
A Booster with a Genotype-Matched Inactivated Newcastle Disease Virus (NDV) Vaccine Candidate Provides Better Protection against a Virulent Genotype XIII.2 Virus

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Vaccine
2.2. Virus Inactivation and Preparation of NDV Inactivated Vaccines
2.3. Vaccination and Challenge
2.4. Sample Collection and Processing
2.5. Hemagglutination Inhibition Test
2.6. Detection of Virus Shedding by RT-qPCR
2.7. Sequencing
2.8. Statistical Analysis
3. Results
3.1. Antibody Response following Vaccination and Challenge
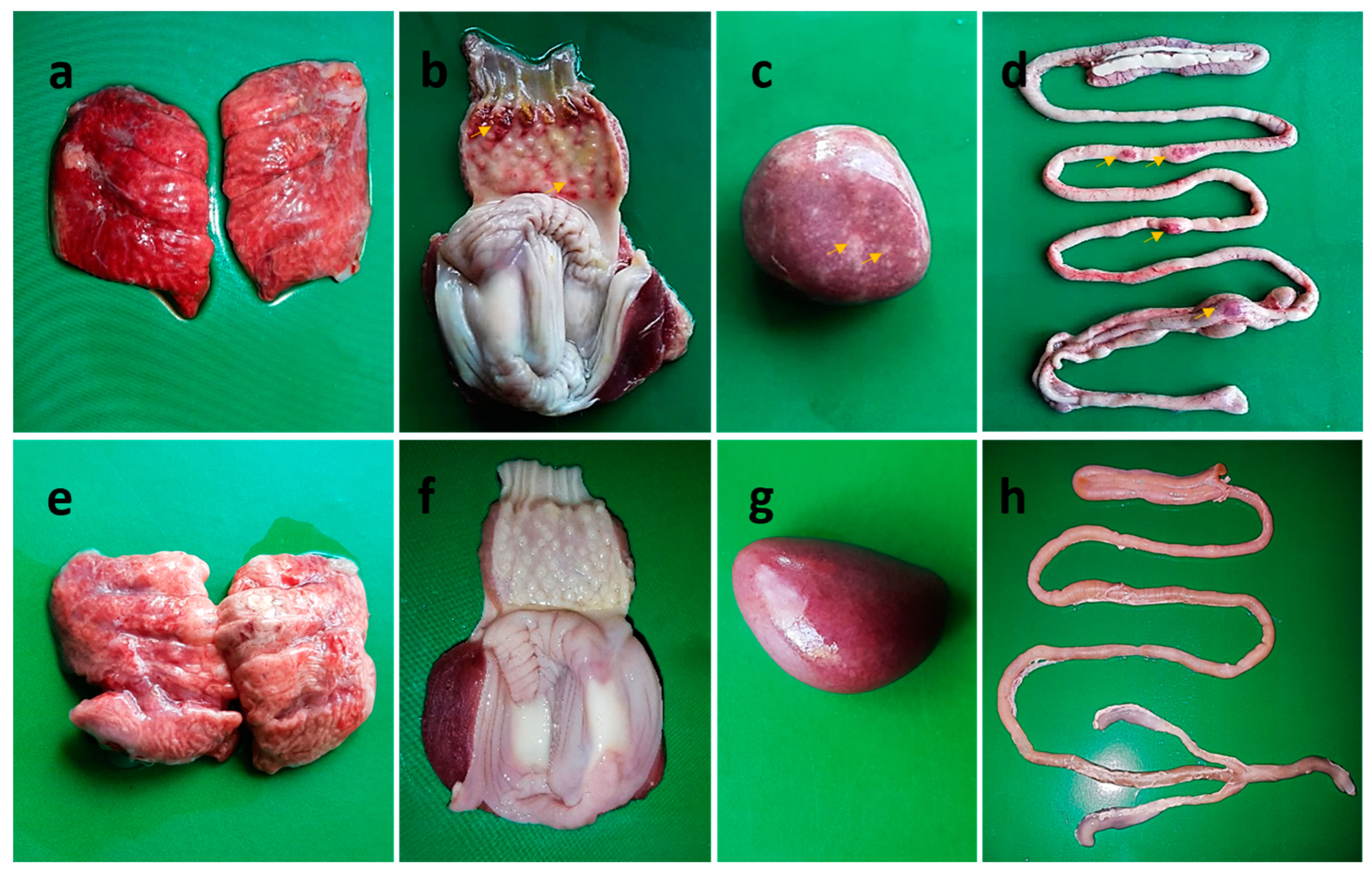
3.2. Clinical Signs, Mortality, and Postmortem Findings following Challenge
3.3. Virus Shedding following Challenge
3.4. Genetic Variability between LaSota and BD-C161/2010 NDV Strains at Putative Neutralizing Epitopes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kaleta, E.F.; Baldauf, C. Newcastle Disease in Free-Living and Pet Birds. In Newcastle Disease; Alexander, D.J., Ed.; Springer US: Boston, MA, USA, 1988; pp. 197–246. [Google Scholar]
- Barman, L.R.; Nooruzzaman, M.; Sarker, R.D.; Rahman, M.T.; Saife, M.R.B.; Giasuddin, M.; Das, B.C.; Das, P.M.; Chowdhury, E.H.; Islam, M.R. Phylogenetic analysis of Newcastle disease viruses from Bangladesh suggests continuing evolution of genotype XIII. Arch. Virol. 2017, 162, 3177–3182. [Google Scholar] [CrossRef]
- Belgrad, J.P.; Rahman, M.A.; Abdullah, M.S.; Rashid, M.H.; Sayeed, M.A.; Anwer, M.S.; Hoque, M.A. Newcastle disease sero and viro-prevalence in rural poultry in Chittagong, Bangladesh. Prev. Vet. Med. 2018, 160, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Nooruzzaman, M.; Barman, L.R.; Mumu, T.T.; Chowdhury, E.H.; Dimitrov, K.M.; Islam, M.R. A pigeon-derived sub-genotype XXI.1.2 Newcastle disease virus from Bangladesh induces high mortality in chickens. Viruses 2021, 13, 1520. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Nooruzzaman, M.; Kabiraj, C.K.; Mumu, T.T.; Das, P.M.; Chowdhury, E.H.; Islam, M.R. Surveillance on respiratory diseases reveals enzootic circulation of both H5 and H9 avian influenza viruses in small-scale commercial layer farms of Bangladesh. Zoonoses Public Health 2021, 68, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.C.; Khatun, S.; Nooruzzaman, M.; Chowdhury, E.H.; Islam, M.R. Isolation and identification of Newcastle disease viruses from field outbreaks in chickens and pigeons. Bangladesh Vet. 2012, 29, 41–48. [Google Scholar] [CrossRef]
- Roky, S.A.; Das, M.; Akter, S.; Islam, A.; Paul, S. Determinants of Newcastle disease in commercial layer chicken farms in two districts of Bangladesh: A case-control study. Heliyon 2022, 8, e10229. [Google Scholar] [CrossRef]
- Nooruzzaman, M.; Mumu, T.T.; Kabiraj, C.K.; Hasnat, A.; Rahman, M.M.; Chowdhury, E.H.; Dimitrov, K.M.; Islam, M.R. Genetic and biological characterization of Newcastle disease viruses circulating in Bangladesh during 2010–2017: Further genetic diversification of class II genotype XIII in Southcentral Asia. J. Gen. Virol. 2021, 102, 001554. [Google Scholar] [CrossRef]
- Nooruzzaman, M.; Hossain, I.; Begum, J.A.; Moula, M.; Khaled, S.A.; Parvin, R.; Chowdhury, E.H.; Islam, M.R.; Diel, D.G.; Dimitrov, K.M. The first report of a virulent Newcastle disease virus of genotype VII.2 causing outbreaks in chickens in Bangladesh. Viruses 2022, 14, 2627. [Google Scholar] [CrossRef]
- Hossain, I.; Parvin, R.; Rahman, M.M.; Begum, J.A.; Chowdhury, E.H.; Islam, M.R.; Diel, D.G.; Nooruzzaman, M. Comparative pathogenicity of a genotype XXI.1.2 pigeon Newcastle disease virus isolate in pigeons and chickens. Microb. Pathog. 2023, 178, 106068. [Google Scholar] [CrossRef]
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P.; et al. ICTV Virus Taxonomy Profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593–1594. [Google Scholar] [CrossRef]
- Meulemans, G.; Gonze, M.; Carlier, M.C.; Petit, P.; Burny, A.; Long, L. Protective effects of HN and F glycoprotein-specific monoclonal antibodies on experimental Newcastle disease. Avian Pathol. 1986, 15, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Boursnell, M.E.; Green, P.F.; Samson, A.C.; Campbell, J.I.; Deuter, A.; Peters, R.W.; Millar, N.S.; Emmerson, P.T.; Binns, M.M. A recombinant fowlpox virus expressing the hemagglutinin-neuraminidase gene of Newcastle disease virus (NDV) protects chickens against challenge by NDV. Virology 1990, 178, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Cosset, F.L.; Bouquet, J.F.; Drynda, A.; Chebloune, Y.; Rey-Senelonge, A.; Kohen, G.; Nigon, V.M.; Desmettre, P.; Verdier, G. Newcastle disease virus (NDV) vaccine based on immunization with avian cells expressing the NDV hemagglutinin-neuraminidase glycoprotein. Virology 1991, 185, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Karaca, K.; Sharma, J.M.; Winslow, B.J.; Junker, D.E.; Reddy, S.; Cochran, M.; McMillen, J. Recombinant fowlpox viruses coexpressing chicken type I IFN and Newcastle disease virus HN and F genes: Influence of IFN on protective efficacy and humoral responses of chickens following in ovo or post-hatch administration of recombinant viruses. Vaccine 1998, 16, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.L.; Wang, Y.F.; Tong, G.Z.; Zhang, P.J.; Miao, D.Y.; Zhi, H.D.; Wang, M.; Wang, M. Protection of chickens from Newcastle disease and infectious laryngotracheitis with a recombinant fowlpox virus co-expressing the F, HN genes of Newcastle disease virus and gB gene of infectious laryngotracheitis virus. Avian Dis. 2008, 52, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Wei, Q.; Bi, Y.; Li, Y.; Huo, N.; Mou, S.; Wang, W.; Liu, H.; Yang, Z.; Chen, H.; et al. Identification of a potential neutralizing linear epitope of hemagglutinin-neuraminidase in Newcastle disease virus. Virology 2021, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- OIE. OIE Terrestrial Mannual 2018; World Organization for Animal Health: Paris, France, 2018; Chapter 3.3.14. Newcastle Disease (Infection with Newcastle Disease Virus); pp. 964–983. [Google Scholar]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Diagnostic and vaccination approaches for Newcastle disease virus in poultry: The current and emerging perspectives. BioMed Res. Int. 2018, 2018, 7278459. [Google Scholar] [CrossRef]
- Alexander, D.J. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech. 2000, 19, 443–462. [Google Scholar] [CrossRef]
- Lindh, E.; Ek-Kommonen, C.; Väänänen, V.M.; Alasaari, J.; Vaheri, A.; Vapalahti, O.; Huovilainen, A. Molecular epidemiology of outbreak-associated and wild-waterfowl-derived newcastle disease virus strains in Finland, including a novel class I genotype. J. Clin. Microbiol. 2012, 50, 3664–3673. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect. Genet. Evol. 2016, 39, 22–34. [Google Scholar] [CrossRef]
- Ramey, A.M.; Reeves, A.B.; Ogawa, H.; Ip, H.S.; Imai, K.; Bui, V.N.; Yamaguchi, E.; Silko, N.Y.; Afonso, C.L. Genetic diversity and mutation of avian paramyxovirus serotype 1 (Newcastle disease virus) in wild birds and evidence for intercontinental spread. Arch. Virol. 2013, 158, 2495–2503. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Chang, C.Y.; Lee, F.; Chiou, C.J.; Tsai, H.J. Phylogenetic analysis of avian paramyxoviruses 1 isolated in Taiwan from 2010 to 2018 and evidence for their intercontinental dispersal by migratory birds. J. Vet. Med. Sci. 2020, 82, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- Diel, D.G.; da Silva, L.H.; Liu, H.; Wang, Z.; Miller, P.J.; Afonso, C.L. Genetic diversity of avian paramyxovirus type 1: Proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect. Genet. Evol. 2012, 12, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Nooruzzaman, M.; Mazumder, A.C.; Khatun, S.; Chowdhury, E.H.; Das, P.M.; Islam, M.R. Pathotypic and genotypic characterization of two Bangladeshi isolates of Newcastle disease virus of chicken and pigeon origin. Transbound. Emerg. Dis. 2015, 62, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Deka, P.; Nath, M.K.; Das, S.; Das, B.C.; Phukan, A.; Lahkar, D.; Bora, B.; Shokeen, K.; Kumar, A.; Deka, P. A study of risk factors associated with Newcastle disease and molecular characterization of genotype XIII Newcastle disease virus in backyard and commercial poultry in Assam, India. Res. Vet. Sci. 2022, 150, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, Z.; Tabatabaeizadeh, S.E.; Toroghi, R.; Farzin, H.; Saffarian, P. First detection and characterisation of sub-genotype XIII.2.1 Newcastle disease virus isolated from backyard chickens in Iran. Vet. Med. Sci. 2022, 8, 2521–2531. [Google Scholar] [CrossRef]
- da Silva, A.P.; Aston, E.J.; Chiwanga, G.H.; Birakos, A.; Muhairwa, A.P.; Kayang, B.B.; Kelly, T.; Zhou, H.; Gallardo, R.A. Molecular characterization of Newcastle disease viruses isolated from chickens in Tanzania and Ghana. Viruses 2020, 12, 916. [Google Scholar] [CrossRef]
- Forrester, N.L.; Widen, S.G.; Wood, T.G.; Travassos da Rosa, A.P.; Ksiazek, T.G.; Vasilakis, N.; Tesh, R.B. Identification of a new Newcastle disease virus isolate from Indonesia represents an ancestral lineage of class II genotype XIII. Virus Genes 2013, 47, 168–172. [Google Scholar] [CrossRef]
- Miller, P.J.; Haddas, R.; Simanov, L.; Lublin, A.; Rehmani, S.F.; Wajid, A.; Bibi, T.; Khan, T.A.; Yaqub, T.; Setiyaningsih, S.; et al. Identification of new sub-genotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Genet. Evol. 2015, 29, 216–229. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Wajid, A.; Basharat, A.; Bibi, T.; Rehmani, S.F. Comparison of protection and viral shedding following vaccination with Newcastle disease virus strains of different genotypes used in vaccine formulation. Trop. Anim. Health Prod. 2018, 50, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Kwon, H.J.; Kim, T.E.; Kim, J.H.; Yoo, H.S.; Park, M.H.; Park, Y.H.; Kim, S.J. Characterization of a recombinant Newcastle disease virus vaccine strain. Clin. Vaccine Immunol. 2008, 15, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Ma, H.; Wu, Y.; Liu, W.; Wang, X.; Liu, Y.; Liu, X. A vaccine candidate of attenuated genotype VII Newcastle disease virus generated by reverse genetics. Vaccine 2009, 27, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Mahamud, S.N.A.; Bello, M.B.; Ideris, A.; Omar, A.R. Efficacy of genotype-matched Newcastle disease virus vaccine formulated in carboxymethyl sago starch acid hydrogel in chickens vaccinated via different routes. J. Vet. Sci. 2022, 23, e25. [Google Scholar] [CrossRef]
- Izquierdo-Lara, R.; Chumbe, A.; Calderón, K.; Fernández-Díaz, M.; Vakharia, V.N. Genotype-matched Newcastle disease virus vaccine confers improved protection against genotype XII challenge: The importance of cytoplasmic tails in viral replication and vaccine design. PLoS ONE 2019, 14, e0209539. [Google Scholar] [CrossRef]
- Sultan, H.A.; Talaat, S.; Elfeil, W.K.; Selim, K.; Kutkat, M.A.; Amer, S.A.; Choi, K.S. Protective efficacy of the Newcastle disease virus genotype VII-matched vaccine in commercial layers. Poultry Sci. 2020, 99, 1275–1286. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef]
- Barman, L.R.; Flensburg, M.F.; Permin, A.; Madsen, M.; Islam, M.R. A controlled study to assess the effects of vaccination against Newcastle disease in village chickens. Bangladesh Vet. 2010, 27, 56–61. [Google Scholar] [CrossRef]
- Biswas, H.R.; Haoque, M.M.; Oxley, M.; Prodhan, M.A.M. A comparative study on the protection of indigenous chickens against Newcastle disease induced by Australian NDV4HR and locally produced conventional vaccines in Bangladesh. Prev. Vet. Med. 1996, 26, 157–164. [Google Scholar] [CrossRef]
- Rahman, M.M.; Sarker, R.D.; Nooruzzaman, M. Evaluation of serum antibody titer level against Newcastle disease virus in vaccinated broiler chickens. Ann. Vet. Anim. Sci. 2017, 4, 94–98. [Google Scholar]
- Parvin, R.; Kabiraj, C.K.; Hossain, I.; Hassan, A.; Begum, J.A.; Nooruzzaman, M.; Islam, M.T.; Chowdhury, E.H. Investigation of respiratory disease outbreaks of poultry in Bangladesh using two real-time PCR-based simultaneous detection assays. Front. Vet. Sci. 2022, 9, 1036757. [Google Scholar] [CrossRef] [PubMed]
- Kabiraj, C.K.; Mumu, T.T.; Chowdhury, E.H.; Islam, M.R.; Nooruzzaman, M. Sequential pathology of a genotype XIII Newcastle disease virus from Bangladesh in chickens on experimental infection. Pathogens 2020, 9, 539. [Google Scholar] [CrossRef]
- Yusoff, K.; Nesbit, M.; McCartney, H.; Meulemans, G.; Alexander, D.J.; Collins, M.S.; Emmerson, P.T.; Samson, A.C. Location of neutralizing epitopes on the fusion protein of Newcastle disease virus strain Beaudette C. J. Gen. Virol. 1989, 70, 3105–3109. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef]
- Cornax, I.; Miller, P.J.; Afonso, C.L. Characterization of live LaSota vaccine strain-induced protection in chickens upon early challenge with a virulent Newcastle disease virus of heterologous genotype. Avian Dis. 2012, 56, 464–470. [Google Scholar] [CrossRef]
- van Eck, J.H.; van Wiltenburg, N.; Jaspers, D. An Ulster 2C strain-derived Newcastle disease vaccine: Efficacy and excretion in maternally immune chickens. Avian Pathol. 1991, 20, 481–495. [Google Scholar] [CrossRef]
- Miller, P.J.; Afonso, C.L.; El Attrache, J.; Dorsey, K.M.; Courtney, S.C.; Guo, Z.; Kapczynski, D.R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev. Comp. Immunol. 2013, 41, 505–513. [Google Scholar] [CrossRef]
- Lancaster, J.E. Newcastle Disease: A Review 1926–1964. Avian Dis. 1968, 12, 80–115. [Google Scholar]
- Folitse, R.; Halvorson, D.A.; Sivanandan, V. Efficacy of combined killed-in-oil emulsion and live Newcastle disease vaccines in chickens. Avian Dis. 1998, 42, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Chansiripornchai, N.; Sasipreeyajan, J. Efficacy of live B1 or Ulster 2C Newcastle disease vaccines simultaneously vaccinated with inactivated oil adjuvant vaccine for protection of Newcastle disease virus in broiler chickens. Acta Vet. Scand. 2006, 48, 2. [Google Scholar] [CrossRef] [PubMed]
- Winterfield, R.W.; Dhillon, A.S.; Alby, L.J. Vaccination of chickens against Newcastle disease with live and inactivated Newcastle disease virus. Poultry Sci. 1980, 59, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Hamed, M.; Osman, N. Evaluation of protection efficiency of different vaccination programs against velogenic Newcastle disease virus in broiler chickens: Comparative in field and in laboratory studies. Thai Vet. Med. 2021, 51, 141–150. [Google Scholar] [CrossRef]
- Yang, H.M.; Zhao, J.; Xue, J.; Yang, Y.L.; Zhang, G.Z. Antigenic variation of LaSota and genotype VII Newcastle disease virus (NDV) and their efficacy against challenge with velogenic NDV. Vaccine 2017, 35, 27–32. [Google Scholar] [CrossRef]
- Cardenas-Garcia, S.; Diel, D.G.; Susta, L.; Lucio-Decanini, E.; Yu, Q.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Development of an improved vaccine evaluation protocol to compare the efficacy of Newcastle disease vaccines. Biologicals 2015, 43, 136–145. [Google Scholar] [CrossRef]
- Panshin, A.; Shihmanter, E.; Weisman, Y.; Orvell, C.; Lipkind, M. Antigenic heterogeneity amongst the field isolates of Newcastle disease virus (NDV) in relation to the vaccine strain. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 173–185. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Booster Vaccination | Challenge | Respiratory Distress | Ocular Discharge | Nasal Discharge | Diarrhea | Nervous Sign |
|---|---|---|---|---|---|---|---|
| A1 | LaSota (Live) | Yes | 0/10 * | 0/10 | 0/10 | 3/10 | 0/10 |
| A2 | LaSota (Killed) | Yes | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| A3 | BD-C161/2010 (Killed) | Yes | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| B1 | Unvaccinated | Yes | 10/10 | 4/10 | 2/10 | 7/10 | 3/10 |
| B2 | Unvaccinated | No | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| 5 dpc (Group A1, A2, A3)/3-5 dpc (Group B1) | 7 dpc | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Booster Vaccine | Oropharyngeal Swab | Cloacal Swab | Oropharyngeal Swab | Cloacal Swab | ||||
| No. | Ct | No. | Ct | No. | Ct | No. | Ct | ||
| A1 | LaSota (Live) | 3/10 | 31.5–33.2 | 5/10 | 26.2–32.4 | 5/10 | 22.3–30.1 | 3/10 | 19.6–23.0 |
| A2 | LaSota (Killed) | 2/10 | 31.4–31.7 | 0/10 | - | 1/10 | 31.8 | 0/10 | - |
| A3 | BD-C161/2010 (Killed) | 1/10 | 32.6 | 0/10 | - | 0/10 | - | 0/10 | - |
| B1 | Unvaccinated | 10/10 | 16.7–25.1 | 10/10 | 18.1–25.0 | N/A | - | N/A | - |
| Epitopes | A1 | A2 | A3 | A4 | A5 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Position | 72 | 78 | 79 | 157 | 158 | 159 | 160 | 161 | 162 | 163 | 164 | 165 | 166 | 167 | 168 | 169 | 170 | 171 | 343 |
| LaSota | D | K | A | S | I | A | A | T | N | E | A | V | H | E | V | T | D | G | L |
| BD-C161/2010 | • | • | T | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, I.; Subarna, J.F.; Kabiraj, C.K.; Begum, J.A.; Parvin, R.; Martins, M.; Diel, D.G.; Chowdhury, E.H.; Islam, M.R.; Nooruzzaman, M. A Booster with a Genotype-Matched Inactivated Newcastle Disease Virus (NDV) Vaccine Candidate Provides Better Protection against a Virulent Genotype XIII.2 Virus. Vaccines 2023, 11, 1005. https://doi.org/10.3390/vaccines11051005
Hossain I, Subarna JF, Kabiraj CK, Begum JA, Parvin R, Martins M, Diel DG, Chowdhury EH, Islam MR, Nooruzzaman M. A Booster with a Genotype-Matched Inactivated Newcastle Disease Virus (NDV) Vaccine Candidate Provides Better Protection against a Virulent Genotype XIII.2 Virus. Vaccines. 2023; 11(5):1005. https://doi.org/10.3390/vaccines11051005
Chicago/Turabian StyleHossain, Ismail, Jannatul Ferdous Subarna, Congriev Kumar Kabiraj, Jahan Ara Begum, Rokshana Parvin, Mathias Martins, Diego G. Diel, Emdadul Haque Chowdhury, Mohammad Rafiqul Islam, and Mohammed Nooruzzaman. 2023. "A Booster with a Genotype-Matched Inactivated Newcastle Disease Virus (NDV) Vaccine Candidate Provides Better Protection against a Virulent Genotype XIII.2 Virus" Vaccines 11, no. 5: 1005. https://doi.org/10.3390/vaccines11051005