Agronomic Biofortification with Selenium in Tomato Crops (Solanum lycopersicon L. Mill)

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Samples Preparation
2.3. Agronomic Yield
2.4. Selenium Content and Nutritional Status
2.5. Statistical Data Analysis
3. Results
3.1. Yield of Agronomic Variables
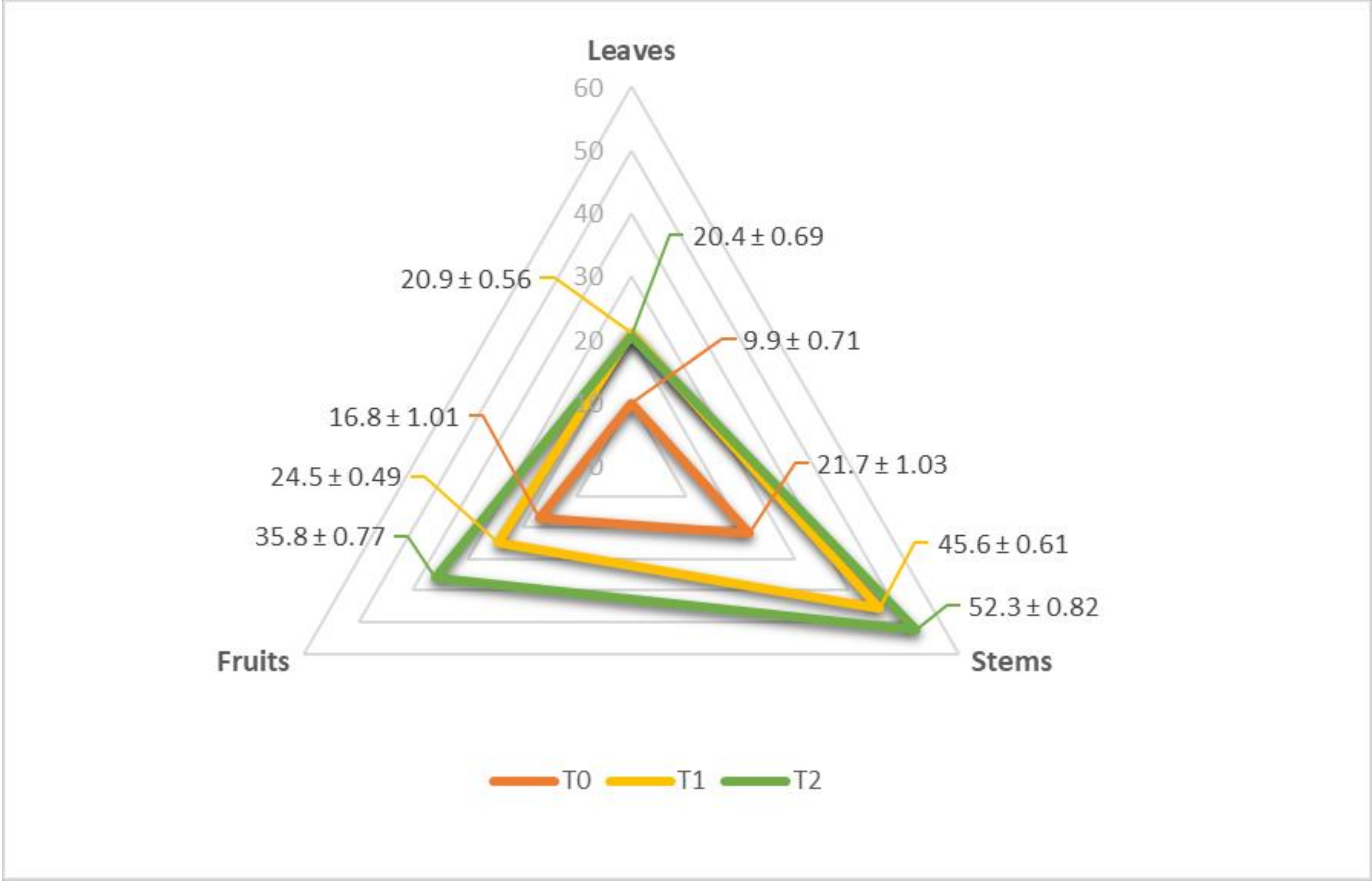
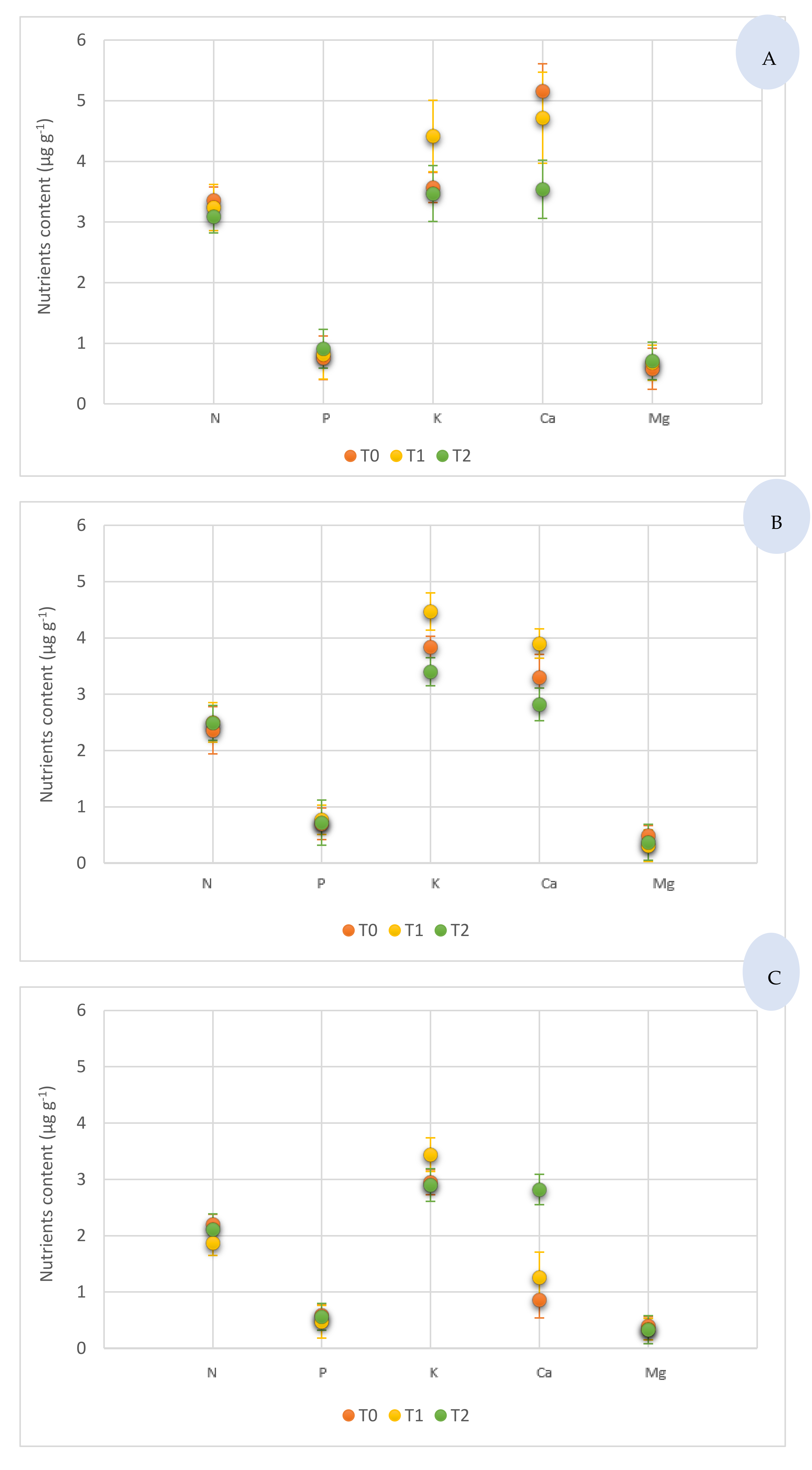
3.2. Selenium and Macronutrient Content in Different Tomato Components, under Sodium Selenite Treatments
4. Discussion
4.1. Agronomic Variables of Tomato under Sodium Selenite Treatments
4.2. Selenium Content in Different Components of Tomato under Sodium Selenite Treatments
4.3. Macronutrients Content from Different Components of Tomato under Sodium Selenite Treatments
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Sec. 2017, 12, 49–58. [Google Scholar] [CrossRef]
- FAO; IFAD; WFP. The State of Food Insecurity in the World; FAO: Rome, Italy, 2015. [Google Scholar]
- Bouis, H.E.; Hotz, C.; McClafferty, B.; Meenakshi, J.V.; Pfeiffer, W.H. Biofortification: A new tool to reduce micronutrient malnutrition. Food Nutr. Bull. 2011, 32 (Suppl. 1), S31–S40. [Google Scholar] [CrossRef] [PubMed]
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the Health-Promoting Effects of Tomato Fruit for Biofortified Food. Mediat. Inflamm. 2014, 2014, 139873. [Google Scholar] [CrossRef] [PubMed]
- Canene-Adams, K.; Campbell, J.K.; Zaripheh, S.; Jeffery, E.H.; Erdman, J.W., Jr. The tomato as a functional food. J. Nutr. 2005, 135, 1226–1230. [Google Scholar] [CrossRef]
- Frusciante, L.; Carli, P.; Ercolano, M.R. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Riso, P.; Grande, S.; Galli, C.; Porrini, M. Protective activity of tomato products on in vivo markers of lipid S. oxidation. Eur. J. Nutr. 2003, 42, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Periago, M.J.; García-Alonso, J.; Jacob, K.; Belén Olivares, A.; José Bernal, M.; Dolores Iniesta, M.; Martínez, C.; Ros, G. Bioactive compounds, folates and antioxid.ant properties of tomatoes (Lycopersicum esculentum) during vine ripening. Int. J. Food Sci. Nutr. 2009, 60, 694–708. [Google Scholar] [CrossRef]
- García-Valeverde, V.; Navarro-Gonzáles, I.; García-Alonso, J.; Periago, M.J. Antioxidant bioactive compounds in selected industrial processing and fresh consumption tomato cultivars. Food Bioproc. Tech. 2013, 6, 391–402. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V. Selenium and plants. Acta Agric. Slov. 2007, 89, 1. [Google Scholar] [CrossRef]
- Kuznetsov, V.; Kuznetsov, V. Selenium regulates the water status of plants exposed to drought. Dokl. Biol. Sci. 2003, 390, 266–268. [Google Scholar] [CrossRef]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth promoting effect of selenium on senescing lettuce. Plant Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Pennanen, A.; Xue, T.; Hartikainen, H. Protective role of selenium in plant subjected to severe UV irradiation stress. J. Appl. Bot. 2002, 76, 66–76. [Google Scholar]
- Schwarz, K.; Foltz, C.M. Selenium as an Integral Part of Factor 3 against Dietary Necrotic Liver Degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- Achibat, H.; AlOmari, N.A.; Messina, F.; Sancineto, L.; Khouili, M.; Santi, C. Organoselenium Compounds as Phytochemicals from the Natural Kingdom. Nat. Prod. Commun. 2015, 10, 1934578X1501001. [Google Scholar] [CrossRef] [Green Version]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Selenium influences glucosinolate and isothiocyanates and increases sulfur uptake in Arabidopsis thaliana and rapid-cycling Brassica oleracea. J. Agric. Food Chem. 2013, 61, 202–209. [Google Scholar] [CrossRef]
- Schiavon, M.; Dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.H.; Sambo, P.; Masi, A.; Malagoli, M. Selenium fertilization alters the chemical composition and antioxidant constituents of tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef]
- D’Amato, R.; De Feudis, M.; Guiducci, M.; Businelli, D. Zea mays L. Grain: Increase in Nutraceutical and Antioxidant Properties Due to Se Fortification in Low and High Water Regimes. J. Agric. Food Chem. 2019, 67, 7050–7059. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Tsuji, P.A.; Carlson, B.A.; Gladyshev, V.N. Selenium and selenocysteine: Roles in cancer, health and development. Trends Biochem. Sci. 2014, 39, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; Mc Grath, S.P.; Zhao, F.J.; Breward, N.; et al. Biofortification of UK food crops with selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Li, H.F.; Mc Grath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Feist, L.J.; Parker, D.R. Ecotypic variation in selenium accumulation among populations of Stanleya pinnata. New Phytol. 2001, 149, 61–69. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, G.; Chen, J.; Hu, Q. Uptake and transport of selenite and selenate by soybean seedlings of two genotypes. Plant Soil 2003, 253, 437–443. [Google Scholar] [CrossRef]
- Malagoli, M.; Schiavon, M.; dall’Acqua, S.; Pilon-Smits, E.A. Effects of selenium biofortification on crop nutritional quality. Front. Plant. Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, M.P.; Pilon-Smits, E.A.H.; Lytle, C.M.; Hwang, S.; Tai, J.; Honma, T.S.U.; Yeh, L.; Terry, N. Rate-limiting steps in selenium assimilation and volatilization by Indian mustard. Plant Physiol. 1998, 117, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Cartes, P.; Shene, C.; Mora, M.L. Selenium distribution in ryegrass and its antioxidant role as affected by sulfur fertilization. Plant Soil. 2006, 285, 187–195. [Google Scholar] [CrossRef]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Cartes, P.; Gianfrera, L.; Mora, M.L. Uptake of selenium and its antioxidative activity in ryegrass when applied a selenate and selenite forms. Plant Soil 2005, 276, 359–367. [Google Scholar] [CrossRef] [Green Version]
- García, E. Modificaciones al Sistema de Clasificación Climática de Koppen Para Adaptarlo a las Condiciones de la República Mexicana, 3rd ed.; UNAM: Mexico City, Mexico, 2004; p. 252. [Google Scholar]
- Preciado-Rangel, P.; Favela-Chavez, E.; Benavides-Mendoza, A. Manual para la Preparación de Soluciones Nutritivas; Universidad Autónoma Agraria Antonio Narro: Saltillo, Mexico, 2006; p. 146. [Google Scholar]
- A.O.A.C. Official Methods of Analysis of the Association of Official Analytical Chemists, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000; p. 2000. [Google Scholar]
- A.O.A.C. Official Methods of Analysis of the Association of Official Analytical Chemist, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990; pp. 829–830. [Google Scholar]
- Harris, W.D.; Popat, P. Determination of the Phosphorus Content of Lipids. J. Am. Oil Chem. Soc. 1954, 31, 124–127. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Modulation of antioxidant machinery and the methylglyoxal detoxification system in selenium-supplemented Brassica napus seedlings confers tolerance to high temperature stress. Biol. Trace Elem. Res. 2014, 161, 297–307. [Google Scholar] [CrossRef]
- Yildiztugay, E.; Ozfidan-Konakci, C.; Kucukoduk, M.; Tekis, S.A. The impact of selenium application on enzymatic and non-enzymatic antioxidant systems in Zea mays roots treated with combined osmotic and heat stress. Arch. Agron. Soil Sci. 2017, 63, 261–275. [Google Scholar] [CrossRef]
- Sattar, A.; Cheema, M.A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Nawaz, A.; Abbas, T.; Ali, Q. Physiological and biochemical attributes of bread wheat (Triticum aestivum L.) seedlings are influenced by foliar application of silicon and selenium under water deficit. Acta Physiol. Plant. 2019, 41, 146. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I.; Iqbal, M. Phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Dresler, S.; Wójcik, M. Selenium affects physiological parameters and phytochelatins accumulation in cucumber (Cucumis sativus L.) plants grown under cadmium exposure. Sci. Hort. 2014, 172, 10–18. [Google Scholar] [CrossRef]
- Huang, C.; Qin, N.; Sun, L.; Yu, M.; Hu, W.; Qi, Z. Selenium improves physiological parameters and alleviates oxidative stress in strawberry seedlings under low temperature stress. Int. J. Mol. Sci. 2018, 19, 1913. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, R.; Awasthi, S.; Tripathi, P.; Mishra, S.; Dwivedi, S.; Niranjan, A.; Mallick, S.; Tripathi, P.; Pande, V.; Tripathi, R.D. Selenite modulates the level of phenolics and nutrient element to alleviate the toxicity of arsenite in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2017, 138, 47–55. [Google Scholar] [CrossRef]
- Mroczek-Zdyrska, M.; Wójcik, M. The influence of selenium on root growth and oxidative stress induced by lead in Vicia faba L. minor plants. Biol. Trace Elem. Res. 2012, 147, 320–328. [Google Scholar] [CrossRef]
- Golob, A.; Kavčič, J.; Stibilj, V.; Gaberščik, A.; Vogel-Mikuš, K.; Germ, M. The effect of selenium and UV radiation on leaf traits and biomass production in Triticum aestivum L. Ecotox. Environ. Saf. 2017, 136, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Mata-Ramírez, D.; Serna-Saldívar, S.O.; Antunes-Ricardo, M. Enhancement of antiinflammatory and antioxidant metabolites in soybean (Glycine max) calluses subjected to selenium or UV-light stresses. Sci. Hortic. 2019, 257, 108669. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron. Soil Sci. 2014, 60, 1593–1624. [Google Scholar] [CrossRef]
- Balk, J.; Pilon, M. Ancient and essential: The assembly of iron-sulfur clusters in plants. Trends Plant Sci. 2011, 16, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Mroczek-Zdyrska, M.; Strubińska, J.; Hanaka, A. Selenium improves physiological parameters and alleviates oxidative stress in shoots of lead-exposed Vicia faba L. minor plants grown under phosphorus-deficient conditions. J. Plant Growth Reg. 2017, 36, 186–199. [Google Scholar] [CrossRef] [Green Version]
- Molnár, Á.; Kolbert, Z.; Kéri, K.; Feigl, G.; Ördög, G.; Szöllösi, R.; Erdei, L. Selenite induced nitro-oxidative stress processes in Arabidopsis thaliana and Brassica juncea. Ecotoxicol. Environ. Saf. 2018, 148, 664–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Kreft, I.; Osvald, J. Influence of UV-B Exclusion and Selenium Treatment on Photochemical Efficiency of Photosystem II, Yield and Respiratory Potential in Pumpkins (Cucurbita pepo L.). Plant Physiol. Bioch. 2005, 43, 445–448. [Google Scholar] [CrossRef]
- Becvort-Azcurra, A.; Fuentes-Lara, L.O.; Benavides-Mendoza, A.; Ramírez, H.; Robledo-Torres, V.; Rodríguez-Mendoza, M.D.L.N. Aplicación de selenio en tomate: Crecimiento, productividad y estado antioxidante del fruto. Terra Latinoam. 2012, 30, 291–301. [Google Scholar]
- Nancy, D.; Arulselvi, P.I. Effect of selenium fortification on biochemical activities of tomato (Solanum lycopersicum) plants. Indo Am. J. Pharm. 2014, 4, 3997–4005. [Google Scholar]
- Nawaz, F.; Ashraf, M.; Ahmad, R.; Waraich, E.A.; Shabbir, R.N.; Bukhari, M.A. Supplemental Selenium Improves Wheat Grain Yield and Quality Through Alterations in Biochemical Processes Under Normal and Water Deficit Conditions. Food Chem. 2014, 175, 350–357. [Google Scholar] [CrossRef]
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Boldrin, K.V.F.; Avila, F.W.; Guilherme, L.R.G. Soil and Foliar Application of Selenium in Rice Biofortification. J. Food Compos. Anal. 2013, 31, 238–244. [Google Scholar] [CrossRef]
- Robbins, R.J.; Keck, A.S.; Banuelos, G.; Finley, J.W. Cultivation conditions and selenium fertilization alter the phenolic profile, glucosinolate and sulforaphane content of broccoli. J. Med. Food 2005, 8, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants; CRC Press Inc.: Boca Raton, FL, USA, 2001. [Google Scholar]
- Lyons, G. Biofortification of Cereals with Foliar Selenium and Iodine Could Reduce Hypothyroidism. Front. Plant Sci. 2018, 9, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilimba, A.; Young, S.; Black, C.; Rogerson, K.; Ander, E.; Watts, M.; Lammel, J.; Broadley, M.R. Maize grain and soil surveys reveal suboptimal dietary selenium intake is widespread in Malawi. Sci. Rep. 2011, 1, 72. [Google Scholar] [CrossRef] [PubMed]
- Winkel, L.; Johnson, C.; Lenz, M.; Grundl, T.; Leupin, O.; Amini, M.; Charlet, L. Environmental selenium research: From microscopic processes to global understanding. Environ. Sci. Technol. 2012, 46, 571–579. [Google Scholar] [CrossRef]
- Yang, G.Q.; Wang, S.Z.; Zhou, R.H.; Sun, S.Z. Endemic selenium intoxication of humans in china. Am. J. Clin. Nutr. 1983, 37, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antiox. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Assoc. Inc.: Sunderland, UK, 2005. [Google Scholar]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant. Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Lester, G.E.; Jifon, J.L.; Makus, D.J. Impact of potassium nutrition on postharvest fruit quality: Melon (Cucumis melo L) case study. Plant. Soil. 2010, 335, 117–131. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Sady, W. Assessment of Biofortification with Iodine and Selenium of Lettuce Cultivated in the NFT Hydroponic System. Sci. Hort. 2014, 166, 9–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Stems | Fruits | Dry Matter | |||
|---|---|---|---|---|---|---|
| H (cm) | D (mm) | N (Fr/plant) | W (g) | L (g) | S (g) | |
| 0 mg L−1 | 61.1 ± 3.9 b | 11.8 ± 0.9 b | 12.00 ± 2.9 a | 869.20 ± 224.6 a | 43.7 ± 14.5 b | 26.6 ± 3.0 a |
| 2 mg L−1 | 67.5 ± 3.0 a | 13.4 ± 0.8 a | 17.67 ± 3.9 a | 1061.53 ± 349.8 a | 48.1 ± 19.1 ab | 28.3 ± 3.6 a |
| 5 mg L−1 | 65.2 ± 2.9 a | 13.3 ± 0.8 a | 16.67 ± 3.6 a | 1184.83 ± 378.9 a | 50.9 ± 19.2 a | 29.9 ± 6.5 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahim, F.P.; Rocio, C.G.; Adalberto, B.M.; Lidia Rosaura, S.C.; Maginot, N.H. Agronomic Biofortification with Selenium in Tomato Crops (Solanum lycopersicon L. Mill). Agriculture 2020, 10, 486. https://doi.org/10.3390/agriculture10100486
Rahim FP, Rocio CG, Adalberto BM, Lidia Rosaura SC, Maginot NH. Agronomic Biofortification with Selenium in Tomato Crops (Solanum lycopersicon L. Mill). Agriculture. 2020; 10(10):486. https://doi.org/10.3390/agriculture10100486
Chicago/Turabian StyleRahim, Foroughbakhch Pournavab, Castillo Godina Rocio, Benavides Mendoza Adalberto, Salas Cruz Lidia Rosaura, and Ngangyo Heya Maginot. 2020. "Agronomic Biofortification with Selenium in Tomato Crops (Solanum lycopersicon L. Mill)" Agriculture 10, no. 10: 486. https://doi.org/10.3390/agriculture10100486