
Role of Epithelial-to-Mesenchymal Transition of Retinal Pigment Epithelial Cells in Glaucoma Cupping

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Culture on Stiffened Substrates
2.3. Cyclical Cell Stretching
2.4. RPE65 Immunofluorescence
2.5. RNA Extraction, cDNA Synthesis, and Quantitative Polymerase Chain Reaction (PCR)
2.6. Real-Time RT-PCR
2.7. Cell-Viability/Crystal-Violet Assay
2.8. Cell Migration
2.9. Cell Proliferation
2.10. Statistical Analysis
3. Results
3.1. RPE65 Is Markedly Expressed throughout the Lamina-Cribrosa Region in Glaucoma
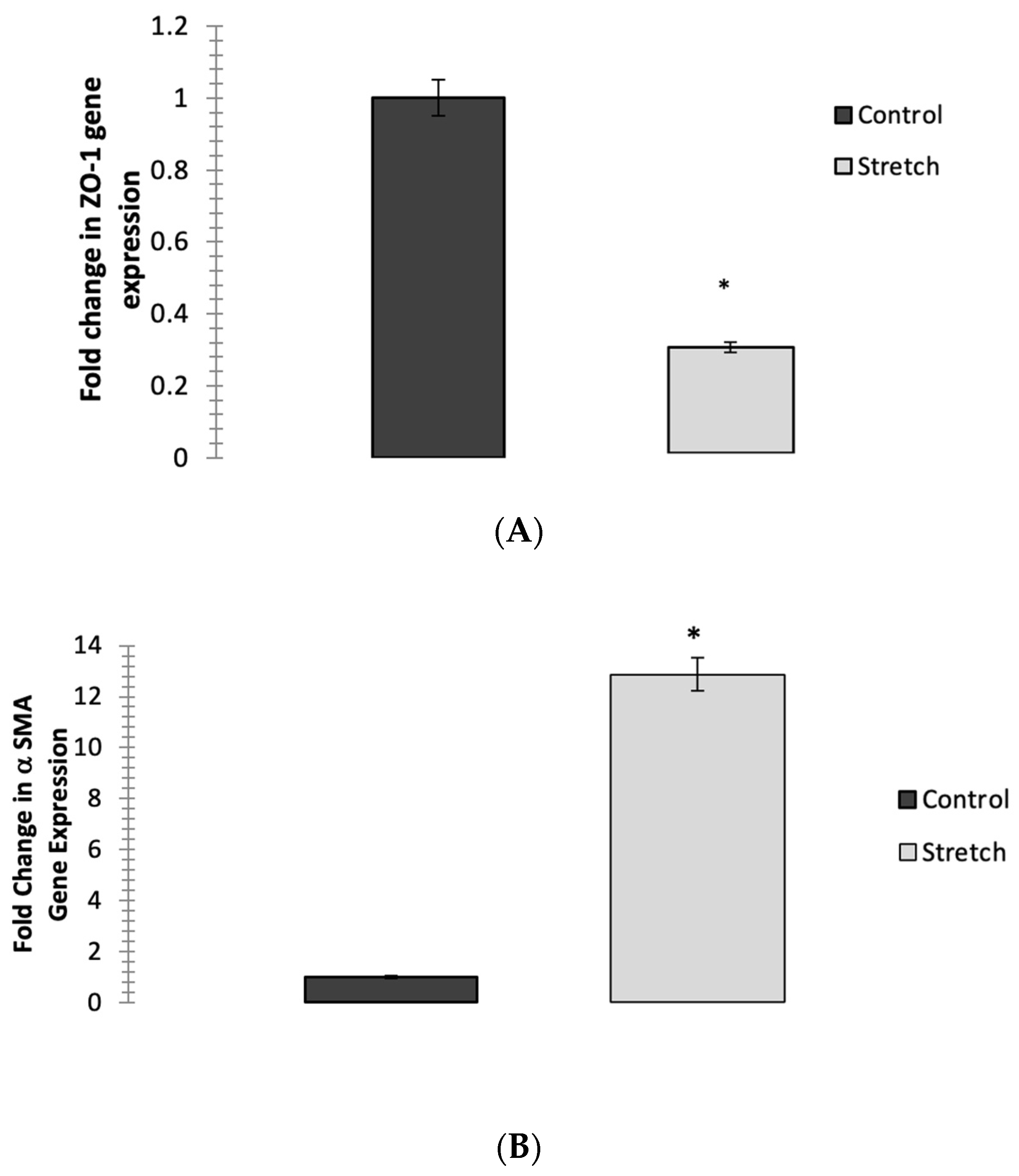
3.2. Cyclic Mechanical Stretching Reduces ZO-1 Expression and Increases αSMA-Gene Expression in ARPE-19 Cells Post-Stretching
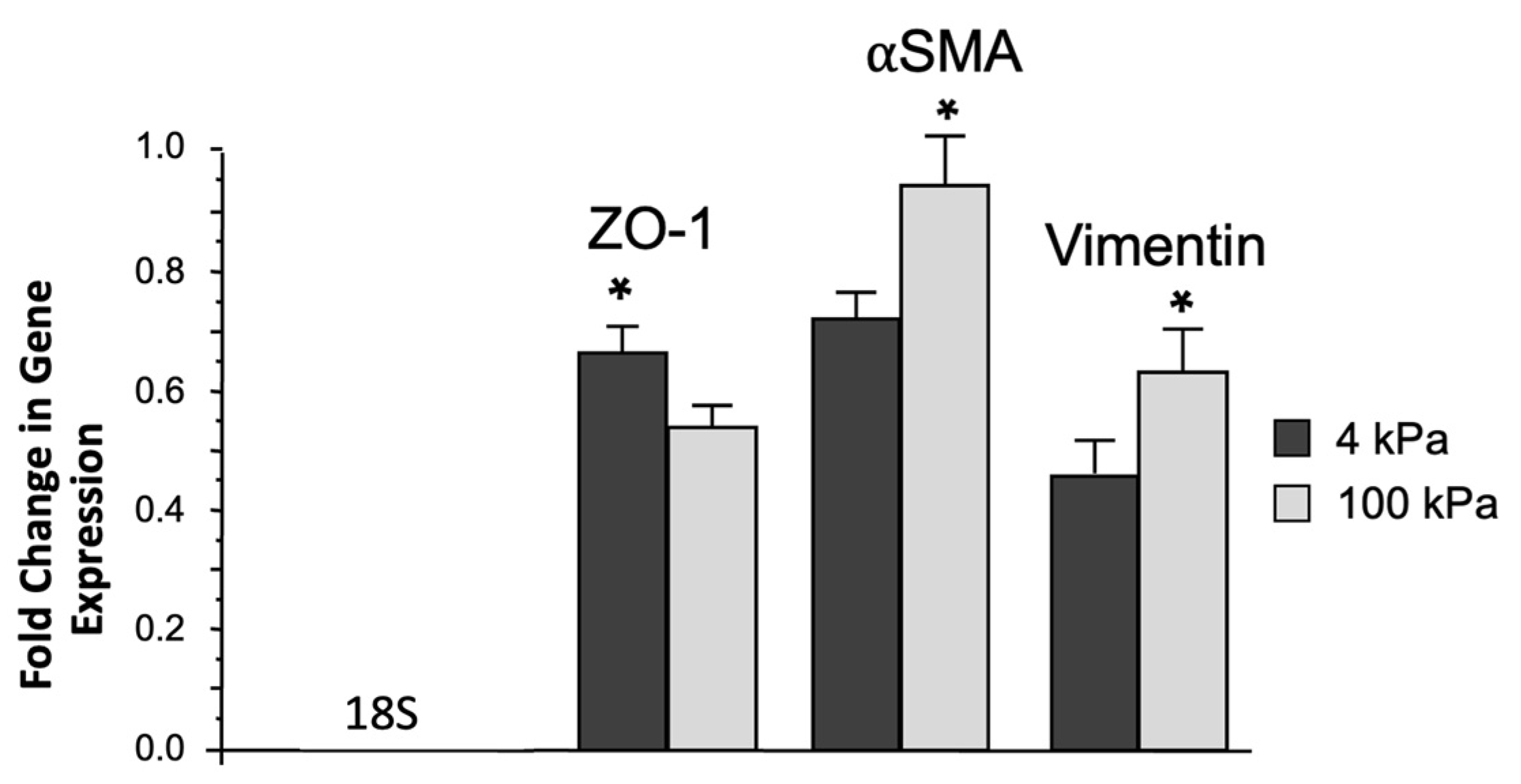
3.3. Increased Biomechanical Stiffness Reduces ZO-1-Gene Expression and Increases Vimentin and αSMA Gene Expression in ARPE-19 Cells
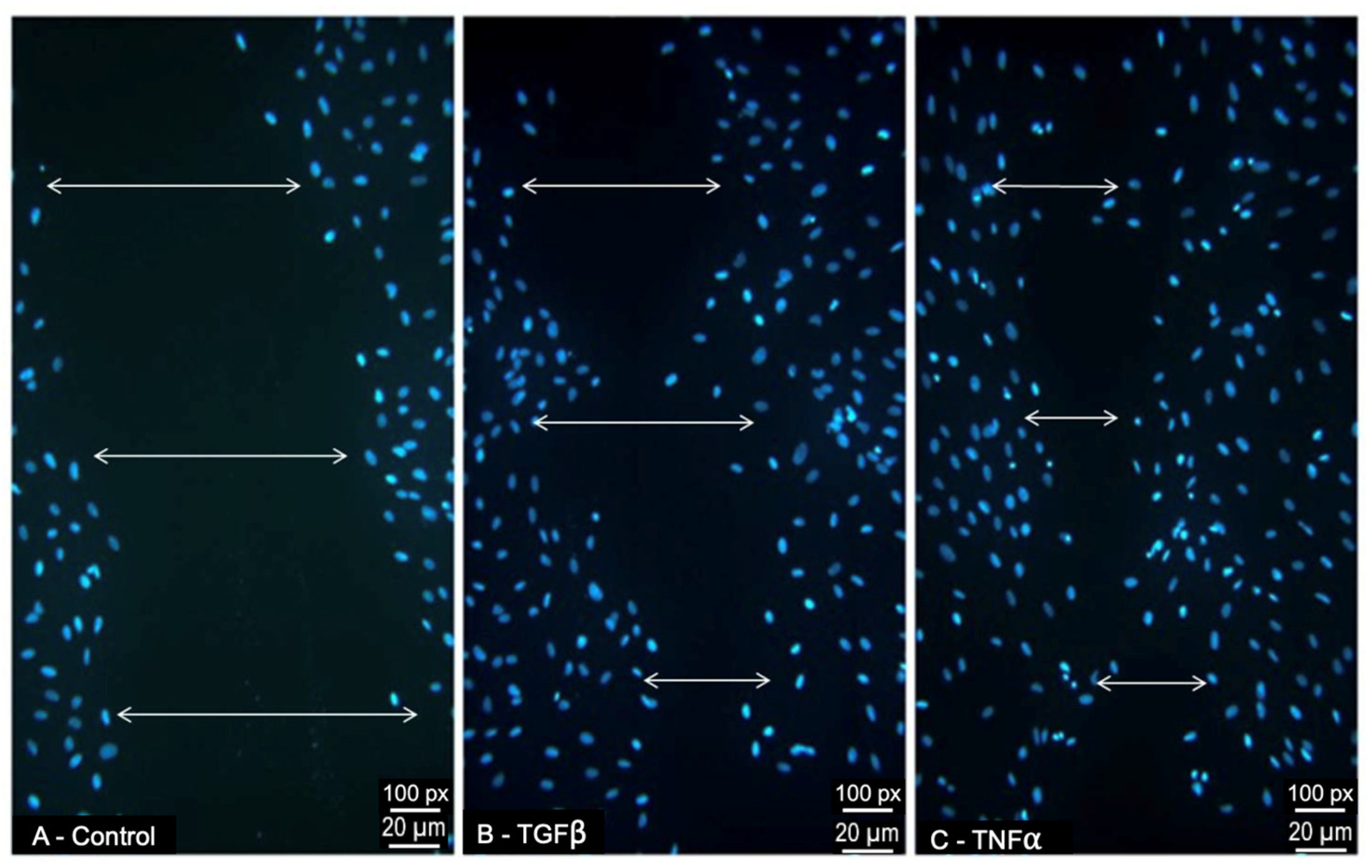
3.4. Effect of TGFβ1 and TNFα on ARPE-19-Cell Migratory Response
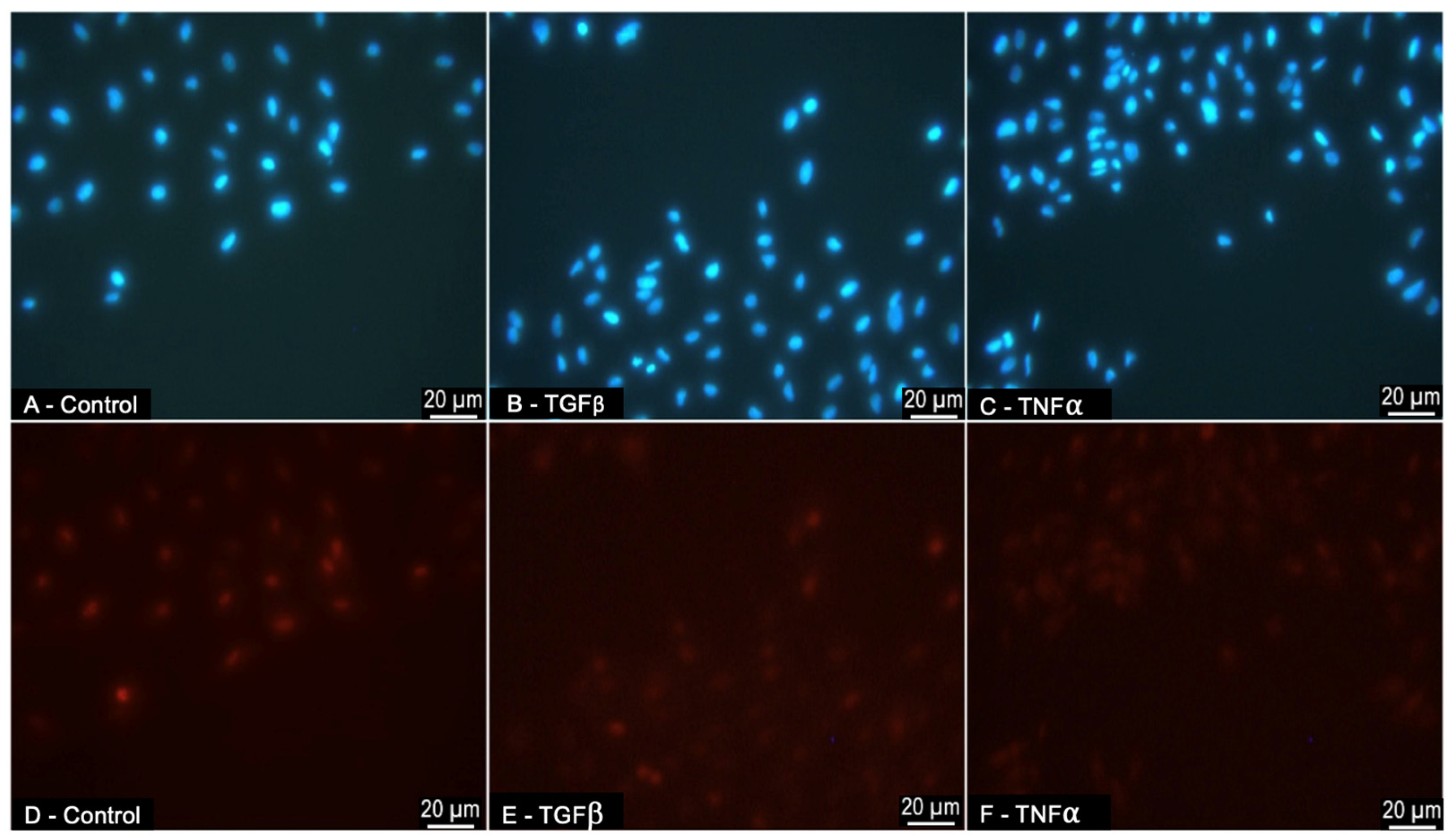
3.5. ARPE-19-Cell Proliferation in Response to Treatment with TGFβ1 and TNFα during Scratch Migration Assay
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kubota, T.; Jonas, J.B.; Naumann, G.O. Direct clinico-histological correlation of parapapillary chorioretinal atrophy. Br. J. Ophthalmol. 1993, 77, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.C.; De Moraes, C.G.V.; Prata, T.S.; Liebmann, C.A.; Tello, C.; Ritch, R.; Liebmann, J.M. The Region of Largest beta-Zone Parapapillary Atrophy Area Predicts the Location of Most Rapid Visual Field Progression. Ophthalmology 2011, 51, 2979. [Google Scholar]
- Jonas, J.B. Clinical implications of peripapillary atrophy in glaucoma. Curr. Opin. Ophthalmol. 2005, 16, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Manalastas, P.I.C.; Belghith, A.; Weinreb, R.N.; Jonas, J.B.; Suh, M.H.; Yarmohammadi, A.; Medeiros, F.A.; Girkin, C.A.; Liebmann, J.M.; Zangwill, L.M. Automated Beta Zone Parapapillary Area Measurement to Differentiate Between Healthy and Glaucoma Eyes. Am. J. Ophthalmol. 2018, 191, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Lee, E.J.; Kim, T.-W.; Kim, M.; Kim, H. Microstructure of β-zone parapapillary atrophy and rate of retinal nerve fiber layer thinning in primary open-angle glaucoma. Ophthalmology 2014, 121, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A.; Hohman, R.M.; Addicks, E.M.; Massof, R.W.; Green, W.R. Morphologic changes in the lamina cribrosa correlated with neural loss in open-angle glaucoma. Am. J. Ophthalmol. 1983, 95, 673–691. [Google Scholar] [CrossRef]
- Hernandez, M.R.; Igoe, F.; Neufeld, A.H. Extracellular matrix of the human optic nerve head. Am. J. Ophthalmol. 1986, 102, 139–148. [Google Scholar] [CrossRef]
- Hernandez, M.R.; Miao, H.; Lukas, T. Astrocytes in glaucomatous optic neuropathy. Prog. Brain Res. 2008, 173, 353–373. [Google Scholar]
- Kirwan, R.P.; Wordinger, R.J.; Clark, A.F.; O’Brien, C.J. Differential global and extra-cellular matrix focused gene expression patterns between normal and glaucomatous human lamina cribrosa cells. Mol. Vis. 2009, 15, 76–88. [Google Scholar]
- Pena JD, O.; Taylor, A.W.; Ricard, C.S.; Vidal, I.; Hernandez, M.R. Transforming growth factor beta isoforms in human optic nerve heads. Br. J. Ophthalmol. 1999, 83, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G.; Li, L.Y.; Patil, R.V.; Wax, M.B. TNF-alpha and TNF-alpha receptor-1 in the retina of normal and glaucomatous eyes. Investig. Opthalmol. Vis. Sci. 2001, 42, 1787–1794. [Google Scholar]
- Kirwan, R.P.; Leonard, M.O.; Murphy, M.; Clark, A.F.; O’Brien, C.J. Transforming growth factor-beta-regulated gene transcription and protein expression in human GFAP-negative lamina cribrosa cells. Glia 2005, 52, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Quill, B.; Docherty, N.G.; Clark, A.F.; O’Brien, C.J. The effect of graded cyclic stretching on extracellular matrix-related gene expression profiles in cultured primary human lamina cribrosa cells. Investig. Opthalmol. Vis. Sci. 2011, 52, 1908–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, R.S.; Dharsee, M.; Ackloo, S.; Sivak, J.; Flanagan, J.G. Proteomics analyses of human optic nerve head astrocytes following biomechanical strain. Mol. Cell Proteom. 2012, 11, M111.012302. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Benitez, N.E.; Parotto, M.; Post, M.; Han, B.; Spieth, P.M.; Cheng, W.-E.; Valladares, F.; Villar, J.; Liu, M.; Sato, M.; et al. Mechanical stress induces lung fibrosis by epithelial-mesenchymal transition. Crit. Care Med. 2012, 40, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y. Dissection of key events in tubular epithelial to myofibroblast transition and its implications in renal interstitial fibrosis. Am. J. Pathol. 2001, 159, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, Q.; Davis, F.; Mao, J.; Zhao, H.; Ma, D. Epithelial–mesenchymal transition in organ fibrosis development: Current understanding and treatment strategies. Burn. Trauma 2022, 10, tkac011. [Google Scholar] [CrossRef]
- Casaroli-Marano, R.P.; Pagan, R.; Vilaró, S. Epithelial-mesenchymal transition in proliferative vitreoretinopathy: Intermediate filament protein expression in retinal pigment epithelial cells. Investig. Opthalmol. Vis. Sci. 1999, 40, 2062–2072. [Google Scholar]
- Lee, H.; O’Meara, S.J.; O’Brien, C.; Kane, R. The role of gremlin, a BMP antagonist, and epithelial-to-mesenchymal transition in proliferative vitreoretinopathy. Investig. Opthalmol. Vis. Sci. 2007, 48, 4291–4299. [Google Scholar] [CrossRef]
- Lamouille, L.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nature reviews. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar]
- Yan, C.; Grimm, W.A.; Garner, W.L.; Qin, L.; Travis, T.; Tan, N.; Han, Y.-P. Epithelial to mesenchymal transition in human skin wound healing is induced by tumor necrosis factor-alpha through bone morphogenic protein-2. Am. J. Pathol. 2010, 176, 2247–2258. [Google Scholar] [CrossRef] [Green Version]
- Marinez, E.F.; Araujo, V.C.; Sousa, S.C.O.M.; Victor Elias, A.C. TGF-beta1 enhances the expression of alpha-smooth muscle actin in cultured human pulpal fibroblasts: Immunochemical and ultrastructural analyses. J. Endod. 2007, 33, 1313–1318. [Google Scholar] [CrossRef]
- Desmoulière, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- Fronk, A.H.; Vargis, E. Methods for culturing retinal pigment epithelial cells: A review of current protocols and future recommendations. J. Tissue Eng. 2016, 12, 2041731416650838. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Kohyama, T.; Takizawa, H.; Kamitani, S.; Desaki, M.; Takami, K.; Kawasaki, S.; Kato, J.; Nagase, T. Tumor necrosis factor-alpha enhances both epithelial-mesenchymal transition and cell contraction induced in A549 human alveolar epithelial cells by transforming growth factor-beta1. Exp. Lung Res. 2010, 36, 12–24. [Google Scholar] [CrossRef]
- Chen, H.-C.; Zhu, Y.-T.; Chen, S.-Y.; Tseng, S.C. Wnt signaling induces epithelial-mesenchymal transition with proliferation in ARPE-19 cells upon loss of contact inhibition. Lab. Investig. 2012, 92, 676–687. [Google Scholar] [CrossRef] [Green Version]
- Okinami, S.; Ohkuma, M.; Tsukahara, I. Kuhnt intermediary tissue as a barrier between the optic nerve and retina. Albrecht Von Graefes Arch. Für Klin. Und Exp. Ophthalmol. 1976, 201, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.A.; Perkumas, K.M.; Campbell, M.; Farrar, G.J.; Stamer, W.D.; Humphries, P.; O’Callaghan, J.; O’Brien, C.J. Fibrotic Changes to Schlemm’s Canal Endothelial Cells in Glaucoma. Int. J. Mol. Sci. 2021, 22, 9446. [Google Scholar] [CrossRef]
- Takahashi, E.; Inoue, T.; Fujimoto, T.; Kojima, S.; Tanihara, H. Epithelial mesenchymal transition-like phenomenon in trabecular meshwork cells. Exp. Eye Res. 2014, 118, 72–79. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward | Reverse |
|---|---|---|
| 18S | 5′-GTAACCCGTTGAACCCCATT | 5′-CCATCCAATCGGTAGTAGCC |
| ZO-1 | 5′-CCTCTTCCTGATGGATGGGAAC | 5′-TATTCCGCATTGCCTGCCG |
| Vimentin | 5′-TTCTGTACGCAGGTGATTGG | 5′-CATGTTCAGCTTTGTGGACC |
| αSMA | 5′-AAAGCTTCCCAGACTTCCGC | 5′-TTCTTGGGCCTTGATGCGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Driscoll, E.; Hughes, E.; Irnaten, M.; Kuehn, M.; Wallace, D.; O’Brien, C. Role of Epithelial-to-Mesenchymal Transition of Retinal Pigment Epithelial Cells in Glaucoma Cupping. J. Clin. Med. 2023, 12, 2737. https://doi.org/10.3390/jcm12072737
O’Driscoll E, Hughes E, Irnaten M, Kuehn M, Wallace D, O’Brien C. Role of Epithelial-to-Mesenchymal Transition of Retinal Pigment Epithelial Cells in Glaucoma Cupping. Journal of Clinical Medicine. 2023; 12(7):2737. https://doi.org/10.3390/jcm12072737
Chicago/Turabian StyleO’Driscoll, Eabha, Emily Hughes, Mustapha Irnaten, Markus Kuehn, Deborah Wallace, and Colm O’Brien. 2023. "Role of Epithelial-to-Mesenchymal Transition of Retinal Pigment Epithelial Cells in Glaucoma Cupping" Journal of Clinical Medicine 12, no. 7: 2737. https://doi.org/10.3390/jcm12072737