
Immunogenicity and Protective Capacity of a Virus-like Particle Vaccine against Chlamydia trachomatis Type 3 Secretion System Tip Protein, CT584

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
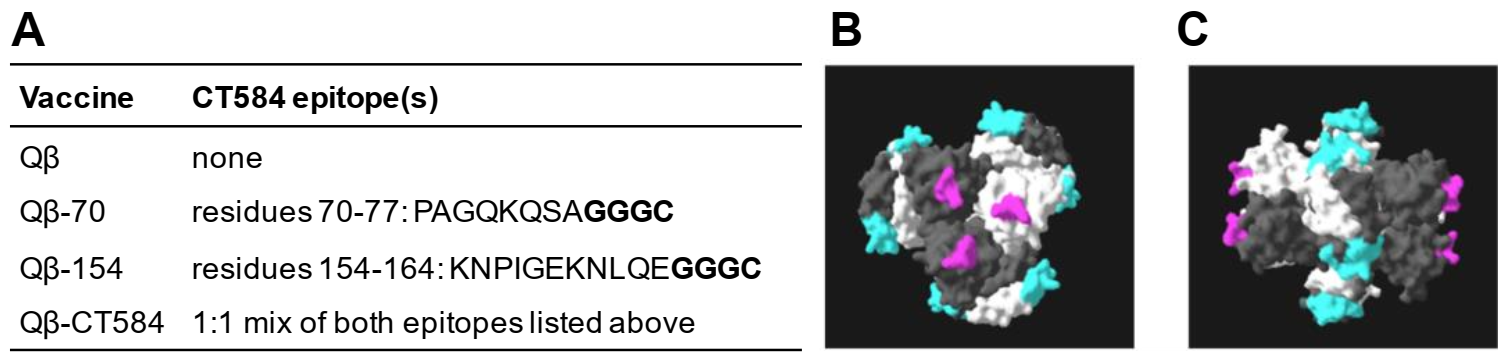
3.1. Creation of a VLP Vaccine against C. trachomatis CT584
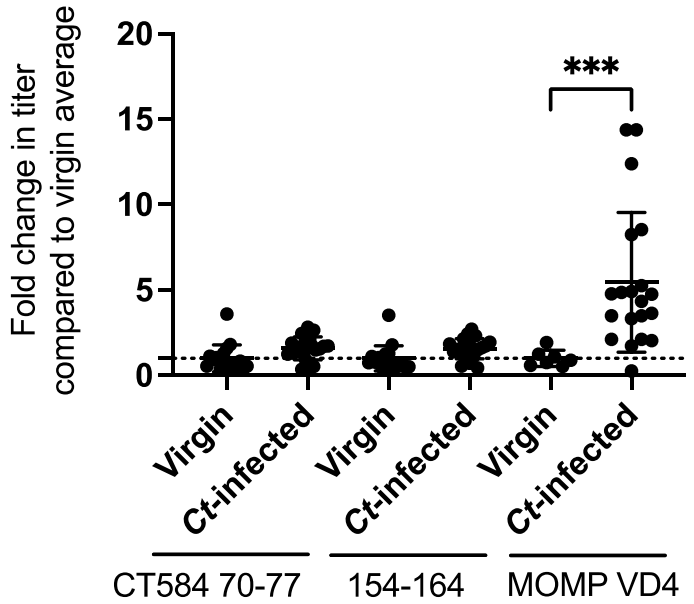
3.2. Urogenital C. trachomatis Infection in Women Does Not Elicit Antibody to Qβ-CT584 Vaccine Epitopes
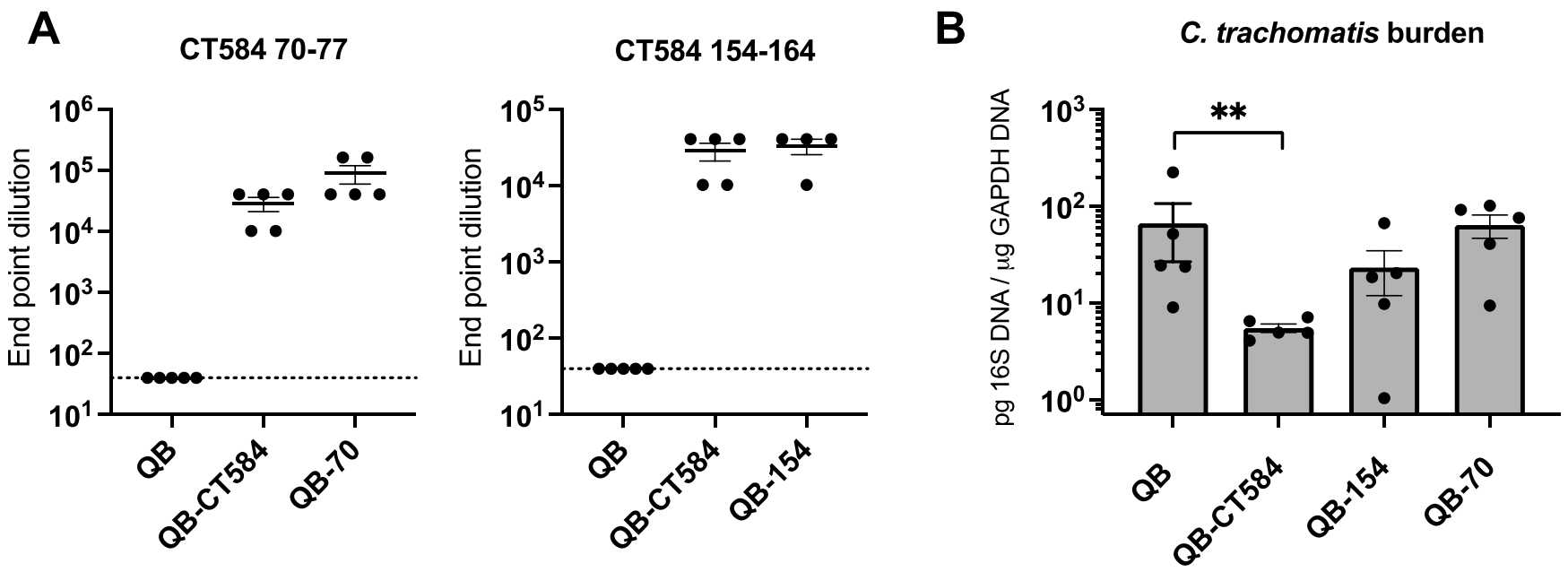
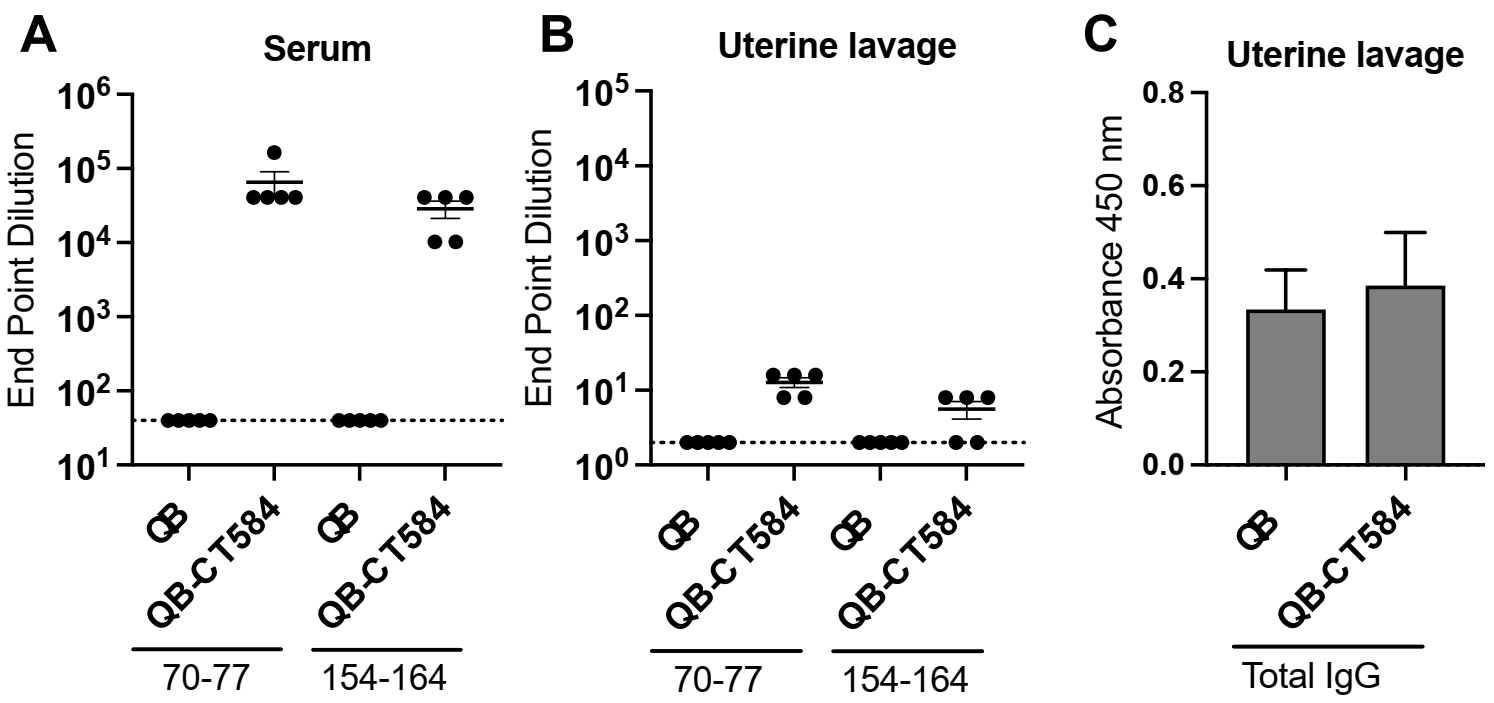
3.3. Immunization with Qβ-CT584 Induces High Titer, Peptide-Specific Serum IgG
3.4. Protection from C. trachomatis Infection after Immunization with Qβ-CT584 Varies
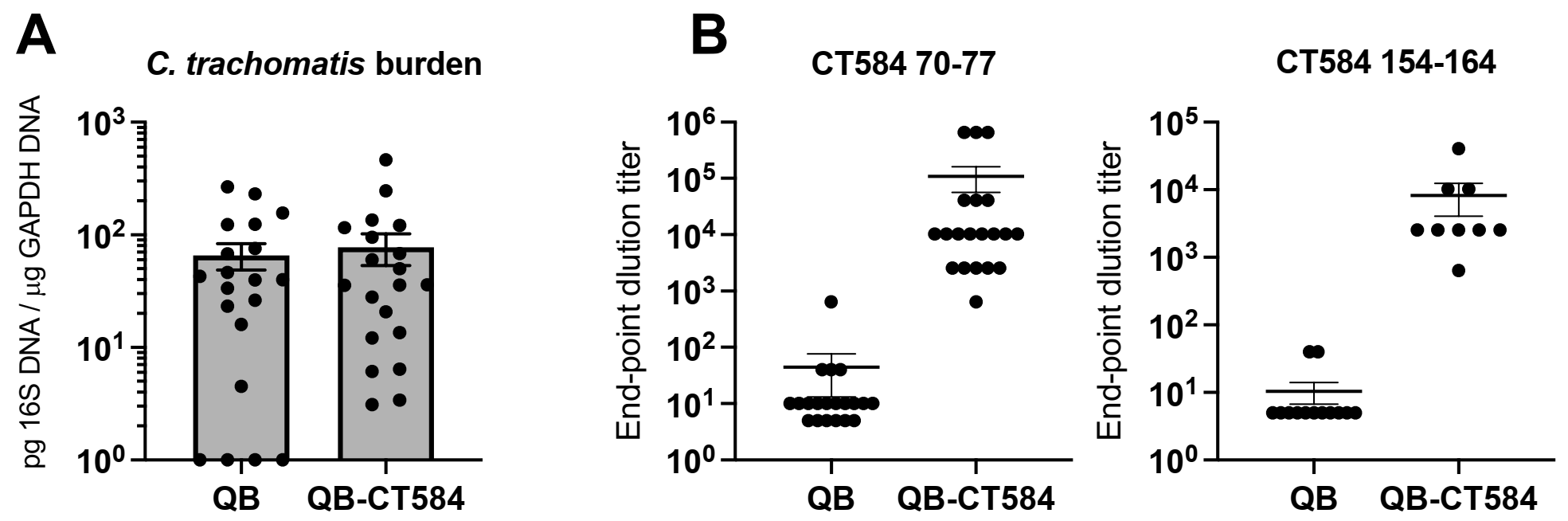
3.5. Vaccine-Induced Antibody Is Detectable at the Site of Infection
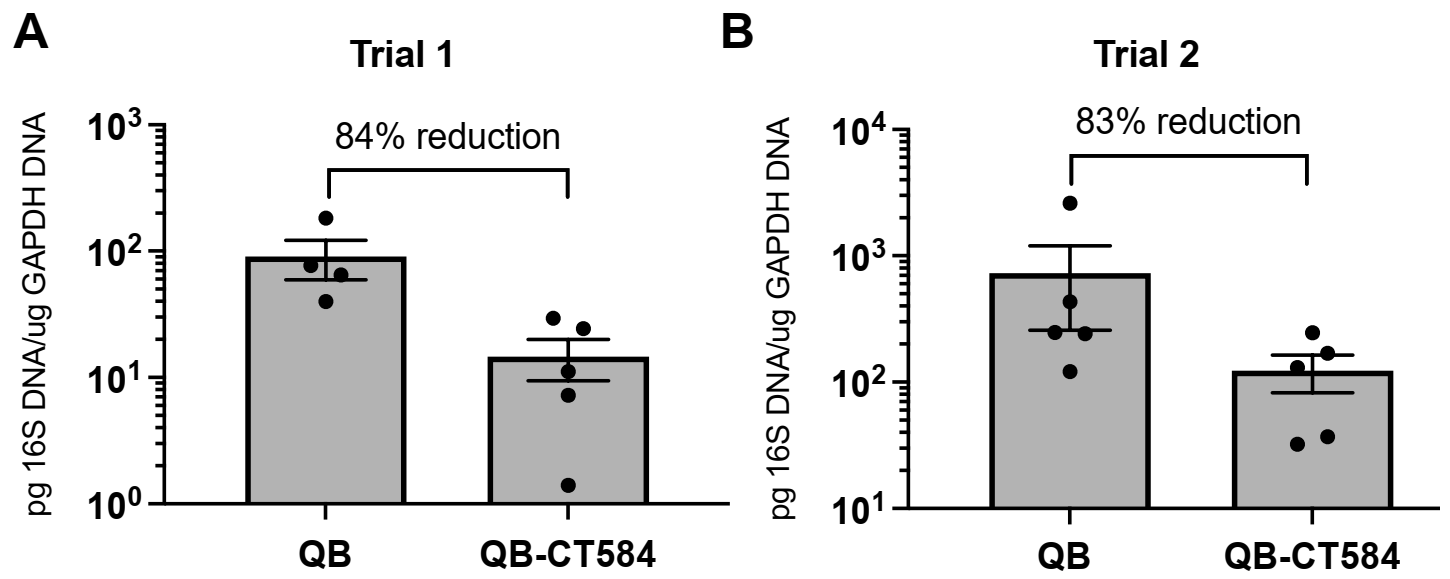
3.6. Pre-Incubation with Qβ-CT584-Immune Sera Reduces C. trachomatis Burden in the Upper Genital Tract
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rowley, J.; Vander Hoorn, S.; Korenromp, E.; Low, N.; Unemo, M.; Abu-Raddad, L.J.; Chico, R.M.; Smolak, A.; Newman, L.; Gottlieb, S.; et al. Chlamydia, Gonorrhoea, Trichomoniasis and Syphilis: Global Prevalence and Incidence Estimates, 2016. Bull. World Health Organ. 2019, 97, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia Cell Biology and Pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Darville, T.; Hiltke, T.J. Pathogenesis of Genital Tract Disease Due To Chlamydia trachomatis. J. Infect. Dis. 2010, 201, 114–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, M.; Sood, S.; Mukherjee, A.; Muralidhar, S.; Bala, M. Genital Chlamydia trachomatis: An Update. Indian J. Med. Res. 2013, 138, 303–316. [Google Scholar]
- Zhong, G.; Brunham, R.C.; de la Maza, L.M.; Darville, T.; Deal, C. National Institute of Allergy and Infectious Diseases Workshop Report: “Chlamydia Vaccines: The Way Forward”. Vaccine 2019, 37, 7346–7354. [Google Scholar] [CrossRef] [Green Version]
- Broutet, N.; Fruth, U.; Deal, C.; Gottlieb, S.L.; Rees, H. Consultation, participants of the 2013 S.V.T. Vaccines against Sexually Transmitted Infections: The Way Forward. Vaccine 2014, 32, 1630–1637. [Google Scholar] [CrossRef]
- Murray, S.M.; McKay, P.F. Chlamydia trachomatis: Cell Biology, Immunology and Vaccination. Vaccine 2021, 39, 2965–2975. [Google Scholar] [CrossRef]
- Abraham, S.; Juel, H.B.; Bang, P.; Cheeseman, H.M.; Dohn, R.B.; Cole, T.; Kristiansen, M.P.; Korsholm, K.S.; Lewis, D.; Olsen, A.W.; et al. Safety and Immunogenicity of the Chlamydia Vaccine Candidate CTH522 Adjuvanted with CAF01 Liposomes or Aluminium Hydroxide: A First-in-Human, Randomised, Double-Blind, Placebo-Controlled, Phase 1 Trial. Lancet Infect. Dis. 2019, 19, 1091–1100. [Google Scholar] [CrossRef]
- de la Maza, L.M.; Zhong, G.; Brunham, R.C. Update on Chlamydia trachomatis Vaccinology. Clin. Vaccine Immunol. I 2017, 24, e00543-16. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.W.; Follmann, F.; Erneholm, K.; Rosenkrands, I.; Andersen, P. Protection Against Chlamydia trachomatis Infection and Upper Genital Tract Pathological Changes by Vaccine-Promoted Neutralizing Antibodies Directed to the VD4 of the Major Outer Membrane Protein. J. Infect. Dis. 2015, 212, 978–989. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, Y.; Lu, C.; Lei, L.; Yu, P.; Zhong, G. A Genome-Wide Profiling of the Humoral Immune Response to Chlamydia trachomatis Infection Reveals Vaccine Candidate Antigens Expressed in Humans. J. Immunol. 2010, 185, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Collar, A.L.; Linville, A.C.; Core, S.B.; Wheeler, C.M.; Geisler, W.M.; Peabody, D.S.; Chackerian, B.; Frietze, K.M. Antibodies to Variable Domain 4 Linear Epitopes of the Chlamydia trachomatis Major Outer Membrane Protein Are Not Associated with Chlamydia Resolution or Reinfection in Women. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Batteiger, B.E.; Tu, W.; Ofner, S.; Van Der Pol, B.; Stothard, D.R.; Orr, D.P.; Katz, B.P.; Fortenberry, J.D. Repeated Chlamydia trachomatis Genital Infections in Adolescent Women. J. Infect. Dis. 2010, 201, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J.T. Papillomavirus L1 Major Capsid Protein Self-Assembles into Virus-like Particles That Are Highly Immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef] [Green Version]
- Stanley, M.; Pinto, L.A.; Trimble, C. Human Papillomavirus Vaccines–Immune Responses. Vaccine 2012, 30 (Suppl. 5), F83–F87. [Google Scholar] [CrossRef]
- Schiller, J.T.; Castellsagué, X.; Garland, S.M. A Review of Clinical Trials of Human Papillomavirus Prophylactic Vaccines. Vaccine 2012, 30 (Suppl. 5), F123–F138. [Google Scholar] [CrossRef] [Green Version]
- Longet, S.; Schiller, J.T.; Bobst, M.; Jichlinski, P.; Nardelli-Haefliger, D. A Murine Genital-Challenge Model Is a Sensitive Measure of Protective Antibodies against Human Papillomavirus Infection. J. Virol. 2011, 85, 13253–13259. [Google Scholar] [CrossRef] [Green Version]
- Suzich, J.A.; Ghim, S.J.; Palmer-Hill, F.J.; White, W.I.; Tamura, J.K.; Bell, J.A.; Newsome, J.A.; Jenson, A.B.; Schlegel, R. Systemic Immunization with Papillomavirus L1 Protein Completely Prevents the Development of Viral Mucosal Papillomas. Proc. Natl. Acad. Sci. USA 1995, 92, 11553–11557. [Google Scholar] [CrossRef] [Green Version]
- Breitburd, F.; Kirnbauer, R.; Hubbert, N.L.; Nonnenmacher, B.; Trin-Dinh-Desmarquet, C.; Orth, G.; Schiller, J.T.; Lowy, D.R. Immunization with Viruslike Particles from Cottontail Rabbit Papillomavirus (CRPV) Can Protect against Experimental CRPV Infection. J. Virol. 1995, 69, 3959–3963. [Google Scholar] [CrossRef] [Green Version]
- Scherpenisse, M.; Mollers, M.; Schepp, R.M.; Meijer, C.J.L.M.; de Melker, H.E.; Berbers, G.A.M.; van der Klis, F.R.M. Detection of Systemic and Mucosal HPV-Specific IgG and IgA Antibodies in Adolescent Girls One and Two Years after HPV Vaccination. Hum. Vaccines Immunother. 2013, 9, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Day, P.M.; Kines, R.C.; Thompson, C.D.; Jagu, S.; Roden, R.B.; Lowy, D.R.; Schiller, J.T. In Vivo Mechanisms of Vaccine-Induced Protection against HPV Infection. Cell Host Microbe 2010, 8, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Frietze, K.M.; Lijek, R.; Chackerian, B. Applying Lessons from Human Papillomavirus Vaccines to the Development of Vaccines against Chlamydia trachomatis. Expert Rev. Vaccines 2018, 17, 959–966. [Google Scholar] [CrossRef]
- Patel, K.G.; Swartz, J.R. Surface Functionalization of Virus-like Particles by Direct Conjugation Using Azide-Alkyne Click Chemistry. Bioconjug. Chem. 2011, 22, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, J.D.; Higginson, C.; Hovlid, M.L.; Kislukhin, A.A.; Castillejos, A.; Manzenrieder, F.; Campbell, M.G.; Voss, N.R.; Potter, C.S.; Carragher, B.; et al. Engineered Mutations Change the Structure and Stability of a Virus-like Particle. Biomacromolecules 2012, 13, 2339–2348. [Google Scholar] [CrossRef] [Green Version]
- Fasciano, A.C.; Shaban, L.; Mecsas, J. Promises and Challenges of the Type Three Secretion System Injectisome as an Antivirulence Target. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef]
- Yang, F.; Gu, J.; Yang, L.; Gao, C.; Jing, H.; Wang, Y.; Zeng, H.; Zou, Q.; Lv, F.; Zhang, J. Protective Efficacy of the Trivalent Pseudomonas aeruginosa Vaccine Candidate PcrV-OprI-Hcp1 in Murine Pneumonia and Burn Models. Sci. Rep. 2017, 7, 3957. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.M.; Lam, H.N.; Delgado, J.; Luu, J.; Mohammadi, S.; Isberg, R.R.; Wang, H.; Auerbuch, V. An Experimental Pipeline for Initial Characterization of Bacterial Type III Secretion System Inhibitor Mode of Action Using Enteropathogenic Yersinia. Front. Cell. Infect. Microbiol. 2018, 8, 404. [Google Scholar] [CrossRef]
- Martinez-Becerra, F.J.; Kumar, P.; Vishwakarma, V.; Kim, J.H.; Arizmendi, O.; Middaugh, C.R.; Picking, W.D.; Picking, W.L. Characterization and Protective Efficacy of Type III Secretion Proteins as a Broadly Protective Subunit Vaccine against Salmonella enterica Serotypes. Infect. Immun. 2018, 86, e00473-17. [Google Scholar] [CrossRef] [Green Version]
- Clifton, D.R.; Fields, K.A.; Grieshaber, S.S.; Dooley, C.A.; Fischer, E.R.; Mead, D.J.; Carabeo, R.A.; Hackstadt, T. A Chlamydial Type III Translocated Protein Is Tyrosine-Phosphorylated at the Site of Entry and Associated with Recruitment of Actin. Proc. Natl. Acad. Sci. USA 2004, 101, 10166–10171. [Google Scholar] [CrossRef] [Green Version]
- Markham, A.P.; Jaafar, Z.A.; Kemege, K.E.; Middaugh, C.R.; Hefty, P.S. Biophysical Characterization of Chlamydia trachomatis CT584 Supports Its Potential Role as a Type III Secretion Needle Tip Protein. Biochemistry 2009, 48, 10353–10361. [Google Scholar] [CrossRef] [Green Version]
- Barta, M.L.; Hickey, J.; Kemege, K.E.; Lovell, S.; Battaile, K.P.; Hefty, P.S. Structure of CT584 from Chlamydia trachomatis Refined to 3.05 Å Resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 1196–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulir, D.C.; Liang, S.; Lee, A.; Chong, S.; Simms, E.; Stone, C.; Kaushic, C.; Ashkar, A.; Mahony, J.B. Immunization with Chlamydial Type III Secretion Antigens Reduces Vaginal Shedding and Prevents Fallopian Tube Pathology Following Live C. muridarum Challenge. Vaccine 2016, 34, 3979–3985. [Google Scholar] [CrossRef] [PubMed]
- Warner, N.L.; Frietze, K.M. Development of Bacteriophage Virus-Like Particle Vaccines Displaying Conserved Epitopes of Dengue Virus Non-Structural Protein 1. Vaccines 2021, 9, 726. [Google Scholar] [CrossRef] [PubMed]
- Lijek, R.S.; Helble, J.D.; Olive, A.J.; Seiger, K.W.; Starnbach, M.N. Pathology after Chlamydia trachomatis Infection Is Driven by Nonprotective Immune Cells That Are Distinct from Protective Populations. Proc. Natl. Acad. Sci. USA 2018, 115, 2216–2221. [Google Scholar] [CrossRef] [Green Version]
- Gondek, D.C.; Olive, A.J.; Stary, G.; Starnbach, M.N. CD4+ T Cells Are Necessary and Sufficient to Confer Protection against Chlamydia trachomatis Infection in the Murine Upper Genital Tract. J. Immunol. 2012, 189, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Cheng, X.; de la Maza, L.M.; Peterson, E.M. Analysis of the Humoral Response Elicited in Mice by a Chimeric Peptide Representing Variable Segments I and IV of the Major Outer Membrane Protein of Chlamydia trachomatis. Vaccine 1994, 12, 557–564. [Google Scholar] [CrossRef]
- Ambühl, P.M.; Tissot, A.C.; Fulurija, A.; Maurer, P.; Nussberger, J.; Sabat, R.; Nief, V.; Schellekens, C.; Sladko, K.; Roubicek, K.; et al. A Vaccine for Hypertension Based on Virus-like Particles: Preclinical Efficacy and Phase I Safety and Immunogenicity. J. Hypertens. 2007, 25, 63–72. [Google Scholar] [CrossRef]
- Frietze, K.M.; Peabody, D.S.; Chackerian, B. Engineering Virus-like Particles as Vaccine Platforms. Curr. Opin. Virol. 2016, 18, 44–49. [Google Scholar] [CrossRef] [Green Version]
- IEDB.Org: Free Epitope Database and Prediction Resource. Available online: http://www.iedb.org (accessed on 24 November 2021).
- Bernstein, D.I.; Cardin, R.D.; Bravo, F.J.; Awasthi, S.; Lu, P.; Pullum, D.A.; Dixon, D.A.; Iwasaki, A.; Friedman, H.M. Successful Application of Prime and Pull Strategy for a Therapeutic HSV Vaccine. NPJ Vaccines 2019, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Roden, R.; Wu, T.-C. How Will HPV Vaccines Affect Cervical Cancer? Nat. Rev. Cancer 2006, 6, 753–763. [Google Scholar] [CrossRef]
- Cotter, T.W.; Meng, Q.; Shen, Z.L.; Zhang, Y.X.; Su, H.; Caldwell, H.D. Protective Efficacy of Major Outer Membrane Protein-Specific Immunoglobulin A (IgA) and IgG Monoclonal Antibodies in a Murine Model of Chlamydia trachomatis Genital Tract Infection. Infect. Immun. 1995, 63, 4704–4714. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.G.; Morrison, R.P. A Predominant Role for Antibody in Acquired Immunity to Chlamydial Genital Tract Reinfection. J. Immunol. 2005, 175, 7536–7542. [Google Scholar] [CrossRef] [Green Version]
- Farris, C.M.; Morrison, S.G.; Morrison, R.P. CD4+ T Cells and Antibody Are Required for Optimal Major Outer Membrane Protein Vaccine-Induced Immunity to Chlamydia muridarum Genital Infection. Infect. Immun. 2010, 78, 4374–4383. [Google Scholar] [CrossRef] [Green Version]
- Faure, K.; Fujimoto, J.; Shimabukuro, D.W.; Ajayi, T.; Shime, N.; Moriyama, K.; Spack, E.G.; Wiener-Kronish, J.P.; Sawa, T. Effects of Monoclonal Anti-PcrV Antibody on Pseudomonas aeruginosa-Induced Acute Lung Injury in a Rat Model. J. Immune Based Ther. Vaccines 2003, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- François, B.; Luyt, C.-E.; Dugard, A.; Wolff, M.; Diehl, J.-L.; Jaber, S.; Forel, J.-M.; Garot, D.; Kipnis, E.; Mebazaa, A.; et al. Safety and Pharmacokinetics of an Anti-PcrV PEGylated Monoclonal Antibody Fragment in Mechanically Ventilated Patients Colonized with Pseudomonas aeruginosa: A Randomized, Double-Blind, Placebo-Controlled Trial. Crit. Care Med. 2012, 40, 2320–2326. [Google Scholar] [CrossRef]
- Horna, G.; Ruiz, J. Type 3 Secretion System of Pseudomonas aeruginosa. Microbiol. Res. 2021, 246, 126719. [Google Scholar] [CrossRef]
- Jain, R.; Beckett, V.V.; Konstan, M.W.; Accurso, F.J.; Burns, J.L.; Mayer-Hamblett, N.; Milla, C.; VanDevanter, D.R.; Chmiel, J.F.; Group, K.-A.S. KB001-A, a Novel Anti-Inflammatory, Found to Be Safe and Well-Tolerated in Cystic Fibrosis Patients Infected with Pseudomonas aeruginosa. J. Cyst. Fibros. 2018, 17, 484–491. [Google Scholar] [CrossRef]
- Milla, C.E.; Chmiel, J.F.; Accurso, F.J.; VanDevanter, D.R.; Konstan, M.W.; Yarranton, G.; Geller, D.E.; Group, K.S. Anti-PcrV Antibody in Cystic Fibrosis: A Novel Approach Targeting Pseudomonas aeruginosa Airway Infection. Pediatr. Pulmonol. 2014, 49, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Shime, N.; Sawa, T.; Fujimoto, J.; Faure, K.; Allmond, L.R.; Karaca, T.; Swanson, B.L.; Spack, E.G.; Wiener-Kronish, J.P. Therapeutic Administration of Anti-PcrV F(Ab’)(2) in Sepsis Associated with Pseudomonas aeruginosa. J. Immunol. 2001, 167, 5880–5886. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, C.P.; Armitage, C.W.; Andrew, D.W.; Kollipara, A.; Lycke, N.Y.; Potter, A.A.; Gerdts, V.; Petrovsky, N.; Beagley, K.W. Multistage Vaccines Containing Outer Membrane, Type III Secretion System and Inclusion Membrane Proteins Protects against a Chlamydia Genital Tract Infection and Pathology. Vaccine 2017, 35, 3883–3888. [Google Scholar] [CrossRef]
- Crane, D.D.; Carlson, J.H.; Fischer, E.R.; Bavoil, P.; Hsia, R.; Tan, C.; Kuo, C.; Caldwell, H.D. Chlamydia trachomatis Polymorphic Membrane Protein D Is a Species-Common Pan-Neutralizing Antigen. Proc. Natl. Acad. Sci. USA 2006, 103, 1894–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozberk, V.; Pandey, M.; Good, M.F. Contribution of Cryptic Epitopes in Designing a Group A Streptococcal Vaccine. Hum. Vaccines Immunother. 2018, 14, 2034–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roden, R.B.; Yutzy, W.H.; Fallon, R.; Inglis, S.; Lowy, D.R.; Schiller, J.T. Minor Capsid Protein of Human Genital Papillomaviruses Contains Subdominant, Cross-Neutralizing Epitopes. Virology 2000, 270, 254–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.A.; Verrips, C.T. Nanobodies That Neutralize HIV. Vaccines 2019, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A Highly Conserved Cryptic Epitope in the Receptor Binding Domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Webster, E.; Seiger, K.W.; Core, S.B.; Collar, A.L.; Knapp-Broas, H.; Graham, J.; Shrestha, M.; Afzaal, S.; Geisler, W.M.; Wheeler, C.M.; et al. Immunogenicity and Protective Capacity of a Virus-like Particle Vaccine against Chlamydia trachomatis Type 3 Secretion System Tip Protein, CT584. Vaccines 2022, 10, 111. https://doi.org/10.3390/vaccines10010111
Webster E, Seiger KW, Core SB, Collar AL, Knapp-Broas H, Graham J, Shrestha M, Afzaal S, Geisler WM, Wheeler CM, et al. Immunogenicity and Protective Capacity of a Virus-like Particle Vaccine against Chlamydia trachomatis Type 3 Secretion System Tip Protein, CT584. Vaccines. 2022; 10(1):111. https://doi.org/10.3390/vaccines10010111
Chicago/Turabian StyleWebster, Everett, Kyra W. Seiger, Susan B. Core, Amanda L. Collar, Hannah Knapp-Broas, June Graham, Muskan Shrestha, Sarah Afzaal, William M. Geisler, Cosette M. Wheeler, and et al. 2022. "Immunogenicity and Protective Capacity of a Virus-like Particle Vaccine against Chlamydia trachomatis Type 3 Secretion System Tip Protein, CT584" Vaccines 10, no. 1: 111. https://doi.org/10.3390/vaccines10010111