
Neonatal Hepatic Myeloid Progenitors Expand and Propagate Liver Injury in Mice
 , and
, and 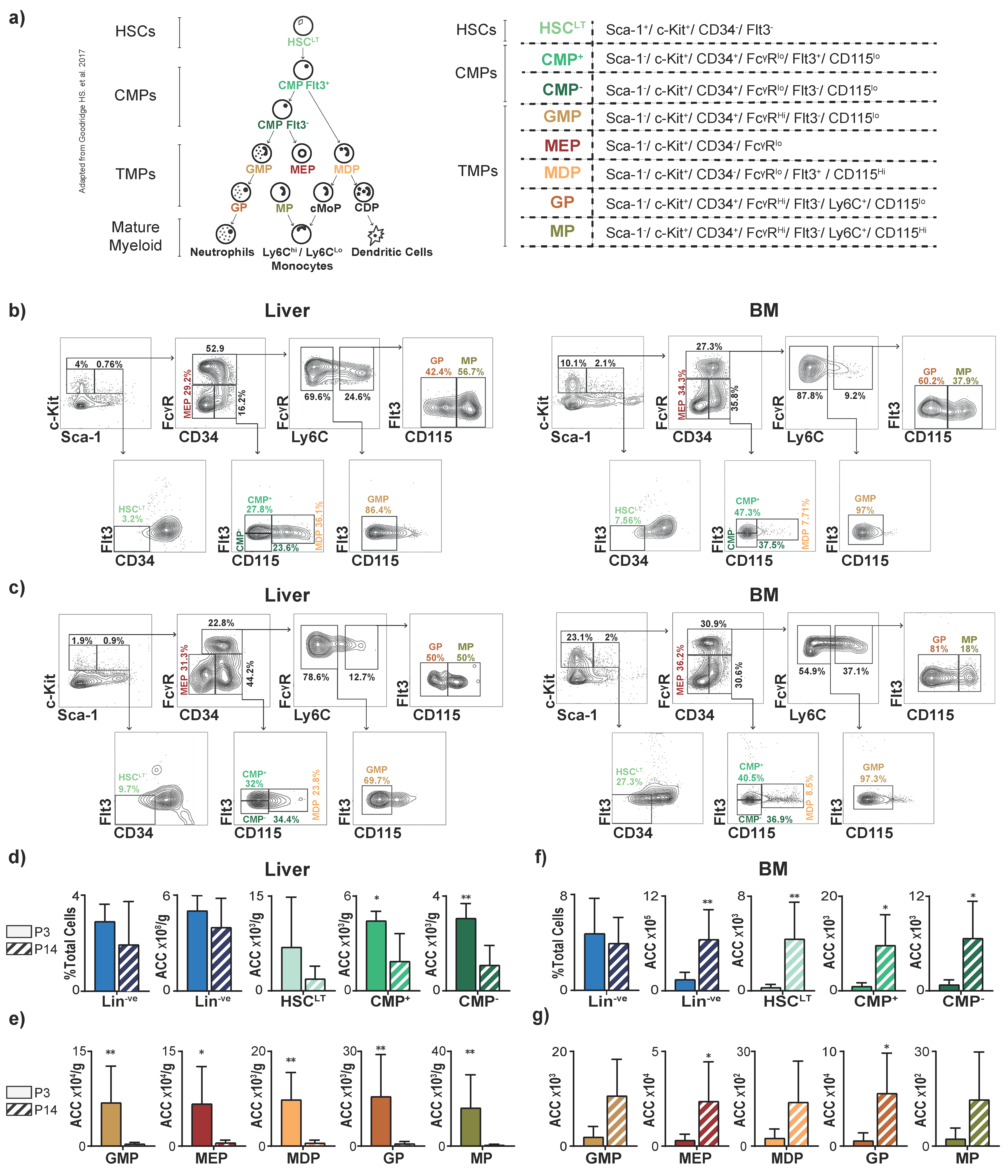{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Creation of Single-Cell Suspensions
2.3. Flow Cytometry
2.4. Colony-Forming Unit (CFU) Assays
2.5. Postnatal Model of Perinatal Liver Injury
2.6. Histologic Analysis
2.7. Antibody-Mediated Myeloablation in Neonatal Mice
2.8. Data Analysis
3. Results
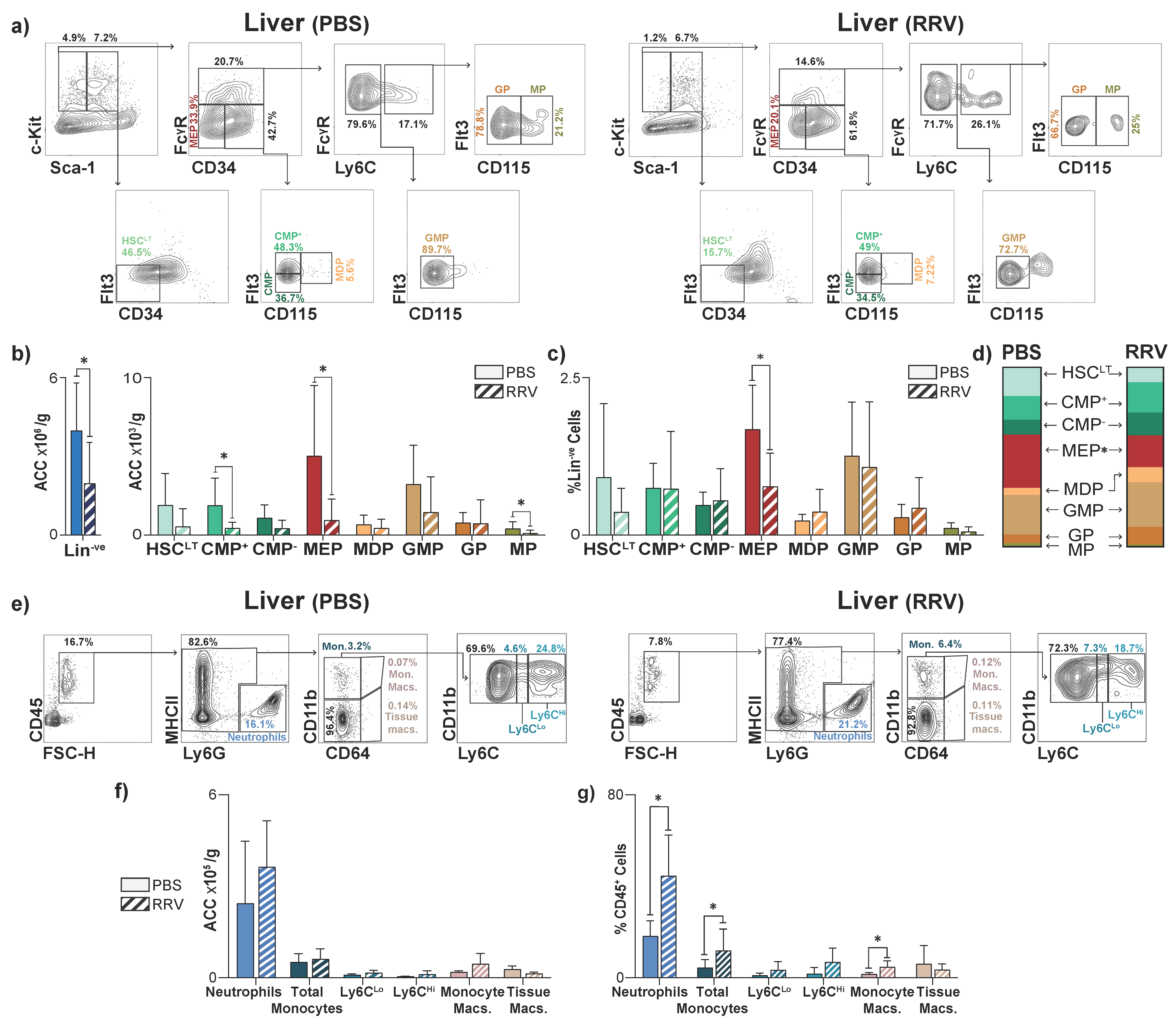
3.1. The Liver Is a Reservoir for Hematopoietic Progenitors in Neonatal Mice
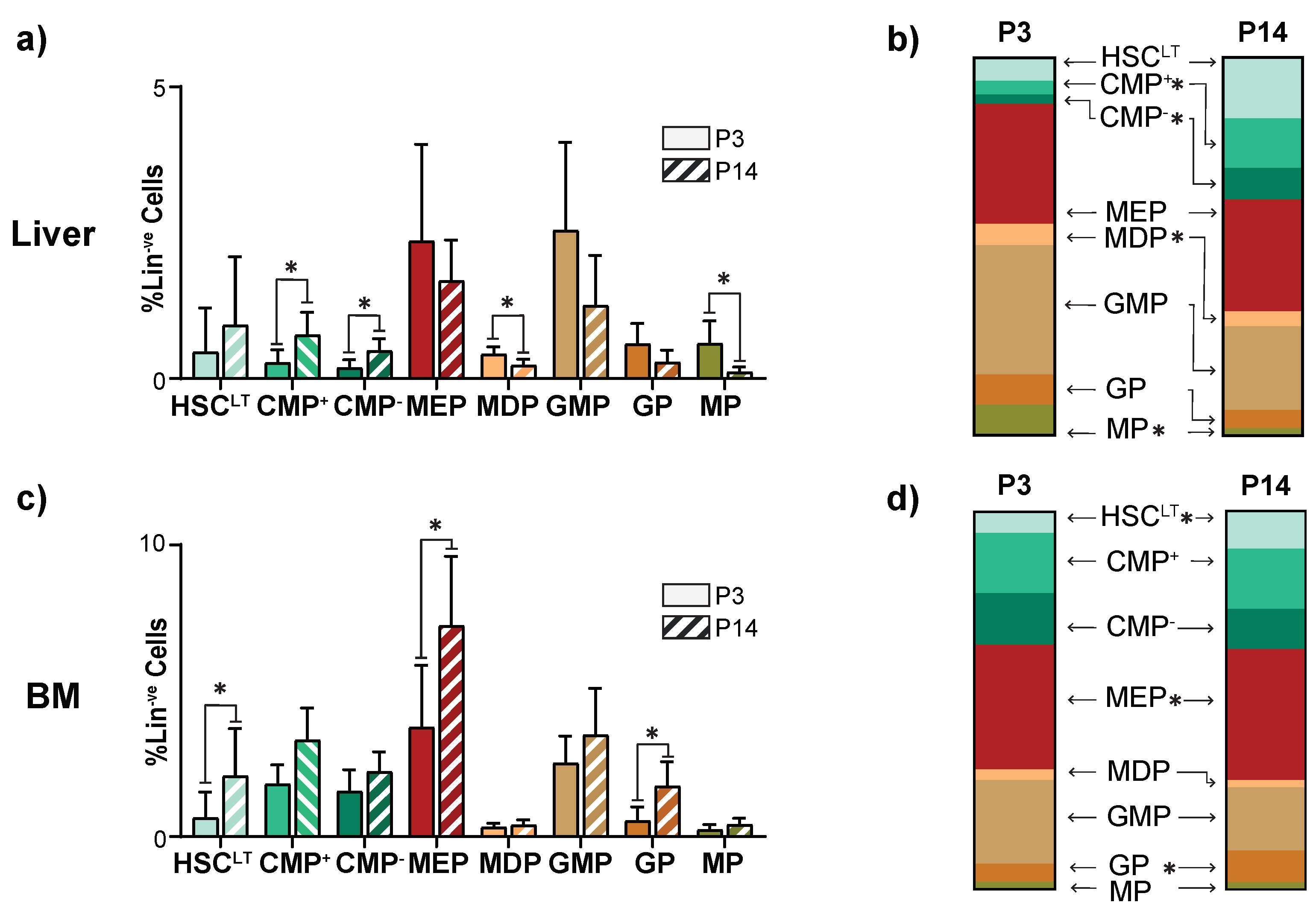
3.2. The Juvenile Liver Retains Common Myeloid Progenitors and Myeloid Differentiation Capacity
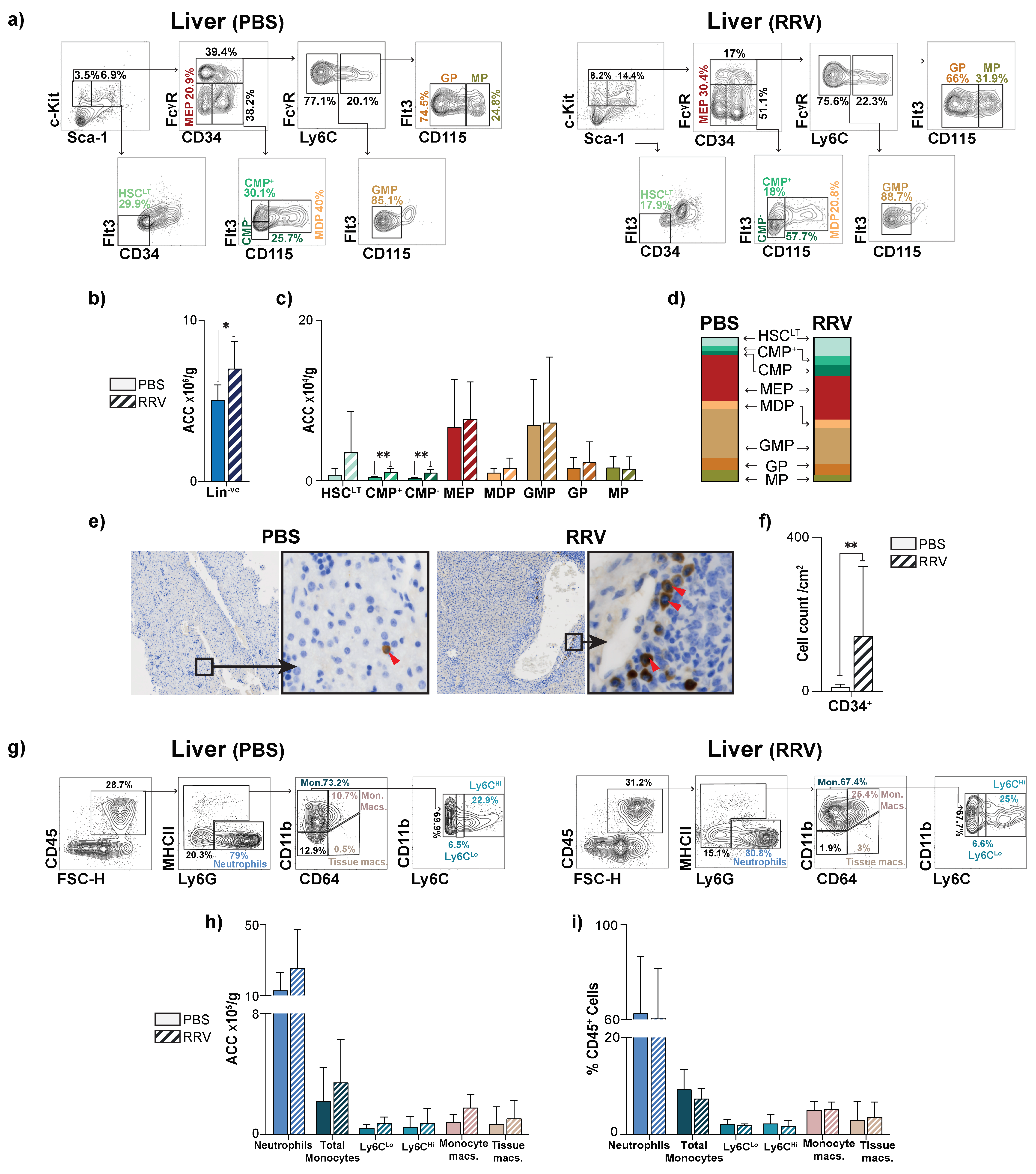
3.3. Perinatal Liver Injury Leads to Expansion of CMPs in the Neonatal Liver
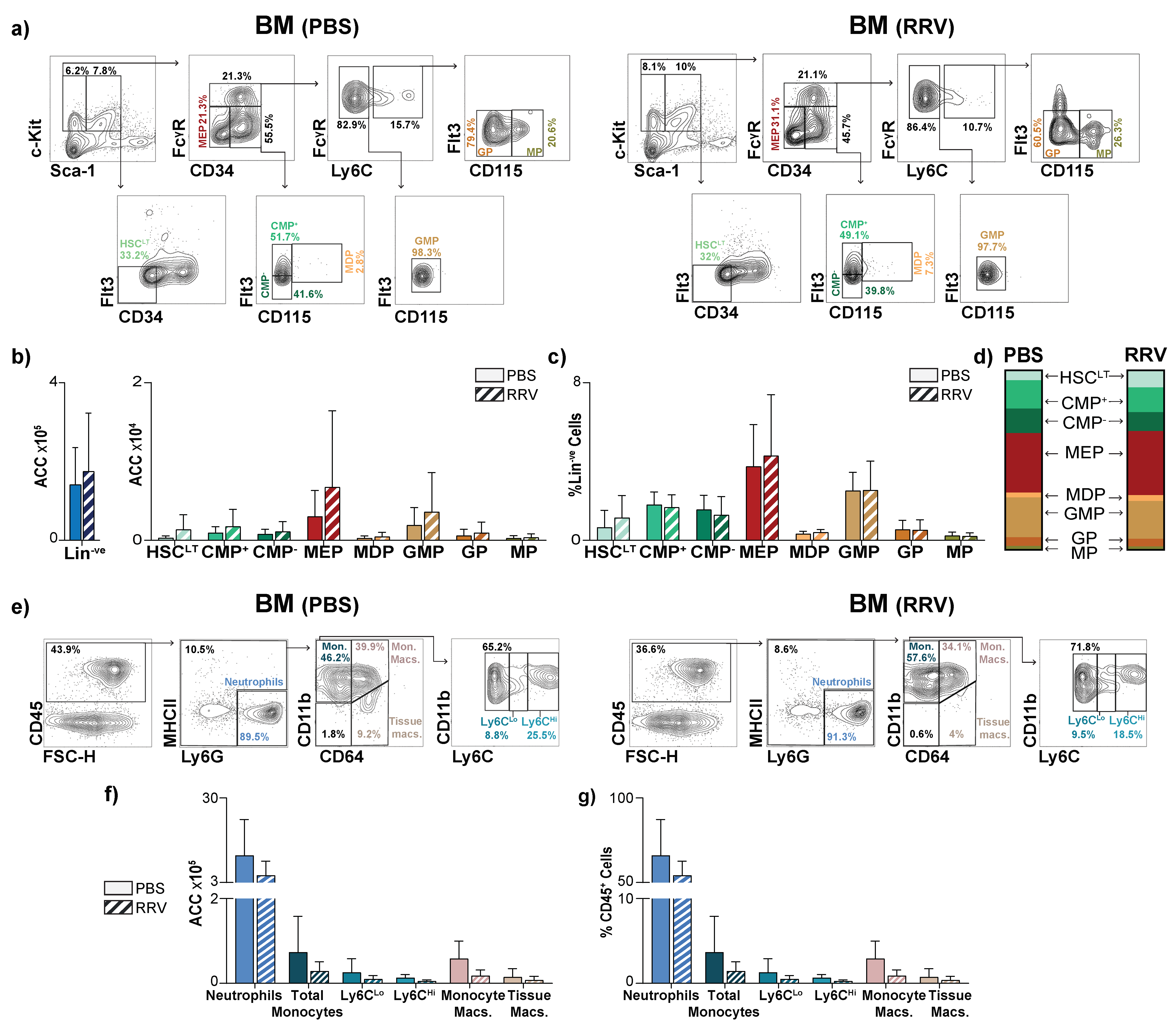
3.4. Perinatal Liver Injury Leads to Contraction of HSPCs and Expansion of Mature Myeloid Populations in the Juvenile Liver
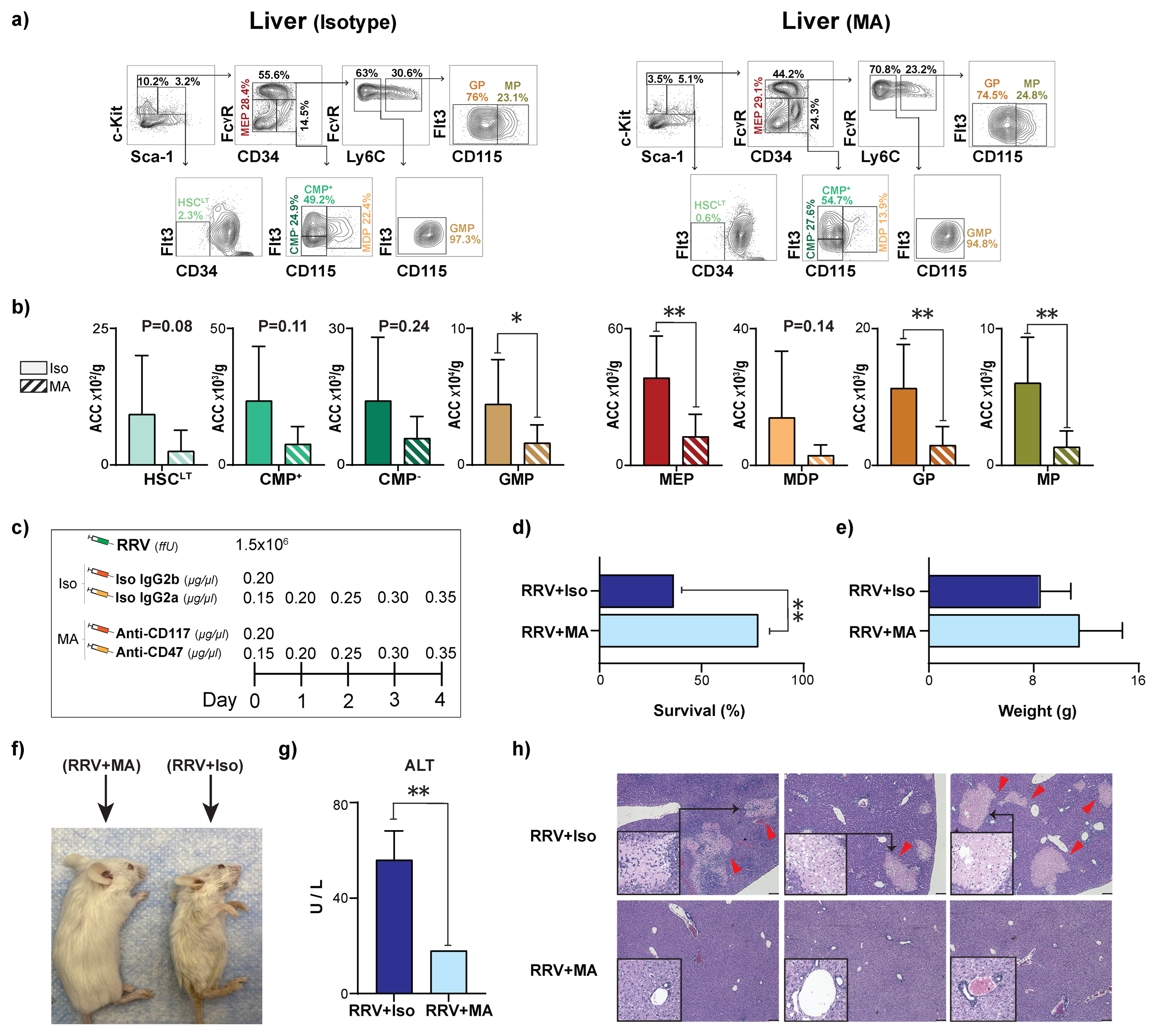
3.5. Myeloablation Protects Mice from RRV-Mediated Perinatal Liver Injury
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | Absolute Cell Count |
| BA | Biliary Atresia |
| BM | Bone Marrow |
| CFU | Colony Forming Units |
| CMP | Common Myeloid Progenitors |
| ffU | Focus Forming Units |
| G-CSF | Granulocyte-colony-stimulating-factor |
| GEMM | Granulocyte, Erythrocyte, Macrophage, Megakaryocyte |
| GM | Granulocyte, Macrophage |
| GMP | Granulocytic-monocytic Progenitors |
| GP | Granulocytic Progenitors |
| H&E | Hematoxylin Eosin |
| HSCLT | Long-term Hematopoietic Stem Cells |
| HSPC | Hematopoietic Stem and Progenitor cells |
| IHC | Immunohistochemistry |
| Iso | Isotype |
| Lin−ve | Lineage negative hematopoietic compartment |
| Lin+ | Lineage positive hematopoietic compartment |
| Ly6CHi | Ly6cHi classical monocytes |
| Ly6CLo | Ly6CLo non-classical monocytes |
| M | Macrophage |
| MA | Myeloablation |
| MDP | Monocytic-dendritic Progenitors |
| MEP | Megakaryocytic-erythroid Progenitors |
| MP | Monocytic and common monocytic Progenitors |
| PLI | Perinatal Liver Injury |
| RRV | Rhesus Rotavirus |
| TMP | Terminal Myeloid Progenitors |
References
- D’Souza, R.; Grammatikopoulos, T.; Pradhan, A.; Sutton, H.; Douiri, A.; Davenport, M.; Verma, A.; Dhawan, A. Acute-on-chronic liver failure in children with biliary atresia awaiting liver transplantation. Pediatr. Transplant. 2019, 23, e13339. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, J.A. The Next Challenge in Pediatric Cholestasis: Deciphering the Pathogenesis of Biliary Atresia. J. Pediatr. Gastroenterol. Nutr. 2006, 43, S23–S29. [Google Scholar] [CrossRef] [PubMed]
- Wehrman, A.; Waisbourd-Zinman, O.; Wells, R.G. Recent advances in understanding biliary atresia. F1000Research 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Alkhani, A.; Levy, C.S.; Tsui, M.; Rosenberg, K.A.; Polovina, K.; Mattis, A.N.; Mack, M.; Van Dyken, S.; Wang, B.M.; Maher, J.J.; et al. Ly6c(Lo) non-classical monocytes promote resolution of rhesus rotavirus-mediated perinatal hepatic inflammation. Sci. Rep. 2020, 10, 7165. [Google Scholar] [CrossRef]
- Ciriza, J.; Thompson, H.; Petrosian, R.; Manilay, J.O.; García-Ojeda, M.E. The migration of hematopoietic progenitors from the fetal liver to the fetal bone marrow: Lessons learned and possible clinical applications. Exp. Hematol. 2013, 41, 411–423. [Google Scholar] [CrossRef]
- Gao, S.; Liu, F. Fetal liver: An ideal niche for hematopoietic stem cell expansion. Sci. China Life Sci. 2018, 61, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, K.S.; Nielson, C.; Firpo, M.A.; Park, A.H.; Beaudin, A.E. Early life inflammation and the developing hematopoietic and immune systems: The cochlea as a sensitive indicator of disruption. Cells 2021, 10, 3596. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Yu, X.; Saha, G.; Kalafati, L.; Ioannidis, C.; Mitroulis, I.; Netea, M.G.; Chavakis, T.; Hajishengallis, G. Maladaptive innate immune training of myelopoiesis links inflammatory comorbidities. Cell 2022, 185, 1709–1727.e18. [Google Scholar] [CrossRef]
- Chavakis, T.; Mitroulis, I.; Hajishengallis, G. Hematopoietic progenitor cells as integrative hubs for adaptation to and fine-tuning of inflammation. Nat. Immunol. 2019, 20, 802–811. [Google Scholar] [CrossRef]
- Eckert, C.; Klein, N.; Kornek, M.; Lukacs-Kornek, V. The complex myeloid network of the liver with diverse functional capacity at steady state and in inflammation. Front. Immunol. 2015, 6, 179. [Google Scholar] [CrossRef]
- Weston, C.J.; Zimmermann, H.W.; Adams, D.H. The role of myeloid-derived cells in the progression of liver disease. Front. Immunol. 2019, 10, 893. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P.; Jenne, C.; Snyder, J. Immune Responses in the Liver. Annu. Rev. Immunol. 2018, 44, 247–277. [Google Scholar] [CrossRef] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Yáñez, A.; Goodridge, H.S. Identification and isolation of oligopotent and lineage-committed myeloid progenitors from mouse bone marrow. J. Vis. Exp. 2018, 2018, e58061. [Google Scholar] [CrossRef] [Green Version]
- Copley, M.R.; Eaves, C.J. Developmental changes in hematopoietic stem cell properties. Exp. Mol. Med. 2013, 45, e55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oetzmann Von Sochaczewski, C.; Pintelon, I.; Brouns, I.; Dreier, A.; Klemann, C.; Timmermans, J.-P.; Petersen, C.; Kuebler, J.F. Rotavirus particles in the extrahepatic bile duct in experimental biliary atresia. J. Pediatr. Surg. 2014, 49, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, A.; Ring, A.M.; Weiskopf, K.; Schnorr, P.J.; Gordon, S.; Le, A.C.; Kwon, H.-S.; Ring, N.G.; Volkmer, J.; Ho, P.Y.; et al. Hematopoietic stem cell transplantation in immunocompetent hosts without radiation or chemotherapy. Sci. Transl. Med. 2016, 8, 351ra105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, W.W.; Czechowicz, A.; Logan, A.C.; Bhardwaj, R.; Poyser, J.; Park, C.Y.; Weissman, I.L.; Shizuru, J.A. Anti-CD117 antibody depletes normal and myelodysplastic syndrome human hematopoietic stem cells in xenografted mice. Blood 2019, 133, 2069–2078. [Google Scholar] [CrossRef]
- Wang, X.Q.; Lo, C.M.; Chen, L.; Cheung, C.K.; Yang, Z.F.; Chen, Y.X.; Ng, M.N.; Yu, W.C.; Ming, X.; Zhang, W.; et al. Hematopoietic chimerism in liver transplantation patients and hematopoietic stem/progenitor cells in adult human liver. Hepatology 2012, 56, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Soares-da-Silva, F.; Peixoto, M.; Cumano, A.; Pinto-do-Ó, P. Crosstalk Between the Hepatic and Hematopoietic Systems During Embryonic Development. Front. Cell Dev. Biol. 2020, 8, 612. [Google Scholar] [CrossRef]
- King, K.Y.; Goodell, M.A. Inflammatory modulation of HSCs: Viewing the HSC as a foundation for the immune response. Nat. Rev. Immunol. 2011, 11, 685–692. [Google Scholar] [CrossRef]
- Apostol, A.C.; Jensen, K.D.C.; Beaudin, A.E. Training the Fetal Immune System Through Maternal Inflammation-A Layered Hygiene Hypothesis. Front. Immunol. 2020, 11, 123. [Google Scholar] [CrossRef]
- Holterman, A.X.; Nguyen, H.P.A.; Nadler, E.; Vu, G.H.; Mohan, P.; Vu, M.; Trinh, T.T.; Bui, H.T.T.; Nguyen, B.T.; Quynh, A.T.; et al. Granulocyte-colony stimulating factor GCSF mobilizes hematopoietic stem cells in Kasai patients with biliary atresia in a phase 1 study and improves short term outcome. J. Pediatr. Surg. 2021, 56, 1179–1185. [Google Scholar] [CrossRef]
- Liu, J.; Fei, Y.; Zhou, T.; Ji, H.; Wu, J.; Gu, X.; Luo, Y.; Zhu, J.; Feng, M.; Wan, P.; et al. Bile Acids Impair Vaccine Response in Children With Biliary Atresia. Front. Immunol. 2021, 12, 642546. [Google Scholar] [CrossRef]
- Wu, J.F.; Ni, Y.H.; Chen, H.L.; Hsu, H.Y.; Lai, H.S.; Chang, M.H. Humoral immunogenicity to measles, rubella, and varicella-zoster vaccines in biliary atresia children. Vaccine 2009, 27, 2812–2815. [Google Scholar] [CrossRef]
- Ruth, N.D.; Kelly, D.; Sharif, K.; Morland, B.; Lloyd, C.; McKiernan, P.J. Rejection is less common in children undergoing liver transplantation for hepatoblastoma. Pediatr. Transplant. 2014, 18, 52–57. [Google Scholar] [CrossRef]
- Divangahi, M.; Aaby, P.; Khader, S.A.; Barreiro, L.B.; Bekkering, S.; Chavakis, T.; van Crevel, R.; Curtis, N.; DiNardo, A.R.; Dominguez-Andres, J.; et al. Trained immunity, tolerance, priming and differentiation: Distinct immunological processes. Nat. Immunol. 2021, 22, 2–6. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Derderian, S.C.; Togarrati, P.P.; King, C.; Moradi, P.W.; Reynaud, D.; Czechowicz, A.; Weissman, I.L.; MacKenzie, T.C. In utero depletion of fetal hematopoietic stem cells improves engraftment after neonatal transplantation in mice. Blood 2014, 124, 973–980. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Zhao, F.; Tseng, S.; Narayanan, C.; Shura, L.; Willingham, S.; Howard, M.; Prohaska, S.; Volkmer, J.; et al. Pre-Clinical Development of a Humanized Anti-CD47 Antibody with Anti-Cancer Therapeutic Potential. PLoS ONE 2015, 10, e0137345. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhani, A.; Korsholm, C.; Levy, C.S.; Mohamedaly, S.; Duwaerts, C.C.; Pietras, E.M.; Nijagal, A. Neonatal Hepatic Myeloid Progenitors Expand and Propagate Liver Injury in Mice. J. Clin. Med. 2023, 12, 337. https://doi.org/10.3390/jcm12010337
Alkhani A, Korsholm C, Levy CS, Mohamedaly S, Duwaerts CC, Pietras EM, Nijagal A. Neonatal Hepatic Myeloid Progenitors Expand and Propagate Liver Injury in Mice. Journal of Clinical Medicine. 2023; 12(1):337. https://doi.org/10.3390/jcm12010337
Chicago/Turabian StyleAlkhani, Anas, Cathrine Korsholm, Claire S. Levy, Sarah Mohamedaly, Caroline C. Duwaerts, Eric M. Pietras, and Amar Nijagal. 2023. "Neonatal Hepatic Myeloid Progenitors Expand and Propagate Liver Injury in Mice" Journal of Clinical Medicine 12, no. 1: 337. https://doi.org/10.3390/jcm12010337