
A Comparative Study on Delivery of Externally Attached DNA by Papillomavirus VLPs and Pseudoviruses


Abstract
:1. Introduction
2. Material and Methods
2.1. L1 VLPs, L1/L2 VLPs, and PSV Production and Validation
2.2. Cells for Externally Attached DNA Delivery
2.3. External DNA Delivery Using VLPs and PSVs in In Vitro Cell Cultures
2.4. Fluorescent Detection and Quantification of Delivery Efficacy Using Flow Cytometry Analysis
2.5. VLP Delivery of Viral DNA for In Vivo Infection of Rabbits
2.6. Statistics
3. Results
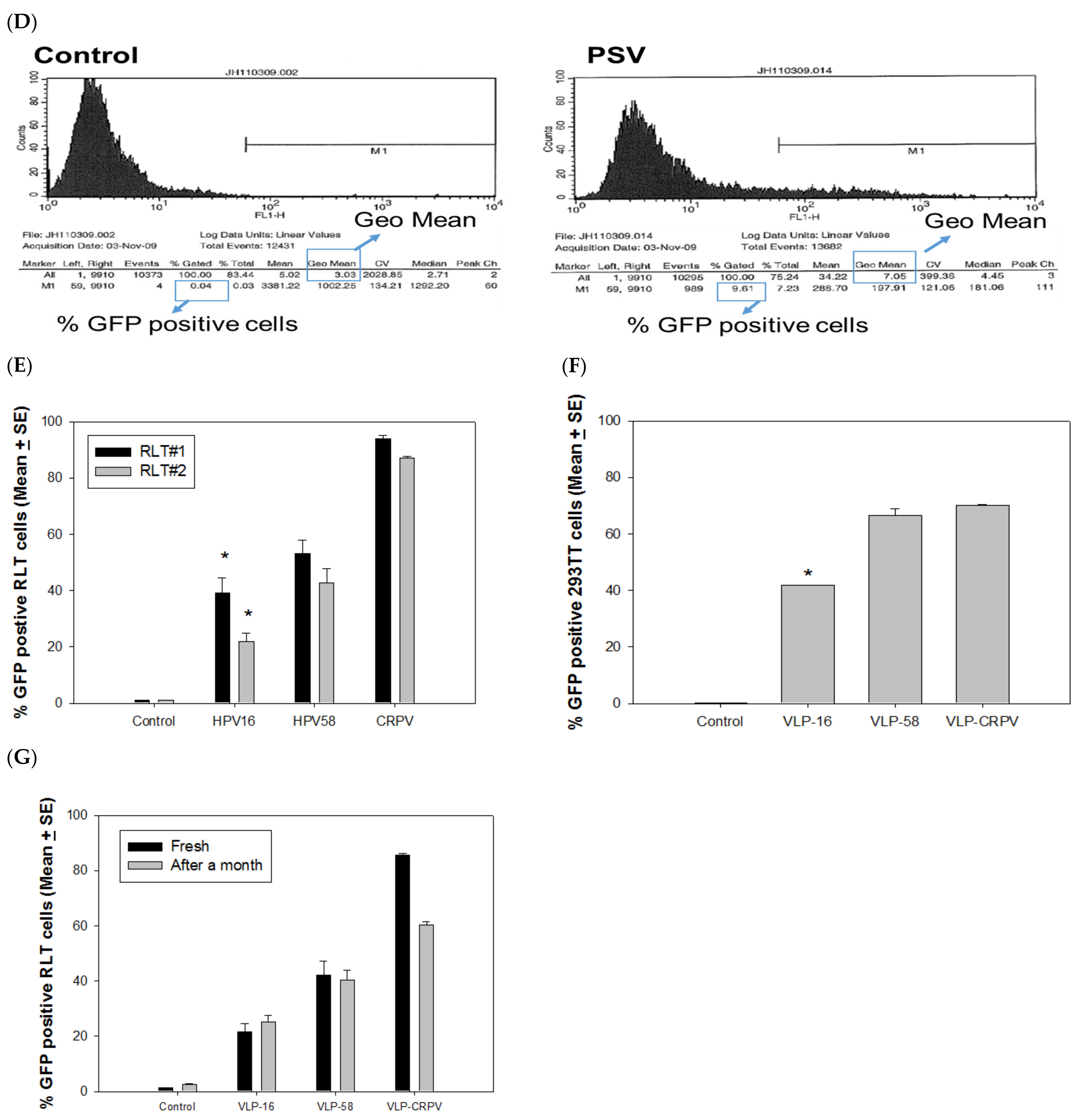
3.1. L2 Is Required for Delivering Externally Attached DNA to the Cells In Vitro
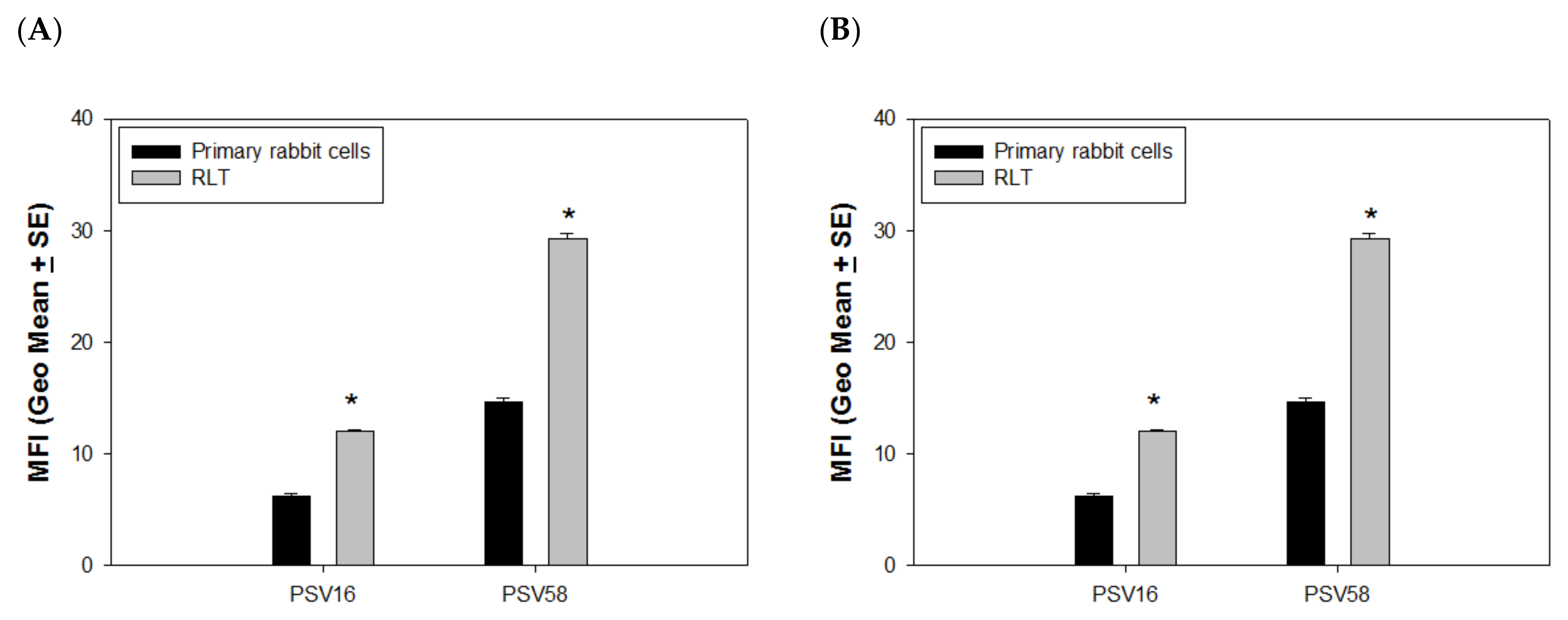
3.2. PSVs and VLPs Deliver Attached DNA to Both Rabbit and Human Cells
3.3. PSVs Were More Efficient in Delivering DNA When Encapisidating a Papillomavirus Genome DNA
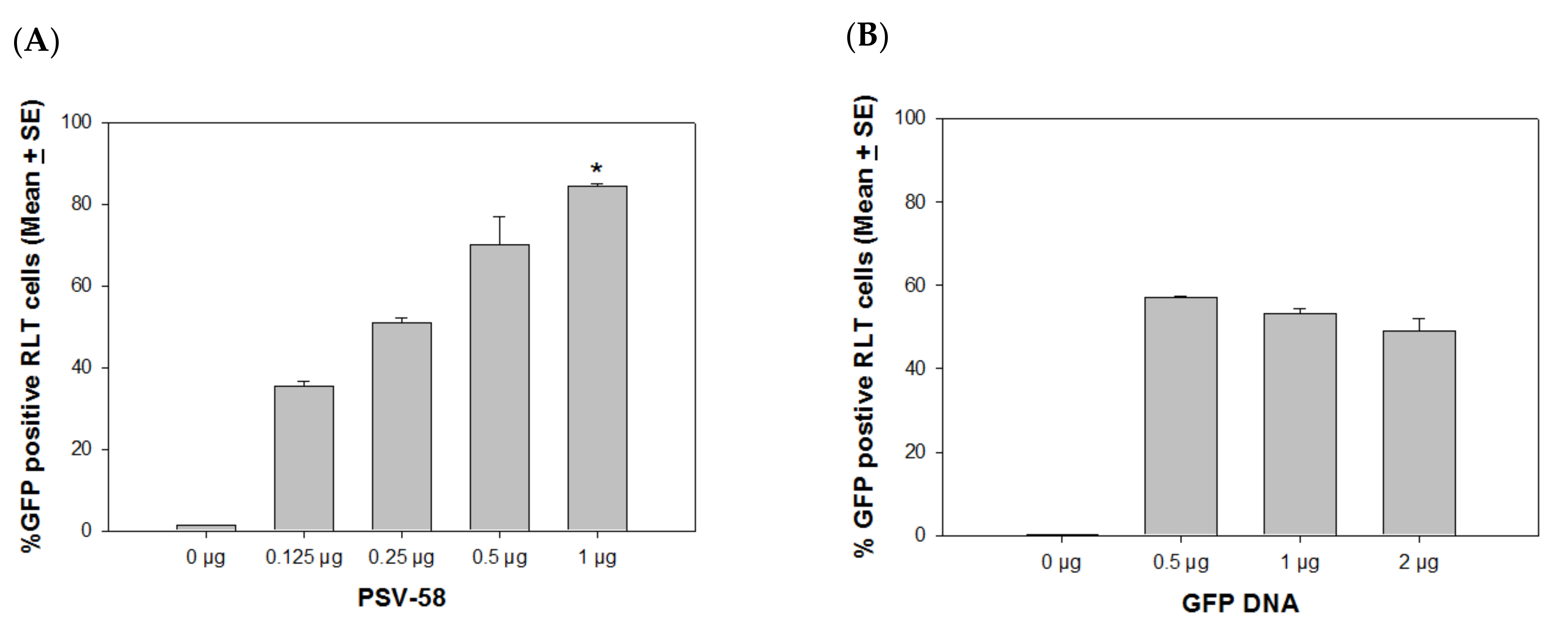
4. Optimal VLP/External DNA Ratio for Delivery
4.1. PSVs Deliver DNA More Efficiently in Immortalized Cells vs. Normal Cells
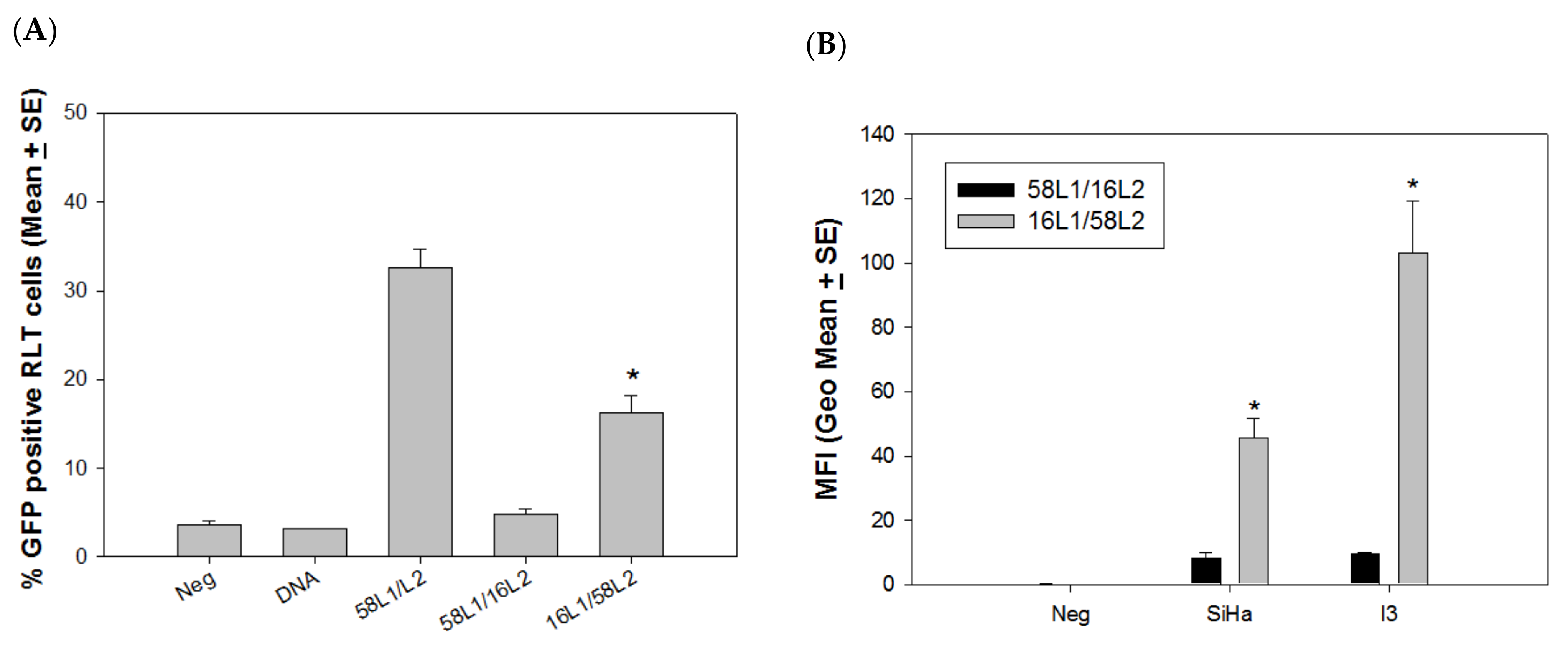
4.2. L2 Played a Critical Role in Delivery Efficiency
4.3. VLP-CRPV Increased Viral Infectivity In Vivo
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.W.; Roden, R.B. L2, the minor capsid protein of papillomavirus. Virology 2013, 445, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, C.B.; Day, P.M.; Trus, B.L. The papillomavirus major capsid protein L1. Virology 2013, 445, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, J.T.; Lowy, D.R. Papillomavirus-like particle vaccines. J. Natl. Cancer Inst. Monogr. 2001, 2000, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Dochez, C.; Bogers, J.J.; Verhelst, R.; Rees, H. HPV vaccines to prevent cervical cancer and genital warts: An update. Vaccine 2014, 32, 1595–1601. [Google Scholar] [CrossRef]
- Peng, S.; Frazer, I.H.; Fernando, G.J.; Zhou, J. Papillomavirus virus-like particles can deliver defined CTL epitopes to the MHC class I pathway. Virology 1998, 240, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Sun, X.-Y.; Louis, K.; Frazer, I.H. Interaction of human papillomavirus (HPV) type 16 capsid proteins with HPV DNA requires an intact L2 N-terminal sequence. J. Virol. 1994, 68, 619–625. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Gissmann, L.; Cristiano, R.J.; Sun, X.-Y.; Frazer, I.H.; Jenson, A.B.; Alonso, A.; Zentgraf, H.; Zhou, J. Papillomavirus capsid binding and uptake by cells from different tissues and species. J. Virol. 1995, 69, 948–954. [Google Scholar] [CrossRef] [Green Version]
- Kamper, N.; Day, P.M.; Nowak, T.; Selinka, H.C.; Florin, L.; Bolscher, J.; Hilbig, L.; Schiller, J.T.; Sapp, M. A membrane-destabilizing peptide in capsid protein L2 is required for egress of papillomavirus genomes from endosomes. J. Virol. 2006, 80, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Handisurya, A.; Day, P.M.; Thompson, C.D.; Buck, C.B.; Kwak, K.; Roden, R.B.; Lowy, D.R.; Schiller, J.T. Murine skin and vaginal mucosa are similarly susceptible to infection by pseudovirions of different papillomavirus classifications and species. Virology 2012, 433, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Kines, R.C.; Zarnitsyn, V.; Johnson, T.R.; Pang, Y.Y.; Corbett, K.S.; Nicewonger, J.D.; Gangopadhyay, A.; Chen, M.; Liu, J.; Prausnitz, M.R.; et al. Vaccination with human papillomavirus pseudovirus-encapsidated plasmids targeted to skin using microneedles. PLoS ONE 2015, 10, e0120797. [Google Scholar] [CrossRef]
- Cuburu, N.; Graham, B.S.; Buck, C.B.; Kines, R.C.; Pang, Y.Y.; Day, P.M.; Lowy, D.R.; Schiller, J.T. Intravaginal immunization with HPV vectors induces tissue-resident CD8+ T cell responses. J. Clin. Invest. 2012, 122, 4606–4620. [Google Scholar] [CrossRef]
- Mitsunaga, M.; Kosaka, N.; Kines, R.C.; Roberts, J.N.; Lowy, D.R.; Schiller, J.T.; Ishihara, Y.; Hasegawa, A.; Choyke, P.L.; Kobayashi, H. In vivo longitudinal imaging of experimental human papillomavirus infection in mice with a multicolor fluorescence mini-endoscopy system. Cancer Prev. Res. 2011, 4, 767–773. [Google Scholar] [CrossRef] [Green Version]
- Malboeuf, C.M.; Simon, D.A.; Lee, Y.E.; Lankes, H.A.; Dewhurst, S.; Frelinger, J.G.; Rose, R.C. Human papillomavirus-like particles mediate functional delivery of plasmid DNA to antigen presenting cells in vivo. Vaccine 2007, 25, 3270–3276. [Google Scholar] [CrossRef]
- Broutian, T.R.; Brendle, S.A.; Christensen, N.D. Differential binding patterns to host cells associated with particles of several human alphapapillomavirus types. J. Gen Virol. 2010, 91 Pt 2, 531–540. [Google Scholar] [CrossRef]
- Touze, A.; Coursaget, P. In vitro gene transfer using human papillomavirus-like particles. Nucleic Acids Res. 1998, 26, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Gissmann, L.; Sun, X.Y.; Kanjanahaluethai, A.; Muller, M.; Doorbar, J.; Zhou, J. Sequence close to the N-terminus of L2 protein is displayed on the surface of bovine papillomavirus type 1 virions. Virology 1997, 227, 474. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, S.C.; Patterson, N.A.; Ozbun, M.A.; Lambert, P.F. The minor capsid protein L2 contributes to two steps in the human papillomavirus type 31 life cycle. J. Virol. 2005, 79, 3938–3948. [Google Scholar] [CrossRef] [Green Version]
- Campos, S.K.; Ozbun, M.A. Two highly conserved cysteine residues in HPV16 L2 form an intramolecular disulfide bond and are critical for infectivity in human keratinocytes. PLoS ONE 2009, 4, e4463. [Google Scholar] [CrossRef] [Green Version]
- Culp, T.D.; Cladel, N.M.; Balogh, K.K.; Budgeon, L.R.; Mejia, A.F.; Christensen, N.D. Papillomavirus particles assembled in 293TT cells are infectious in vivo. J. Virol. 2006, 80, 11381–11384. [Google Scholar] [CrossRef] [Green Version]
- Goetschius, D.J.; Hartmann, S.R.; Subramanian, S.; Bator, C.M.; Christensen, N.D.; Hafenstein, S.L. High resolution cryo EM analysis of HPV16 identifies minor structural protein L2 and describes capsid flexibility. Sci. Rep. 2021, 11, 3498. [Google Scholar] [CrossRef]
- Schell, T.D.; Lippolis, J.D.; Tevethia, S.S. Cytotoxic T lymphocytes from HLA-A2.1 transgenic mice define a potential human epitope from simian virus 40 large T antigen. Cancer Res. 2001, 61, 873–879. [Google Scholar]
- Hu, J.; Peng, X.; Schell, T.D.; Budgeon, L.R.; Cladel, N.M.; Christensen, N.D. An HLA-A2.1-Transgenic Rabbit Model to Study Immunity to Papillomavirus Infection. J. Immunol. 2006, 177, 8037–8045. [Google Scholar] [CrossRef] [Green Version]
- Cladel, N.M.; Hu, J.; Balogh, K.; Mejia, A.; Christensen, N.D. Wounding prior to challenge substantially improves infectivity of cottontail rabbit papillomavirus and allows for standardization of infection. J. Virol. Methods 2008, 148, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Roden, R.B.; Kirnbauer, R.; Jenson, A.B.; Lowy, D.R.; Schiller, J.T. Interaction of papillomaviruses with the cell surface. J. Virol. 1994, 68, 7260–7266. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.N.; Buck, C.B.; Thompson, C.D.; Kines, R.; Bernardo, M.; Choyke, P.L.; Lowy, D.R.; Schiller, J.T. Genital transmission of HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat. Med. 2007, 13, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.N.; Sun, X.Y.; Frazer, I.H.; Zhou, J. DNA packaging by L1 and L2 capsid proteins of bovine papillomavirus type 1. Virology 1998, 243, 482–491. [Google Scholar] [CrossRef]
- Roden, R.B.; Greenstone, H.L.; Kirnbauer, R.; Booy, F.P.; Jessie, J.; Lowy, D.R.; Schiller, J.T. In vitro generation and type-specific neutralization of a human papillomavirus type 16 virion pseudotype. J. Virol. 1996, 70, 5875–5883. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Liu, J.; Huang, Y.; Qiao, L. Papillomavirus pseudovirus: A novel vaccine to induce mucosal and systemic cytotoxic T-lymphocyte responses. J. Virol. 2001, 75, 10139–10148. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Zhao, K.N.; Gao, F.G.; Leggatt, G.R.; Fernando, G.J.; Frazer, I.H. Polynucleotide viral vaccines: Codon optimisation and ubiquitin conjugation enhances prophylactic and therapeutic efficacy. Vaccine 2001, 20, 862–869. [Google Scholar] [CrossRef]
- Ohlschlager, P.; Osen, W.; Dell, K.; Faath, S.; Garcea, R.L.; Jochmus, I.; Muller, M.; Pawlita, M.; Schafer, K.; Sehr, P.; et al. Human papillomavirus type 16 L1 capsomeres induce L1-specific cytotoxic T lymphocytes and tumor regression in C57BL/6 mice. J. Virol. 2003, 77, 4635–4645. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.Y.; Frazer, I.; Müller, M.; Gissmann, L.; Zhou, J. Sequences required for the nuclear targeting and accumulation of human papillomavirus type 6B L2 protein. Virology 1995, 213, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touze, A.; Mahe, D.; El Mehdaoui, S.; Dupuy, C.; Combita-Rojas, A.L.; Bousarghin, L.; Sizaret, P.Y.; Coursaget, P. The nine C-terminal amino acids of the major capsid protein of the human papillomavirus type 16 are essential for DNA binding and gene transfer capacity. FEMS Microbiol. Lett. 2000, 189, 121–127. [Google Scholar] [CrossRef]
- Mach, H.; Volkin, D.B.; Troutman, R.D.; Wang, B.; Luo, Z.; Jansen, K.U.; Shi, L. Disassembly and reassembly of yeast-derived recombinant human papillomavirus virus-like particles (HPV VLPs). J. Pharm. Sci. 2006, 95, 2195–2206. [Google Scholar] [CrossRef] [PubMed]
- Garcea, R.L.; Gissmann, L. Virus-like particles as vaccines and vessels for the delivery of small molecules. Curr. Opin. Biotechnol. 2004, 15, 513–517. [Google Scholar] [CrossRef]
- Petry, H.; Goldmann, C.; Ast, O.; Luke, W. The use of virus-like particles for gene transfer. Curr. Opin. Mol. Ther. 2003, 5, 524–528. [Google Scholar]
- Schwarz, K.; Meijerink, E.; Speiser, D.E.; Tissot, A.C.; Cielens, I.; Renhof, R.; Dishlers, A.; Pumpens, P.; Bachmann, M.F. Efficient homologous prime-boost strategies for T cell vaccination based on virus-like particles. Eur. J. Immunol. 2005, 35, 816–821. [Google Scholar] [CrossRef]
- Yang, R.; Day, P.M.; Yutzy, W.H.; Lin, K.Y.; Hung, C.F.; Roden, R.B. Cell surface-binding motifs of L2 that facilitate papillomavirus infection. J. Virol. 2003, 77, 3531–3541. [Google Scholar] [CrossRef] [Green Version]
- Conway, M.J.; Alam, S.; Christensen, N.D.; Meyers, C. Overlapping and independent structural roles for human papillomavirus type 16 L2 conserved cysteines. Virology 2009, 393, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Kawana, K.; Yoshikawa, H.; Taketani, Y.; Yoshiike, K.; Kanda, T. Common neutralization epitope in minor capsid protein L2 of human papillomavirus types 16 and 6. J. Virol. 1999, 73, 6188–6190. [Google Scholar] [CrossRef] [Green Version]
- Rubio, I.; Seitz, H.; Canali, E.; Sehr, P.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Muller, M. The N-terminal region of the human papillomavirus L2 protein contains overlapping binding sites for neutralizing, cross-neutralizing and non-neutralizing antibodies. Virology 2011, 409, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Lowe, J.; Panda, D.; Rose, S.; Jensen, T.; Hughes, W.A.; Tso, F.Y.; Angeletti, P.C. Evolutionary and structural analyses of alpha-papillomavirus capsid proteins yields novel insights into L2 structure and interaction with L1. Virol. J. 2008, 5, 150. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.B.; Cheng, N.; Thompson, C.D.; Lowy, D.R.; Steven, A.C.; Schiller, J.T.; Trus, B.L. Arrangement of L2 within the papillomavirus capsid. J. Virol. 2008, 82, 5190–5197. [Google Scholar] [CrossRef] [Green Version]
- Meyers, C.; Bromberg-White, J.L.; Zhang, J.; Kaupas, M.E.; Bryan, J.T.; Lowe, R.S.; Jansen, K.U. Infectious virions produced from a human papillomavirus type 18/16 genomic DNA chimera. J. Virol. 2002, 76, 4723–4733. [Google Scholar] [CrossRef] [Green Version]
- Cladel, N.M.; Jiang, P.; Li, J.J.; Peng, X.; Cooper, T.K.; Majerciak, V.; Balogh, K.K.; Meyer, T.J.; Brendle, S.A.; Budgeon, L.R.; et al. Papillomavirus can be transmitted through the blood and produce infections in blood recipients: Evidence from two animal models. Emerg. Microbes Infect. 2019, 8, 1108–1121. [Google Scholar] [CrossRef] [Green Version]
- Greenstone, H.L.; Nieland, J.D.; de Visser, K.E.; De Bruijn, M.L.; Kirnbauer, R.; Roden, R.B.; Lowy, D.R.; Kast, W.M.; Schiller, J.T. Chimeric papillomavirus virus-like particles elicit antitumor immunity against the E7 oncoprotein in an HPV16 tumor model. Proc. Natl. Acad. Sci. USA 1998, 95, 1800–1805. [Google Scholar] [CrossRef] [Green Version]
- Fausch, S.C.; Da Silva, D.M.; Kast, W.M. Differential uptake and cross-presentation of human papillomavirus virus-like particles by dendritic cells and Langerhans cells. Cancer Res. 2003, 63, 3478–3482. [Google Scholar]
- Lenz, P.; Thompson, C.D.; Day, P.M.; Bacot, S.M.; Lowy, D.R.; Schiller, J.T. Interaction of papillomavirus virus-like particles with human myeloid antigen-presenting cells. Clin. Immunol. 2003, 106, 231–237. [Google Scholar] [CrossRef]
- Lenz, P.; Day, P.M.; Pang, Y.Y.; Frye, S.A.; Jensen, P.N.; Lowy, D.R.; Schiller, J.T. Papillomavirus-like particles induce acute activation of dendritic cells. J. Immunol. 2001, 166, 5346–5355. [Google Scholar] [CrossRef] [Green Version]
- Fausch, S.C.; Da Silva, D.M.; Kast, W.M. Heterologous papillomavirus virus-like particles and human papillomavirus virus-like particle immune complexes activate human Langerhans cells. Vaccine 2005, 23, 1720–1729. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | L1 | L2 | DNA Plasmid | Confirmation by ELISA | Notes |
|---|---|---|---|---|---|
| L1 VLPs | Yes | No | No | Antibodies used for L1: H16.V5 for HPV16L1, H58.G5 for H58L1, and CRPV.A4 for CRPV L1 L2: (R972 1:100) | All VLPs and PSVs were produced in 293TT cell culture, purified using OptiPrep gradient ultracentrifugation, and stored at −20 °C. For VLPs and PSVs, the positive L1 and L2 fractions were pooled, and concentrations were determined by Bio-Rad protein assay |
| VLPs (L1/L2) | Yes | Yes | No | ||
| PSVs | Yes | Yes | Yes, (pSEAP, 5.5 kb, PCX 5.5 kb, and the CRPV genome 7.8 kb) |
| Dose (ng) | Tumor Outgrowth (Tumor Sites/Challenge Sites) | Average Tumor Size (Geo Mean in mm) | |
|---|---|---|---|
| DNA | VLPs | ||
| 10 | - | 1/4 (25%) | 0.5000 |
| 10 | 20 | 1/2 (50%) | 3.0350 |
| 20 | - | 2/4 (50%) | 3.8675 |
| 20 | 40 | 2/2 (100%) | 7.8550 |
| 40 | - | 3/4 (75%) | 6.3750 |
| 40 | 80 | 2/2 (100%) | 7.2500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brendle, S.; Cladel, N.; Balogh, K.; Alam, S.; Christensen, N.; Meyers, C.; Hu, J. A Comparative Study on Delivery of Externally Attached DNA by Papillomavirus VLPs and Pseudoviruses. Vaccines 2021, 9, 1501. https://doi.org/10.3390/vaccines9121501
Brendle S, Cladel N, Balogh K, Alam S, Christensen N, Meyers C, Hu J. A Comparative Study on Delivery of Externally Attached DNA by Papillomavirus VLPs and Pseudoviruses. Vaccines. 2021; 9(12):1501. https://doi.org/10.3390/vaccines9121501
Chicago/Turabian StyleBrendle, Sarah, Nancy Cladel, Karla Balogh, Samina Alam, Neil Christensen, Craig Meyers, and Jiafen Hu. 2021. "A Comparative Study on Delivery of Externally Attached DNA by Papillomavirus VLPs and Pseudoviruses" Vaccines 9, no. 12: 1501. https://doi.org/10.3390/vaccines9121501