100% similar | [50,51] |
| LOC_Os03g19420 | BGIOSGA010884 | Nicotianamine synthase 2, putative, expressed | 100% similar | [52,53] |
| LOC_Os03g19500 | BGIOSGA012473 | Ubiquitin-conjugating enzyme/RWD-like domain containing protein | 3 bp deletion in indica | [45] |
| LOC_Os03g20980 | BGIOSGA010820 | Zinc finger, RING-type domain-containing protein. Zinc finger, C3HC4 type domain-containing protein, expressed. | 51 bp deletion in indica and 3 bp deletion in japonica | [54] |
| LOC_Os03g21060 | BGIOSGA026407 | NAC domain-containing protein 29 (ANAC029) (NAC2) (NAC-LIKE, ACTIVATED BY AP3/PI protein) (NAP). No apical meristem protein, putative, expressed. | 156 bp deletion in indica and 12 bp deletion in japonica | [55,56,57] |
| LOC_Os03g21090 | BGIOSGA010814 | Axi1 auxin-independent growth promoter protein, putative, expressed | 100% similar | [58,59] |
| LOC_Os03g21490 | BGIOSGA010800 | ABC transporter, ATP-binding component. ABC transporter I family member 6, chloroplastic | 100% similar | [60,61] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabange, N.R.; Park, S.-Y.; Shin, D.; Lee, S.-M.; Jo, S.-M.; Kwon, Y.; Cha, J.-K.; Song, Y.-C.; Ko, J.-M.; Lee, J.-H. Identification of a Novel QTL for Chlorate Resistance in Rice (Oryza sativa L.). Agriculture 2020, 10, 360. https://doi.org/10.3390/agriculture10080360
Kabange NR, Park S-Y, Shin D, Lee S-M, Jo S-M, Kwon Y, Cha J-K, Song Y-C, Ko J-M, Lee J-H. Identification of a Novel QTL for Chlorate Resistance in Rice (Oryza sativa L.). Agriculture. 2020; 10(8):360. https://doi.org/10.3390/agriculture10080360
Chicago/Turabian StyleKabange, Nkulu Rolly, So-Yeon Park, Dongjin Shin, So-Myeong Lee, Su-Min Jo, Youngho Kwon, Jin-Kyung Cha, You-Chun Song, Jong-Min Ko, and Jong-Hee Lee. 2020. "Identification of a Novel QTL for Chlorate Resistance in Rice (Oryza sativa L.)" Agriculture 10, no. 8: 360. https://doi.org/10.3390/agriculture10080360