
Phenotypic and Functional Alterations of Immune Effectors in Periodontitis; A Multifactorial and Complex Oral Disease

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Reagents and Antibodies
2.2. Donor Selection and Diagnostic Criteria
2.3. Isolation of Peripheral and Oral Blood Mononuclear Cells
2.4. Mononuclear Cells Purified from Gingival Tissues
2.5. Enzyme-Linked Immunosorbent Assays (ELISAs) and Multiplex Assays
2.6. Cytotoxicity Assays
2.7. Surface Staining and Cell Death Assays
2.8. Purification of Human NK Cells
2.9. Retroviral Transduction, Transfection and the Generation of Tumor Cell Transfectants
2.10. Luciferase Reporter Assay
2.11. Fusobacterium nucleatum Preparation
2.12. Statistical Analysis
3. Results
3.1. Periodontitis Patients’ Oral Blood Exhibited Higher Percentages of NK Cells and Lower Percentages of B Cells in Comparison to Their Peripheral Blood
3.2. Oral Gingival-Derived Immune Cells from Periodontitis Patients Exhibited Higher Percentages of B Cells and Lower Percentages of T Cells as Compared to Those from Healthy Individuals
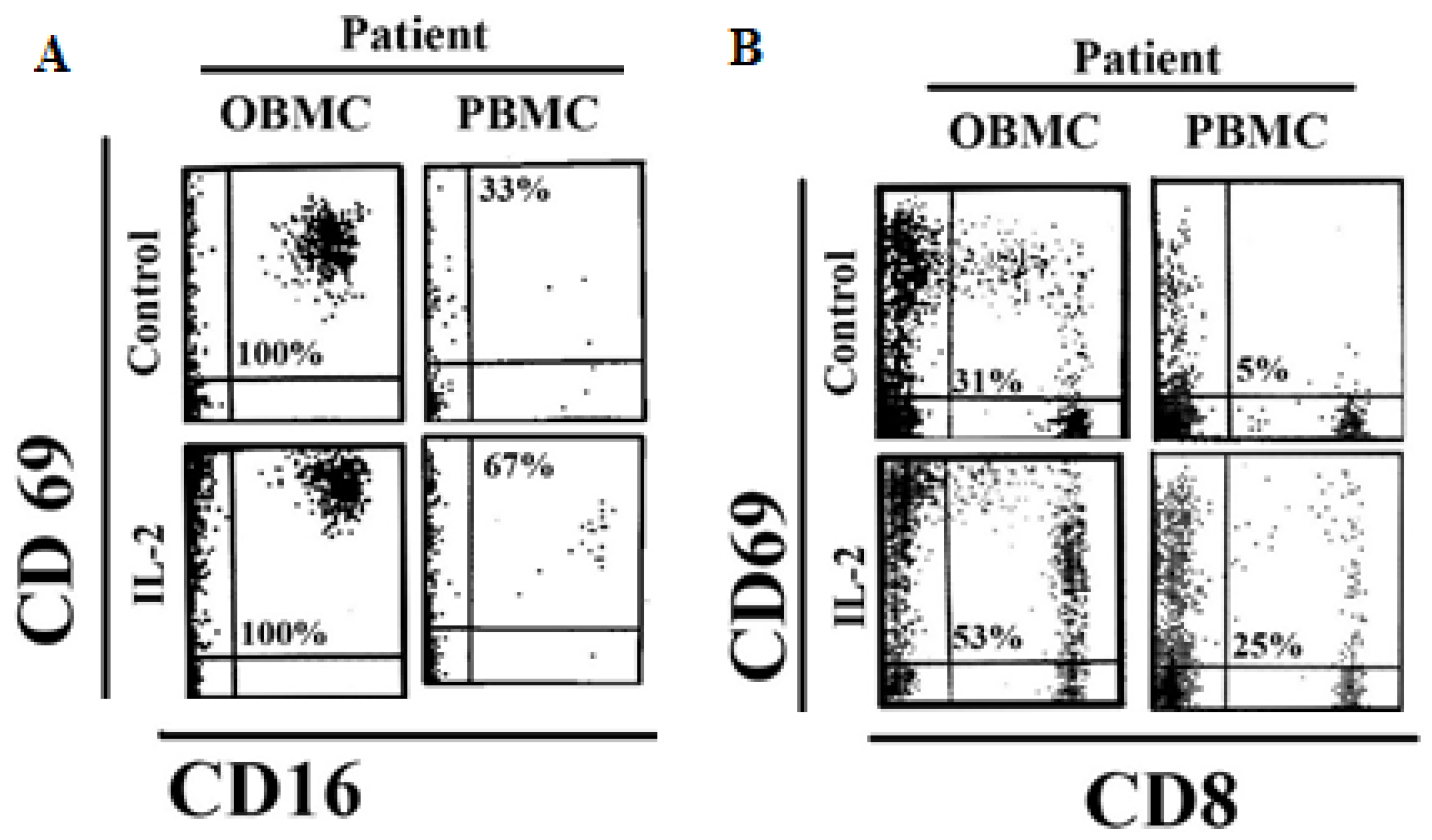
3.3. NK Cells and CD8 + T Cells in Oral Blood of Periodontitis Patients Exhibited Significant Levels of Activation
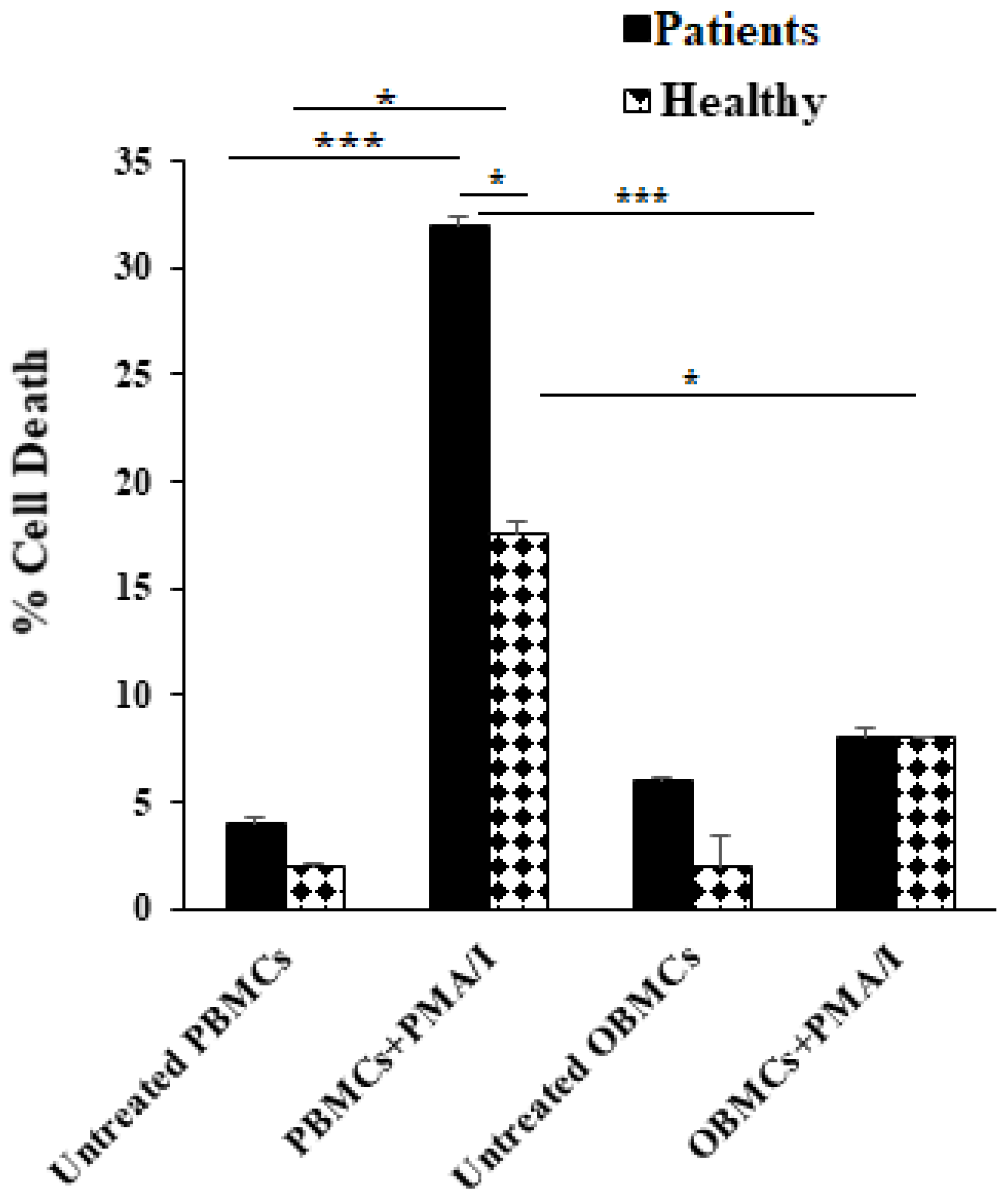
3.4. PBMCs but Not OBMCs Obtained from Periodontitis Patients Demonstrated Significantly Higher Levels of Cell Death When Compared to Those from Healthy Individuals
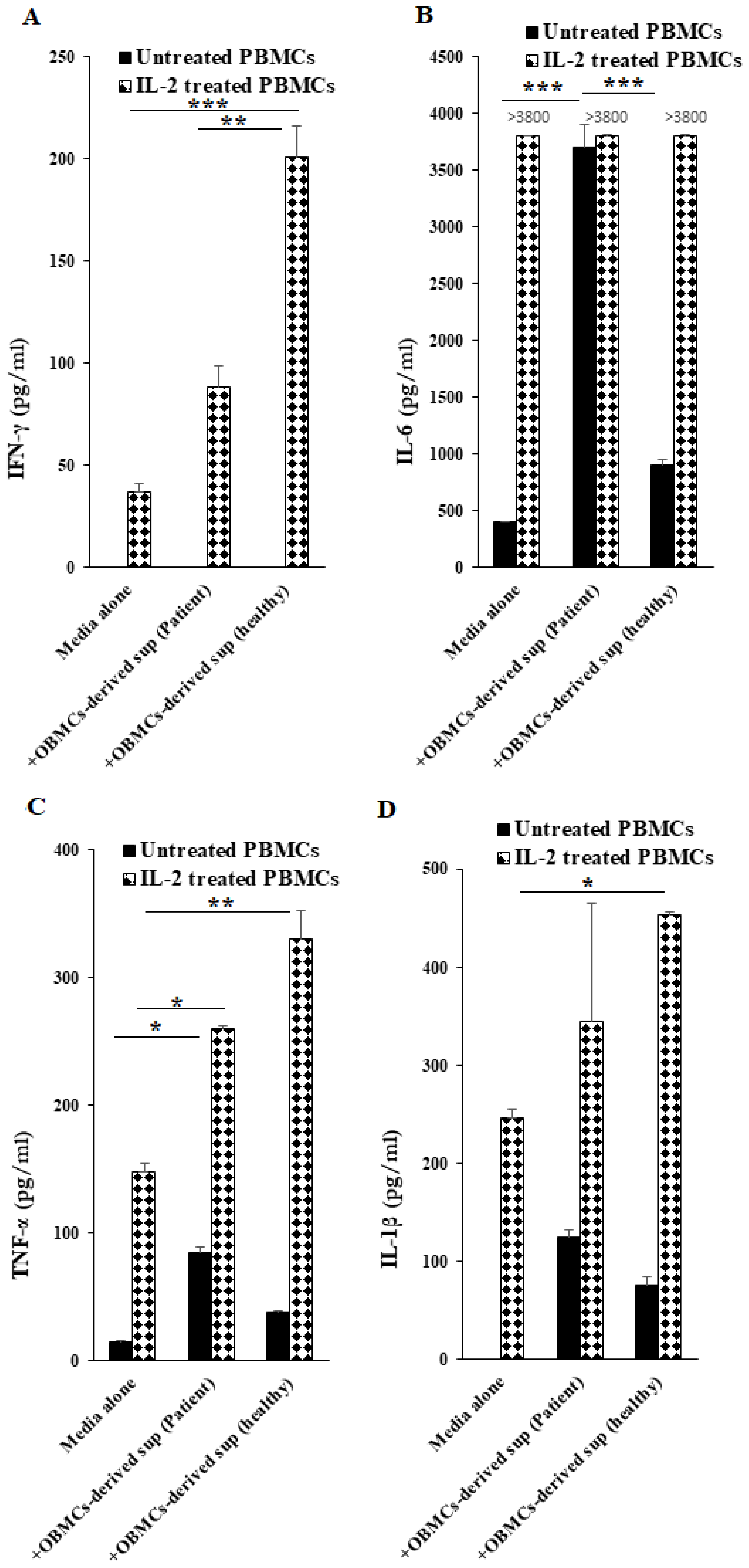
3.5. Increased TNF-α Secretion Was Observed from OBMCs of Periodontitis Patients When Compared to Those from Healthy Individuals
3.6. OBMC-Derived Supernatants from Periodontitis Patients Regulate Secretion of Cytokines by Allogeneic Healthy PBMCs
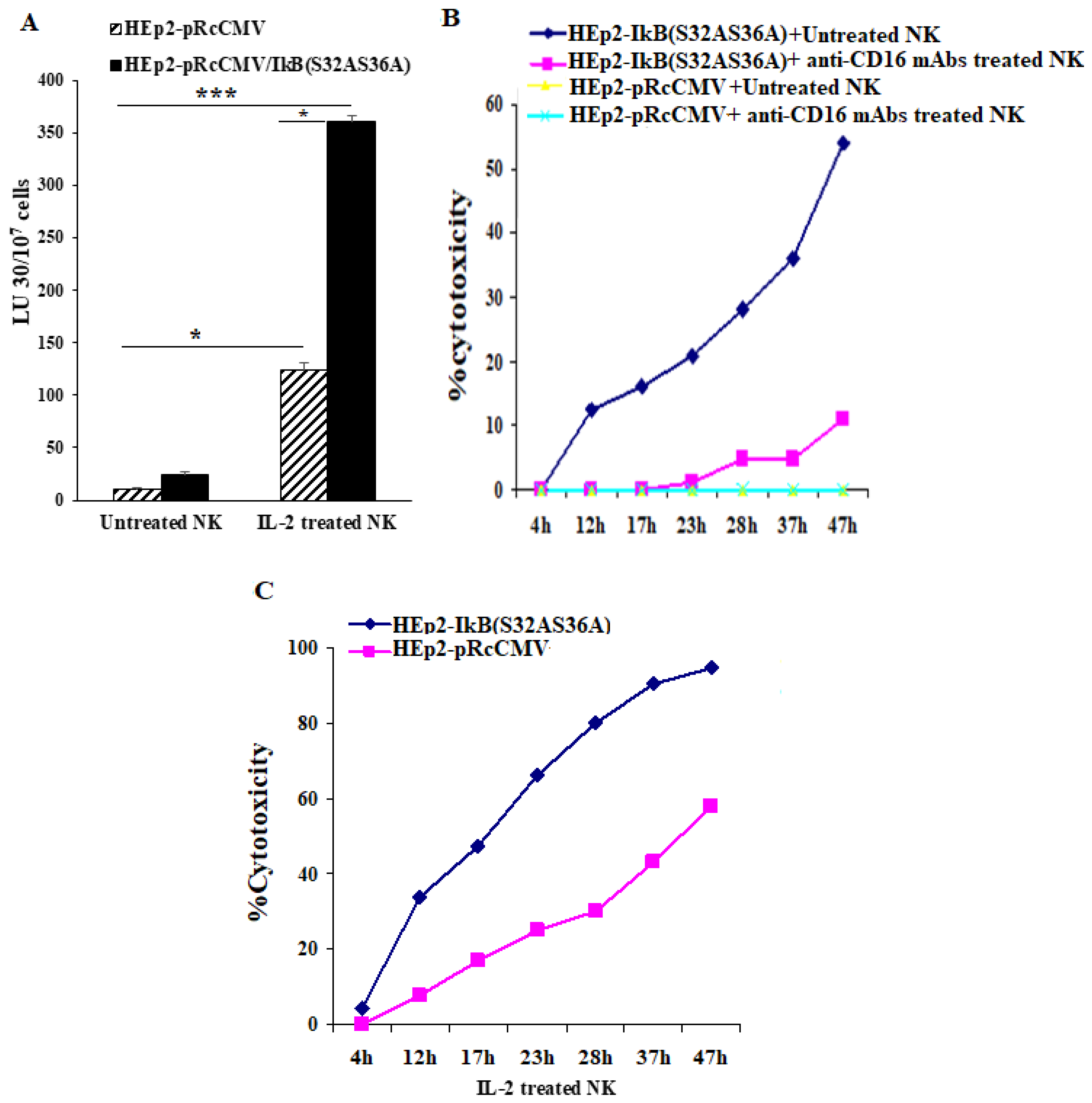
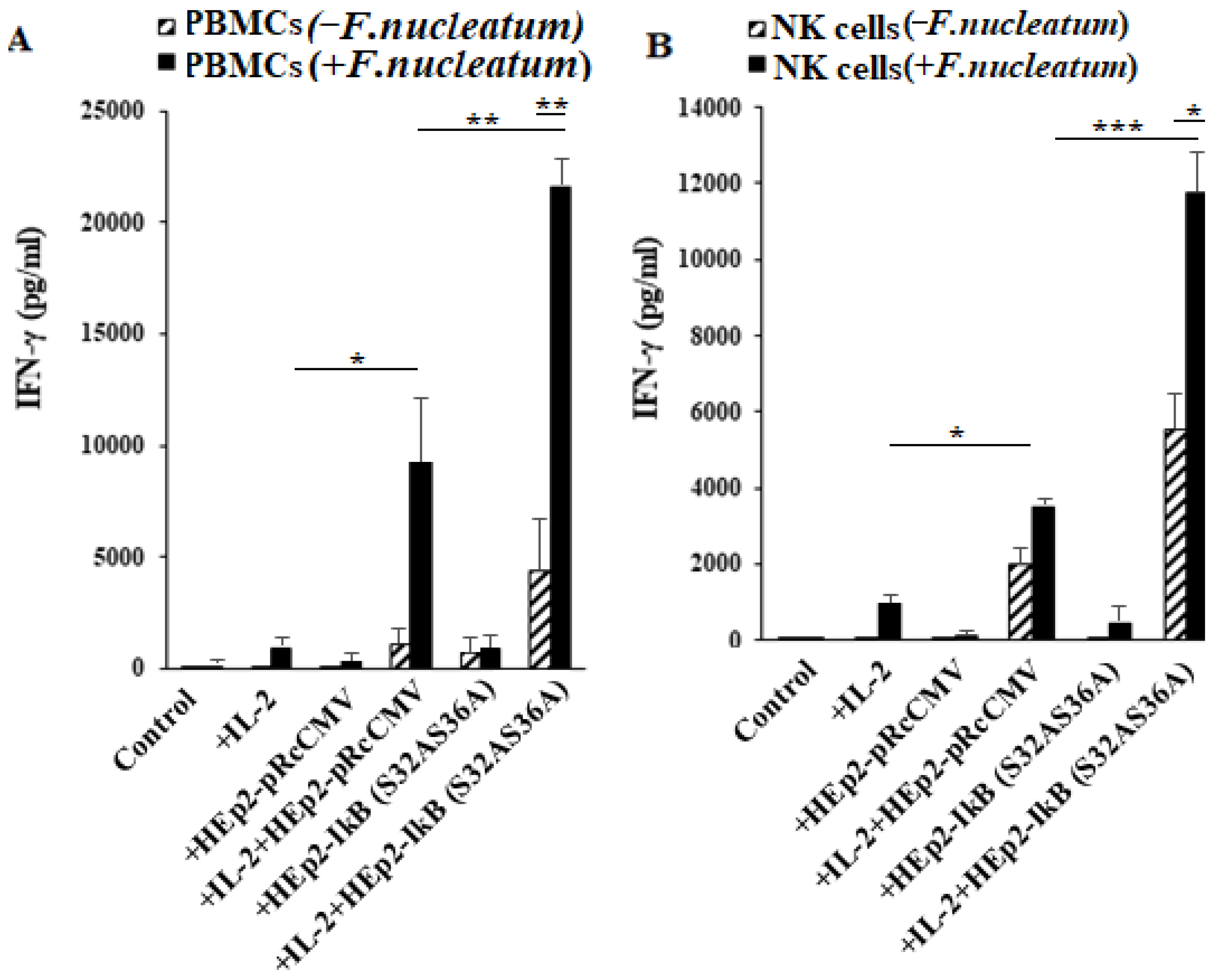
3.7. NFkB Deletion in Oral Epithelial Cells Increases IFN-γ Secretion by PBMCs and NK Cells in the Presence or Absence of Fusobacterium nucleatum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NK cells | Natural killer cells |
| OSCCs | Oral squamous carcinoma cells |
| IFN-γ | Interferon-gamma |
| TNF-α | Tumor necrosis factor-α |
| OBMCs | Oral blood mononuclear cells |
| PBMCs | Peripheral blood mononuclear cells |
| NFκB | Nuclear factor kappa B |
| IL | Interleukin |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
References
- Slots, J. Periodontology: Past, present, perspectives. Periodontol 2000 2013, 62, 7–19. [Google Scholar] [CrossRef]
- Gemmell, E.; Yamazaki, K.; Seymour, G.J. Destructive periodontitis lesions are determined by the nature of the lymphocytic response. Crit. Rev. Oral Biol. Med. 2002, 13, 17–34. [Google Scholar] [CrossRef]
- Teng, Y.T. The role of acquired immunity and periodontal disease progression. Crit. Rev. Oral Biol. Med. 2003, 14, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Page, R.C.; Kornman, K.S. The pathogenesis of human periodontitis: An introduction. Periodontol 2000 1997, 14, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F. Causation and pathogenesis of periodontal disease. Periodontol 2000 2001, 25, 8–20. [Google Scholar] [CrossRef]
- Kornman, K.S.; Page, R.C.; Tonetti, M.S. The host response to the microbial challenge in periodontitis: Assembling the players. Periodontol 2000 1997, 14, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Page, R.C. The role of inflammatory mediators in the pathogenesis of periodontal disease. J. Periodontal Res. 1991, 26, 230–242. [Google Scholar] [CrossRef]
- Graves, D.T. The potential role of chemokines and inflammatory cytokines in periodontal disease progression. Clin. Infect. Dis. 1999, 28, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, U.; Solominidou, E.; Korkmaz, Y.; Rüttermann, S.; Klocke, A.; Flemmig, T.F.; Beikler, T. Regulator of Calcineurin 1 in Periodontal Disease. Mediat. Inflamm. 2016, 2016, 5475821. [Google Scholar] [CrossRef]
- Venugopal, P.; Koshy, T.; Lavu, V.; Rao, S.R.; Ramasamy, S.; Hariharan, S.; Venkatesan, V. Differential expression of microRNAs let-7a, miR-125b, miR-100, and miR-21 and interaction with NF-kB pathway genes in periodontitis pathogenesis. J. Cell Physiol. 2018, 233, 5877–5884. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Takahashi, H.; Okabe, H.; Ozaki, Y.; Hara, Y.; Kato, I. Distribution of natural killer cells in periodontal diseases: An immunohistochemical study. J. Periodontol. 1992, 63, 686–689. [Google Scholar] [CrossRef]
- Kamoda, Y.; Uematsu, H.; Yoshihara, A.; Miyazaki, H.; Senpuku, H. Role of activated natural killer cells in oral diseases. JPN J. Infect. Dis. 2008, 61, 469–474. [Google Scholar]
- Kawai, T.; Eisen-Lev, R.; Seki, M.; Eastcott, J.W.; Wilson, M.E.; Taubman, M.A. Requirement of B7 costimulation for Th1-mediated inflammatory bone resorption in experimental periodontal disease. J. Immunol. 2000, 164, 2102–2109. [Google Scholar] [CrossRef] [Green Version]
- Teng YT, A.; Nguyen, H.; Gao, X.; Kong, Y.Y.; Gorczynski, R.M.; Singh, B.; Ellen, R.P.; Penninger, J. M Functional human T-cell immunity and osteoprotegerin ligand control alveolar bone destruction in periodontal infection. J. Clin. Investig. 2000, 106, R59–R67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvi, G.E.; Brown, C.E.; Fujihashi, K.; Kiyono, H.; Smith, F.W.; Beck, J.D.; Offenbacher, S. Inflammatory mediators of the terminal dentition in adult and early onset periodontitis. J. Periodontal Res. 1998, 33, 212–225. [Google Scholar] [CrossRef]
- Stashenko, P.; Jandinski, J.J.; Fujiyoshi, P.; Rynar, J.; Socransky, S.S. Tissue levels of bone resorptive cytokines in periodontal disease. J. Periodontol. 1991, 62, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.J.; Dixon, M.; Evans, R.T.; Dufour, L.; Johnson, E.; Roopenian, D.C. CD4(+) T cells and the proinflammatory cytokines gamma interferon and interleukin-6 contribute to alveolar bone loss in mice. Infect. Immun. 1999, 67, 2804–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeichi, O.; Haber, J.; Kawai, T.; Smith, D.J.; Moro, I.; Taubman, M.A. Cytokine profiles of T-lymphocytes from gingival tissues with pathological pocketing. J. Dent. Res. 2000, 79, 1548–1555. [Google Scholar] [CrossRef]
- Page, R.C. Periodontal diseases: A new paradigm. J. Dent. Educ. 1998, 62, 812–821. [Google Scholar] [CrossRef]
- Page, R.C. The pathobiology of periodontal diseases may affect systemic diseases: Inversion of a paradigm. Ann. Periodontol. 1998, 3, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Hume, W.R.; Le, H.; Huynh, T.N.; Han, Y.W.; Cheng, G.; Shi, W. Induction of apoptotic cell death in peripheral blood mononuclear and polymorphonuclear cells by an oral bacterium, Fusobacterium nucleatum. Infect. Immun. 2000, 68, 1893–1898. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Bonavida, B. Target-induced inactivation and cell death by apoptosis in a subset of human NK cells. J. Immunol. 1996, 156, 907–915. [Google Scholar]
- Jewett, A.; Wang, M.Y.; Teruel, A.; Poupak, Z.; Bostanian, Z.; Park, N.H. Cytokine dependent inverse regulation of CD54 (ICAM1) and major histocompatibility complex class I antigens by nuclear factor kappaB in HEp2 tumor cell line: Effect on the function of natural killer cells. Hum. Immunol. 2003, 64, 505–520. [Google Scholar] [CrossRef]
- Jewett, A.; Cavalcanti, M.; Bonavida, B. Pivotal role of endogenous TNF-alpha in the induction of functional inactivation and apoptosis in NK cells. J. Immunol. 1997, 159, 4815–4822. [Google Scholar] [PubMed]
- Jewett, A.; Bonavida, B. Interferon-alpha activates cytotoxic function but inhibits interleukin-2-mediated proliferation and tumor necrosis factor-alpha secretion by immature human natural killer cells. J. Clin. Immunol. 1995, 15, 35–44. [Google Scholar] [CrossRef]
- Van Antwerp, D.J.; Martin, S.J.; Kafri, T.; Green, D.R.; Verma, I.M. Suppression of TNF-alpha-induced apoptosis by NF-kappaB. Science 1996, 274, 787–789. [Google Scholar] [CrossRef]
- Jewett, A.; Cacalano, N.A.; Teruel, A.; Romero, M.; Rashedi, M.; Wang, M.; Nakamura, H. Inhibition of nuclear factor kappa B (NFkappaB) activity in oral tumor cells prevents depletion of NK cells and increases their functional activation. Cancer Immunol. Immunother. 2006, 55, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.E.; Vaidya, S.A.; O′Connell, R.; Dadgostar, H.; Dempsey, P.W.; Wu, T.T.; Rao, G.; Sun, R.; Haberland, M.E.; Modlin, R.L.; et al. IRF3 mediates a TLR3/TLR4-specific antiviral gene program. Immunity 2002, 17, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Bartold, P.M.; Clayden, A.M.; Gao, J.; Haase, H.; Li, H.; Stevens, M.; Symsons, A.; Young, W.G.; Zhang, C.Z. The role of growth factors in periodontal and pulpal regeneration. JNZ Soc. Periodontol. 1998, 83, 7–14. [Google Scholar]
- Jotwani, R.; Cutler, C.W. Adult periodontitis--specific bacterial infection or chronic inflammation? J. Med. Microbiol. 1998, 47, 187–188. [Google Scholar] [PubMed]
- Graves, D.T.; Cochran, D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef]
- Kaur, K.; Ko, M.W.; Ohanian, N.; Cook, J.; Jewett, A. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Sci. Rep. 2020, 10, 20363. [Google Scholar] [CrossRef]
- Sawa, T.; Nishimura, F.; Ohyama, H.; Takahashi, K.; Takashiba, S.; Murayama, Y. In vitro induction of activation-induced cell death in lymphocytes from chronic periodontal lesions by exogenous Fas ligand. Infect Immun. 1999, 67, 1450–1454. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kawabata, K.; Fujihashi, K.; McGhee, J.R.; Van Dyke, T.E.; Bamberg, T.V.; Hiroi, T.; Kiyono, H. Absence of exogenous interleukin-4-induced apoptosis of gingival macrophages may contribute to chronic inflammation in periodontal diseases. Am. J. Pathol. 1996, 148, 331–339. [Google Scholar]
- Tseng, H.C.; Cacalano, N.; Jewett, A. Split anergized Natural Killer cells halt inflammation by inducing stem cell differentiation, resistance to NK cell cytotoxicity and prevention of cytokine and chemokine secretion. Oncotarget 2015, 6, 8947–8959. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Arasteh, A.; Tseng, H.C.; Behel, A.; Arasteh, H.; Yang, W.; Nicholas, A.; Paranjpe, A. Strategies to rescue mesenchymal stem cells (MSCs) and dental pulp stem cells (DPSCs) from NK cell mediated cytotoxicity. PLoS ONE 2010, 5, e9874. [Google Scholar] [CrossRef] [PubMed]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef]
- Ottsjö, L.S.; Flach, C.F.; Nilsson, S.; de Waal Malefyt, R.; Walduck, A.K.; Raghavan, S. Defining the Roles of IFN-γ and IL-17A in Inflammation and Protection against Helicobacter pylori Infection. PLoS ONE 2015, 10, e0131444. [Google Scholar]
- Jewett, A.; Cacalano, N.A.; Head, C.; Teruel, A. Coengagement of CD16 and CD94 receptors mediates secretion of chemokines and induces apoptotic death of naive natural killer cells. Clin. Cancer Res. 2006, 12, 1994–2003. [Google Scholar] [CrossRef] [Green Version]
- Tseng, H.-C.; Bui, V.; Man, Y.-G.; Cacalano, N.; Jewett, A. Induction of Split Anergy Conditions Natural Killer Cells to Promote Differentiation of Stem Cells through Cell-Cell Contact and Secreted Factors. Front. Immunol. 2014, 5, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewett, A.; Kos, J.; Kaur, K.; Safaei, T.; Sutanto, C.; Chen, W.; Wang, P.; Namagerdi, A.K.; Fang, C.; Ko, M.-W.; et al. Natural Killer Cells: Diverse Functions in Tumor Immunity and Defects in Pre-neoplastic and Neoplastic Stages of Tumorigenesis. Mol. Ther. Oncolytics 2020, 16, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Jewett, A.; Tseng, H.C.; Arasteh, A.; Saadat, S.; Christensen, R.E.; Cacalano, N.A. Natural killer cells preferentially target cancer stem cells; role of monocytes in protection against NK cell mediated lysis of cancer stem cells. Curr. Drug Deliv. 2012, 9, 5–16. [Google Scholar] [CrossRef]
- Jewett, A.; Man, Y.G.; Tseng, H.C. Dual functions of natural killer cells in selection and differentiation of stem cells; role in regulation of inflammation and regeneration of tissues. J. Cancer 2013, 4, 12–24. [Google Scholar] [CrossRef]
- Cappuyns, I.; Gugerli, P.; Mombelli, A. Viruses in periodontal disease—a review. Oral Dis. 2005, 11, 219–229. [Google Scholar] [CrossRef]
- Kubar, A.; Saygun, I.; Ozdemir, A.; Yapar, M.; Slots, J. Real-time polymerase chain reaction quantification of human cytomegalovirus and Epstein-Barr virus in periodontal pockets and the adjacent gingiva of periodontitis lesions. J. Periodontal Res. 2005, 40, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.J.; Ho, C.C.; Wu, C.Y.; Chen, Y.T.; Hung, S.L. Association between human herpesviruses and the severity of periodontitis. J. Periodontol. 2004, 75, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Slots, J.; Kamma, J.J.; Sugar, C. The herpesvirus-Porphyromonas gingivalis-periodontitis axis. J. Periodontal Res. 2003, 38, 318–323. [Google Scholar] [CrossRef]
- Dybing, J.K.; Walters, N.; Pascual, D.W. Role of endogenous interleukin-18 in resolving wild-type and attenuated Salmonella typhimurium infections. Infect. Immun. 1999, 67, 6242–6248. [Google Scholar] [CrossRef] [Green Version]
- Pontarini, E.; Lucchesi, D.; Fossati-Jimack, L.; Coleby, R.; Tentorio, P.; Croia, C.; Bombardieri, M.; Mavilio, D. NK cell recruitment in salivary glands provides early viral control but is dispensable for tertiary lymphoid structure formation. J. Leukoc. Biol. 2019, 105, 589–602. [Google Scholar] [CrossRef]
- Owen, K.A.; Anderson, C.J.; Casanova, J.E. Salmonella Suppresses the TRIF-Dependent Type I Interferon Response in Macrophages. Mbio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenlee-Wacker, M.C.; Nauseef, W.M. IFN-gamma targets macrophage-mediated immune responses toward Staphylococcus aureus. J. Leukoc. Biol. 2017, 101, 751–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, B.G.; Van Dyke, T.E. The role of inflammation and genetics in periodontal disease. Periodontol 2000 2020, 83, 26–39. [Google Scholar] [CrossRef]
- Hassell, T.M.; Harris, E.L. Genetic influences in caries and periodontal diseases. Crit. Rev. Oral Biol. Med. 1995, 6, 319–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irfan, U.M.; Dawson, D.V.; Bissada, N.F. Epidemiology of periodontal disease: A review and clinical perspectives. J. Int. Acad. Periodontol. 2001, 3, 14–21. [Google Scholar]
- Kantaputra, P.N.; Bongkochwilawan, C.; Lubinsky, M.; Pata, S.; Kaewgahya, M.; Tong, H.J.; Cairns, J.R.K.; Guven, Y.; Chaisrisookumporn, N. Periodontal disease and FAM20A mutations. J. Hum. Genet. 2017, 62, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Kocher, T.; Sawaf, H.; Fanghänel, J.; Timm, R.; Meisel, P. Association between bone loss in periodontal disease and polymorphism of N-acetyltransferase (NAT2). J. Clin. Periodontol. 2002, 29, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Arasteh, A.; Kaur, K.; Kozlowska, A.; Topchyan, P.; Jewett, A. Differential Cytotoxicity but Augmented IFN-γ Secretion by NK Cells after Interaction with Monocytes from Humans, and Those from Wild Type and Myeloid-Specific COX-2 Knockout Mice. Front. Immunol. 2015, 6, 259. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Cook, J.; Park, S.H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Gies, V.; Korganow, A.S.; Guffroy, A. Primary Immunodeficiencies With Defects in Innate Immunity: Focus on Orofacial Manifestations. Front. Immunol. 2020, 11, 1065. [Google Scholar] [CrossRef]
- Raje, N.; Dinakar, C. Overview of Immunodeficiency Disorders. Immunol. Allergy Clin. 2015, 35, 599–623. [Google Scholar] [CrossRef] [Green Version]
- Javali, M.A.; Patil, V.; Ayesha, H. Periodontal disease as the initial oral manifestation of abdominal tuberculosis. Dent. Res. J. 2012, 9, 634–637. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD16+ | CD3+ | CD19+ | CD3 + CD4+ | CD3 + CD8+ | ||
|---|---|---|---|---|---|---|
| Healthy | Peripheral blood | 15.5 ± 3.5 ** | 74 ± 7 | 10.5 ± 3.5 * | 55 ±7 | 19 ± 9 |
| Oral blood | 15.8 ± 9.7 | 74 ± 6.9 | 7.8 ± 7.4 | 52 ± 6 | 22 ± 10 | |
| Patients | Peripheral blood | 3.5 ± 0.7 ** | 71 ± 9.9 | 24.5 ± 9 * | 51 ± 3 | 20 ± 1 |
| Oral blood | 16 ± 10 | 73 ± 15 | 6.7 ± 4 | 53 ± 2 | 20 ± 7 |
| CD16+ | CD3+ | CD19+ | CD3 + CD4+ | CD3 + CD8+ | |
|---|---|---|---|---|---|
| Healthy | 4.7 ± 5.0 | 80 ± 19 | 6.7 ± 5.9 | 62 ± 8 | 18 ± 10 |
| Patients | 5.05 ± 3.9 | 68 ± 23 | 27 ± 23 | 47 ± 10 | 21 ± 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, K.; Vaziri, S.; Romero-Reyes, M.; Paranjpe, A.; Jewett, A. Phenotypic and Functional Alterations of Immune Effectors in Periodontitis; A Multifactorial and Complex Oral Disease. J. Clin. Med. 2021, 10, 875. https://doi.org/10.3390/jcm10040875
Kaur K, Vaziri S, Romero-Reyes M, Paranjpe A, Jewett A. Phenotypic and Functional Alterations of Immune Effectors in Periodontitis; A Multifactorial and Complex Oral Disease. Journal of Clinical Medicine. 2021; 10(4):875. https://doi.org/10.3390/jcm10040875
Chicago/Turabian StyleKaur, Kawaljit, Shahram Vaziri, Marcela Romero-Reyes, Avina Paranjpe, and Anahid Jewett. 2021. "Phenotypic and Functional Alterations of Immune Effectors in Periodontitis; A Multifactorial and Complex Oral Disease" Journal of Clinical Medicine 10, no. 4: 875. https://doi.org/10.3390/jcm10040875