
Hepatic Ly6CLo Non-Classical Monocytes Have Increased Nr4a1 (Nur77) in Murine Biliary Atresia
 , and
, and 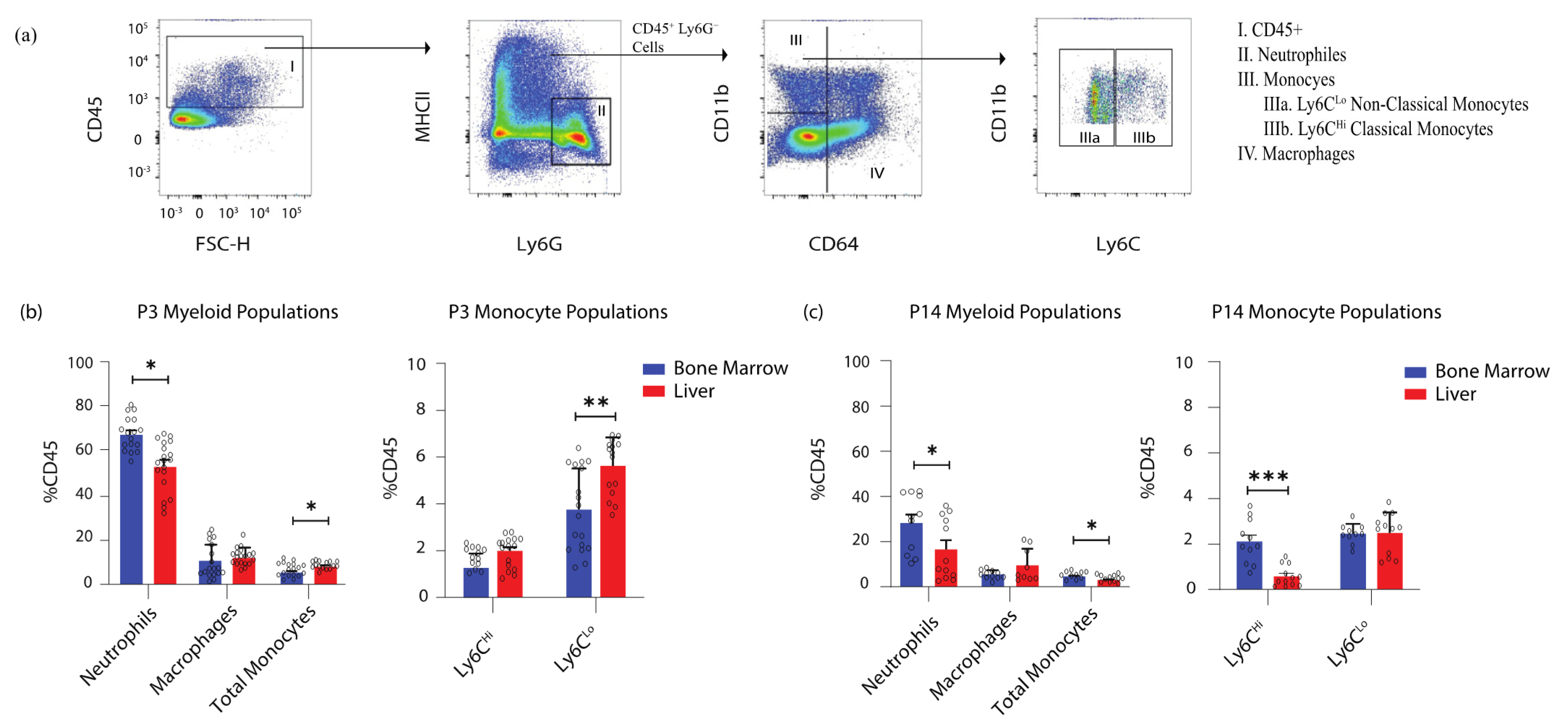{kind=link}
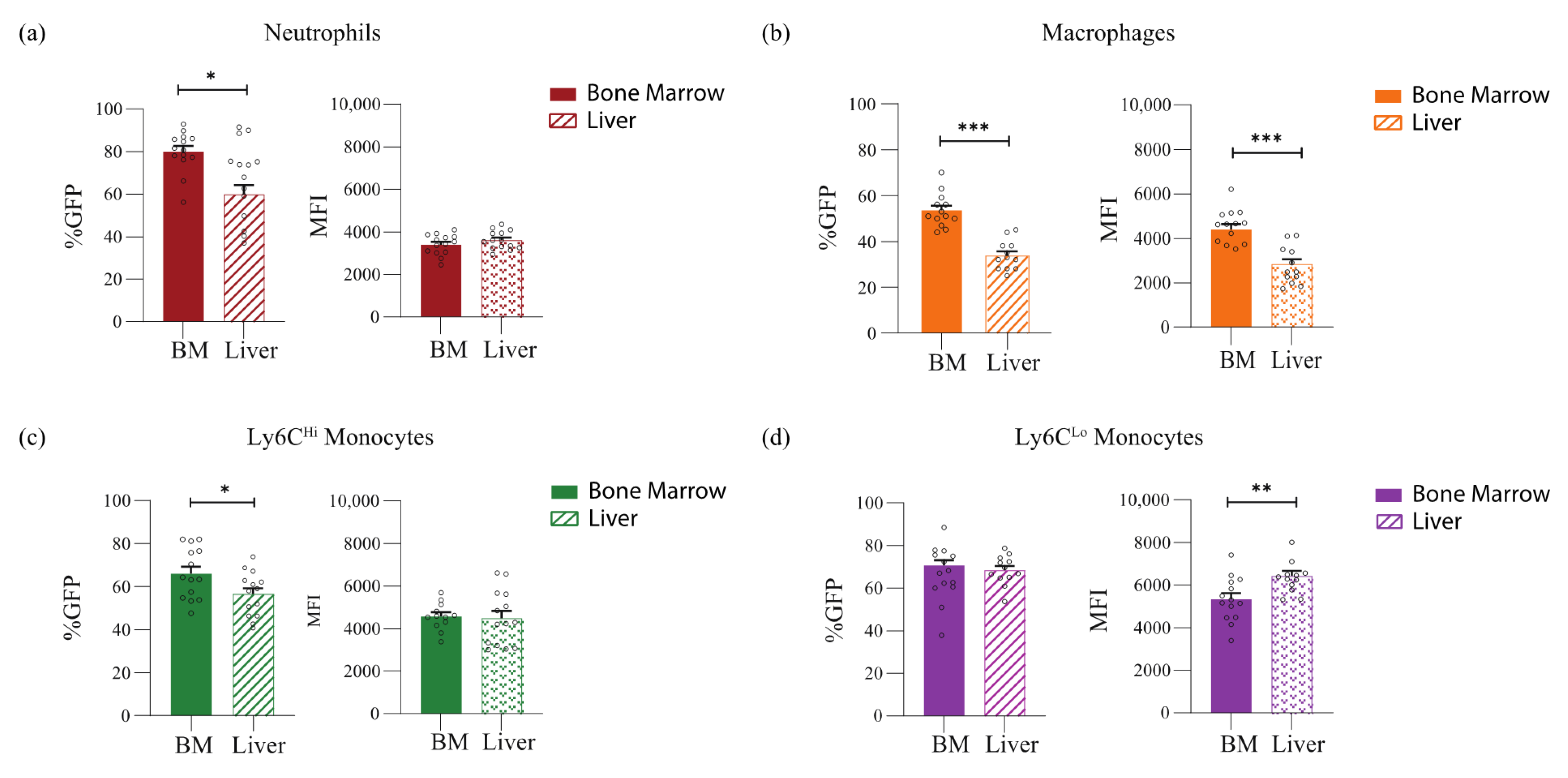{kind=link}
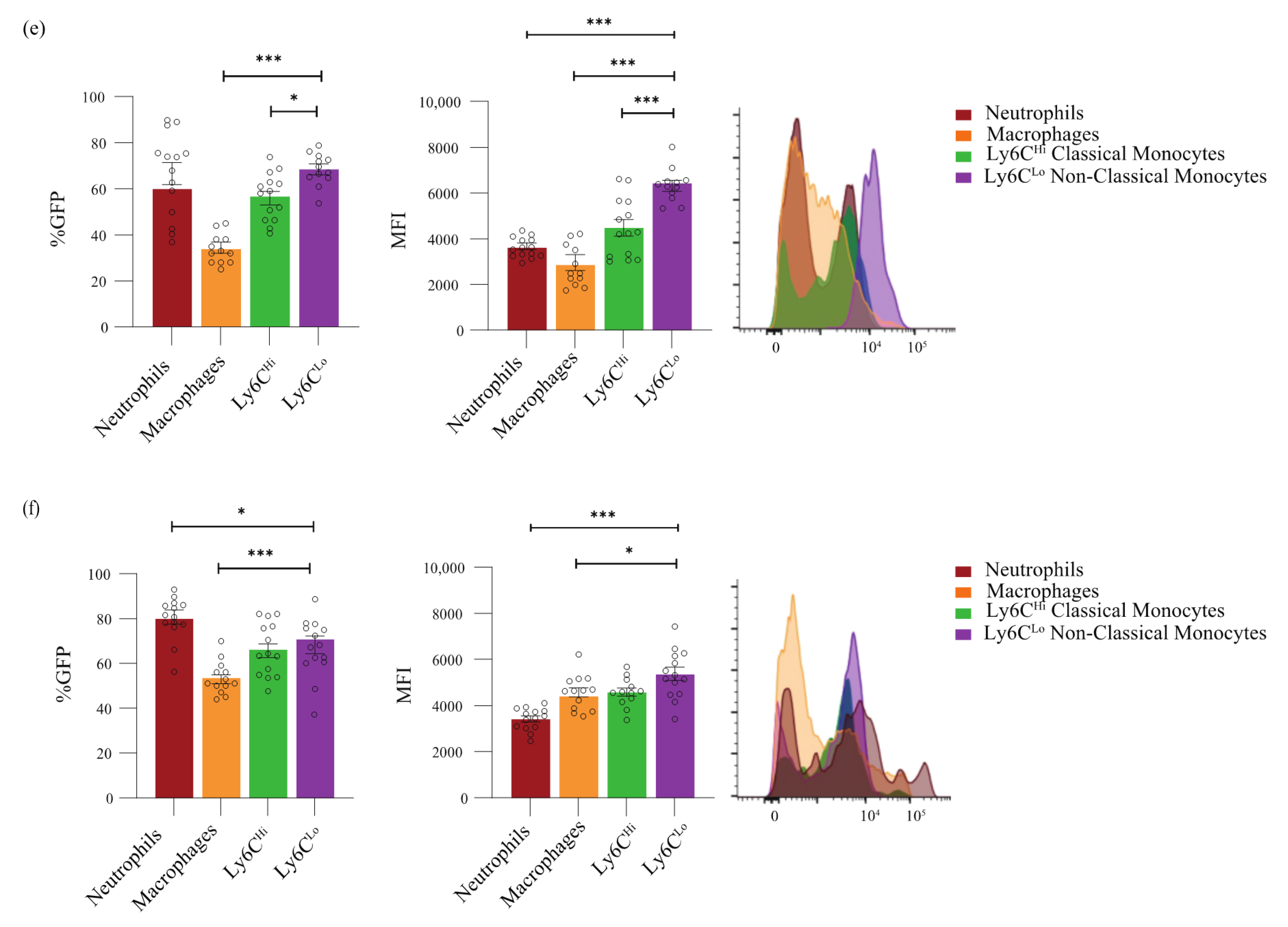{kind=link}
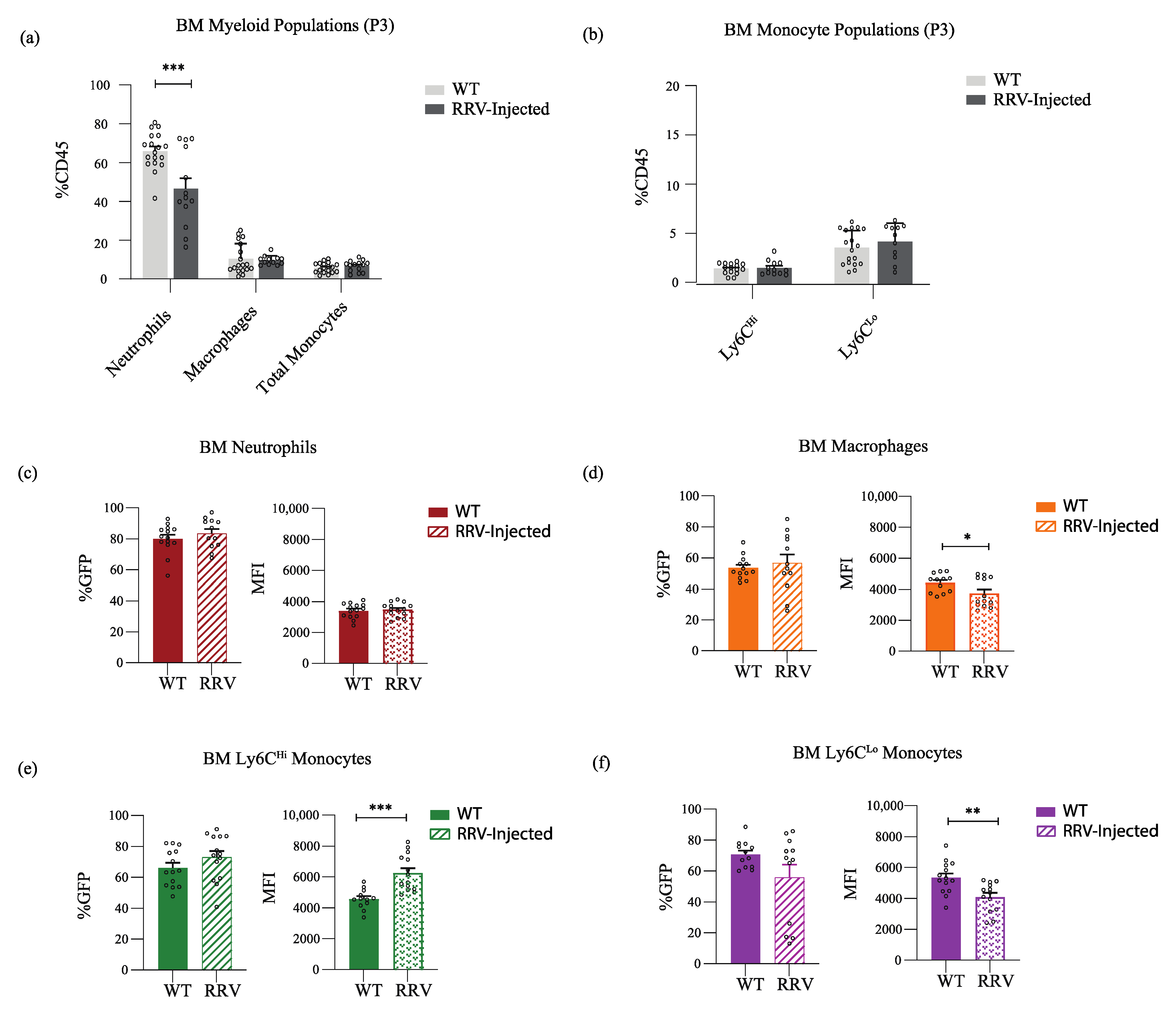{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
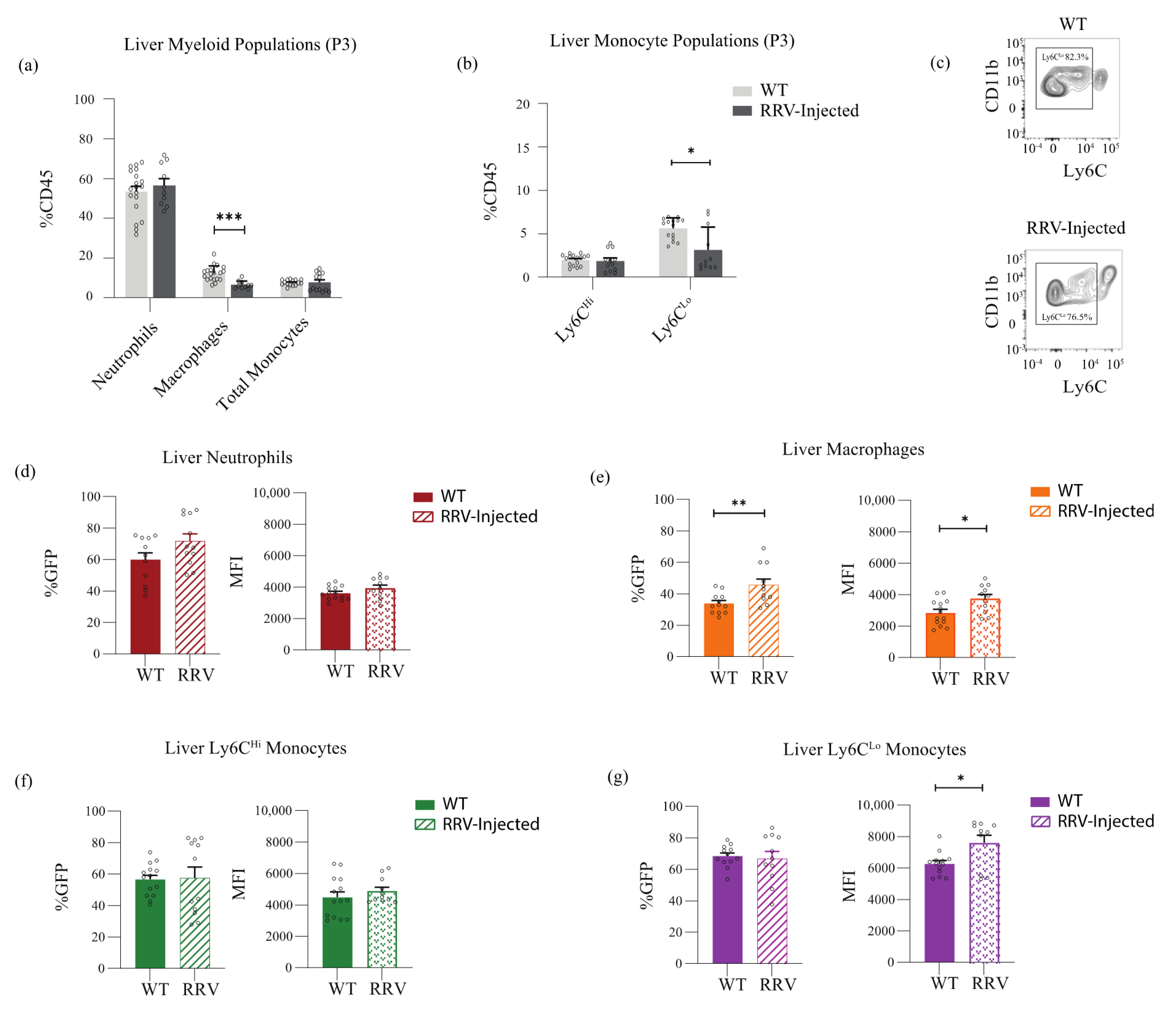
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartley, J.L.; Davenport, M.; Kelly, D.A. Biliary atresia. Lancet 2009, 374, 1704–1713. [Google Scholar] [CrossRef]
- Czaja, A.J. Hepatic inflammation and progressive liver fibrosis in chronic liver disease. World J. Gastroenterol. 2014, 20, 2515–2532. [Google Scholar] [CrossRef] [PubMed]
- Brempelis, K.J.; Crispe, I.N. Infiltrating monocytes in liver injury and repair. Clin. Transl. Immunol. 2016, 5, e113. [Google Scholar] [CrossRef] [PubMed]
- Liaskou, E.; Wilson, D.V.; Oo, Y.H. Innate immune cells in liver inflammation. Mediat. Inflamm. 2012, 2012, 949157. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O'Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Sander, L.E.; Sackett, S.D.; Dierssen, U.; Beraza, N.; Linke, R.P.; Muller, M.; Blander, J.M.; Tacke, F.; Trautwein, C. Hepatic acute-phase proteins control innate immune responses during infection by promoting myeloid-derived suppressor cell function. J. Exp. Med. 2010, 207, 1453–1464. [Google Scholar] [CrossRef]
- Lee, P.Y.; Nelson-Maney, N.; Huang, Y.; Levescot, A.; Wang, Q.; Wei, K.; Cunin, P.; Li, Y.; Lederer, J.A.; Zhuang, H.; et al. High-dimensional analysis reveals a pathogenic role of inflammatory monocytes in experimental diffuse alveolar hemorrhage. JCI Insight 2019, 4, e129703. [Google Scholar] [CrossRef]
- Olingy, C.E.; San Emeterio, C.L.; Ogle, M.E.; Krieger, J.R.; Bruce, A.C.; Pfau, D.D.; Jordan, B.T.; Peirce, S.M.; Botchwey, E.A. Non-classical monocytes are biased progenitors of wound healing macrophages during soft tissue injury. Sci. Rep. 2017, 7, 447. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, J.; Qi, H.; Wei, J.; Yang, Y.; Yue, J.; Liu, X.; Zhang, Y.; Yang, R. LncRNA lncLy6C induced by microbiota metabolite butyrate promotes differentiation of Ly6C(high) to Ly6C(int/neg) macrophages through lncLy6C/C/EBPbeta/Nr4A1 axis. Cell Discov. 2020, 6, 87. [Google Scholar] [CrossRef]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Morias, Y.; Abels, C.; Laoui, D.; Van Overmeire, E.; Guilliams, M.; Schouppe, E.; Tacke, F.; deVries, C.J.; De Baetselier, P.; Beschin, A. Ly6C- Monocytes Regulate Parasite-Induced Liver Inflammation by Inducing the Differentiation of Pathogenic Ly6C+ Monocytes into Macrophages. PLoS Pathog. 2015, 11, e1004873. [Google Scholar] [CrossRef] [PubMed]
- Alkhani, A.; Levy, C.S.; Tsui, M.; Rosenberg, K.A.; Polovina, K.; Mattis, A.N.; Mack, M.; Van Dyken, S.; Wang, B.M.; Maher, J.J.; et al. Ly6c(Lo) non-classical monocytes promote resolution of rhesus rotavirus-mediated perinatal hepatic inflammation. Sci. Rep. 2020, 10, 7165. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.N.; Carlin, L.M.; Hubbeling, H.G.; Nackiewicz, D.; Green, A.M.; Punt, J.A.; Geissmann, F.; Hedrick, C.C. The transcription factor NR4A1 (Nur77) controls bone marrow differentiation and the survival of Ly6C- monocytes. Nat. Immunol. 2011, 12, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Herring, J.A.; Elison, W.S.; Tessem, J.S. Function of Nr4a Orphan Nuclear Receptors in Proliferation, Apoptosis and Fuel Utilization Across Tissues. Cells 2019, 8, 1373. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, J.; Badimon, L. The NR4A subfamily of nuclear receptors: New early genes regulated by growth factors in vascular cells. Cardiovasc. Res. 2005, 65, 609–618. [Google Scholar] [CrossRef]
- Pawlak, A.; Strzadala, L.; Kalas, W. Non-genomic effects of the NR4A1/Nur77/TR3/NGFIB orphan nuclear receptor. Steroids 2015, 95, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, M.A.; Muscat, G.E. The NR4A subgroup: Immediate early response genes with pleiotropic physiological roles. Nucl. Recept. Signal. 2006, 4, e002. [Google Scholar] [CrossRef]
- Mohan, H.M.; Aherne, C.M.; Rogers, A.C.; Baird, A.W.; Winter, D.C.; Murphy, E.P. Molecular pathways: The role of NR4A orphan nuclear receptors in cancer. Clin. Cancer Res. 2012, 18, 3223–3228. [Google Scholar] [CrossRef]
- Pearen, M.A.; Muscat, G.E. Minireview: Nuclear hormone receptor 4A signaling: Implications for metabolic disease. Mol. Endocrinol. 2010, 24, 1891–1903. [Google Scholar] [CrossRef]
- Zikherman, J.; Parameswaran, R.; Weiss, A. Endogenous antigen tunes the responsiveness of naive B cells but not T cells. Nature 2012, 489, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Li, K.; Yang, L.; Tang, S.T.; Wang, X.X.; Cao, G.Q.; Li, S.; Lei, H.Y.; Zhang, X. Dendritic Cells Regulate Treg-Th17 Axis in Obstructive Phase of Bile Duct Injury in Murine Biliary Atresia. PLoS ONE 2015, 10, e0136214. [Google Scholar] [CrossRef]
- Riepenhoff-Talty, M.; Schaekel, K.; Clark, H.F.; Mueller, W.; Uhnoo, I.; Rossi, T.; Fisher, J.; Ogra, P.L. Group A rotaviruses produce extrahepatic biliary obstruction in orally inoculated newborn mice. Pediatr. Res. 1993, 33, 394–399. [Google Scholar] [CrossRef]
- Shivakumar, P.; Campbell, K.M.; Sabla, G.E.; Miethke, A.; Tiao, G.; McNeal, M.M.; Ward, R.L.; Bezerra, J.A. Obstruction of extrahepatic bile ducts by lymphocytes is regulated by IFN-gamma in experimental biliary atresia. J. Clin. Investig. 2004, 114, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Shivakumar, P.; Sabla, G.E.; Whitington, P.; Chougnet, C.A.; Bezerra, J.A. Neonatal NK cells target the mouse duct epithelium via Nkg2d and drive tissue-specific injury in experimental biliary atresia. J. Clin. Investig. 2009, 119, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Clapp, D.W.; Dumenco, L.L.; Hatzoglou, M.; Gerson, S.L. Fetal liver hematopoietic stem cells as a target for in utero retroviral gene transfer. Blood 1991, 78, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Keller, G.; Lacaud, G.; Robertson, S. Development of the hematopoietic system in the mouse. Exp. Hematol. 1999, 27, 777–787. [Google Scholar] [CrossRef]
- Wolber, F.M.; Leonard, E.; Michael, S.; Orschell-Traycoff, C.M.; Yoder, M.C.; Srour, E.F. Roles of spleen and liver in development of the murine hematopoietic system. Exp. Hematol. 2002, 30, 1010–1019. [Google Scholar] [CrossRef]
- Cumano, A.; Ferraz, J.C.; Klaine, M.; Di Santo, J.P.; Godin, I. Intraembryonic, but not yolk sac hematopoietic precursors, isolated before circulation, provide long-term multilineage reconstitution. Immunity 2001, 15, 477–485. [Google Scholar] [CrossRef]
- Golub, R.; Cumano, A. Embryonic hematopoiesis. Blood Cells Mol. Dis. 2013, 51, 226–231. [Google Scholar] [CrossRef]
- McGrath, K.E.; Frame, J.M.; Palis, J. Early hematopoiesis and macrophage development. Semin. Immunol. 2015, 27, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Ema, H.; Nakauchi, H. Expansion of hematopoietic stem cells in the developing liver of a mouse embryo. Blood 2000, 95, 2284–2288. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Tajima, F.; Ogawa, M. Developmental changes of CD34 expression by murine hematopoietic stem cells. Exp. Hematol. 2000, 28, 1269–1273. [Google Scholar] [CrossRef]
- Hettinger, J.; Richards, D.M.; Hansson, J.; Barra, M.M.; Joschko, A.C.; Krijgsveld, J.; Feuerer, M. Origin of monocytes and macrophages in a committed progenitor. Nat. Immunol. 2013, 14, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Dal-Secco, D.; Wang, J.; Zeng, Z.; Kolaczkowska, E.; Wong, C.H.; Petri, B.; Ransohoff, R.M.; Charo, I.F.; Jenne, C.N.; Kubes, P. A dynamic spectrum of monocytes arising from the in situ reprogramming of CCR2+ monocytes at a site of sterile injury. J. Exp. Med. 2015, 212, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Sunderkotter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef]
- Yona, S.; Kim, K.W.; Wolf, Y.; Mildner, A.; Varol, D.; Breker, M.; Strauss-Ayali, D.; Viukov, S.; Guilliams, M.; Misharin, A.; et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 2013, 38, 79–91. [Google Scholar] [CrossRef]
- Auffray, C.; Sieweke, M.H.; Geissmann, F. Blood monocytes: Development, heterogeneity, and relationship with dendritic cells. Annu. Rev. Immunol. 2009, 27, 669–692. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamedaly, S.; Levy, C.S.; Korsholm, C.; Alkhani, A.; Rosenberg, K.; Ashouri, J.F.; Nijagal, A. Hepatic Ly6CLo Non-Classical Monocytes Have Increased Nr4a1 (Nur77) in Murine Biliary Atresia. J. Clin. Med. 2022, 11, 5290. https://doi.org/10.3390/jcm11185290
Mohamedaly S, Levy CS, Korsholm C, Alkhani A, Rosenberg K, Ashouri JF, Nijagal A. Hepatic Ly6CLo Non-Classical Monocytes Have Increased Nr4a1 (Nur77) in Murine Biliary Atresia. Journal of Clinical Medicine. 2022; 11(18):5290. https://doi.org/10.3390/jcm11185290
Chicago/Turabian StyleMohamedaly, Sarah, Claire S. Levy, Cathrine Korsholm, Anas Alkhani, Katherine Rosenberg, Judith F. Ashouri, and Amar Nijagal. 2022. "Hepatic Ly6CLo Non-Classical Monocytes Have Increased Nr4a1 (Nur77) in Murine Biliary Atresia" Journal of Clinical Medicine 11, no. 18: 5290. https://doi.org/10.3390/jcm11185290