
PP19128R, a Multiepitope Vaccine Designed to Prevent Latent Tuberculosis Infection, Induced Immune Responses In Silico and In Vitro Assays



Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Analysis
2.1.1. Selection of Candidate Antigens for MTB
2.1.2. Screening for Immunodominant T Cell Epitopes
2.1.3. Immunodominant B-Cell Epitope Prediction
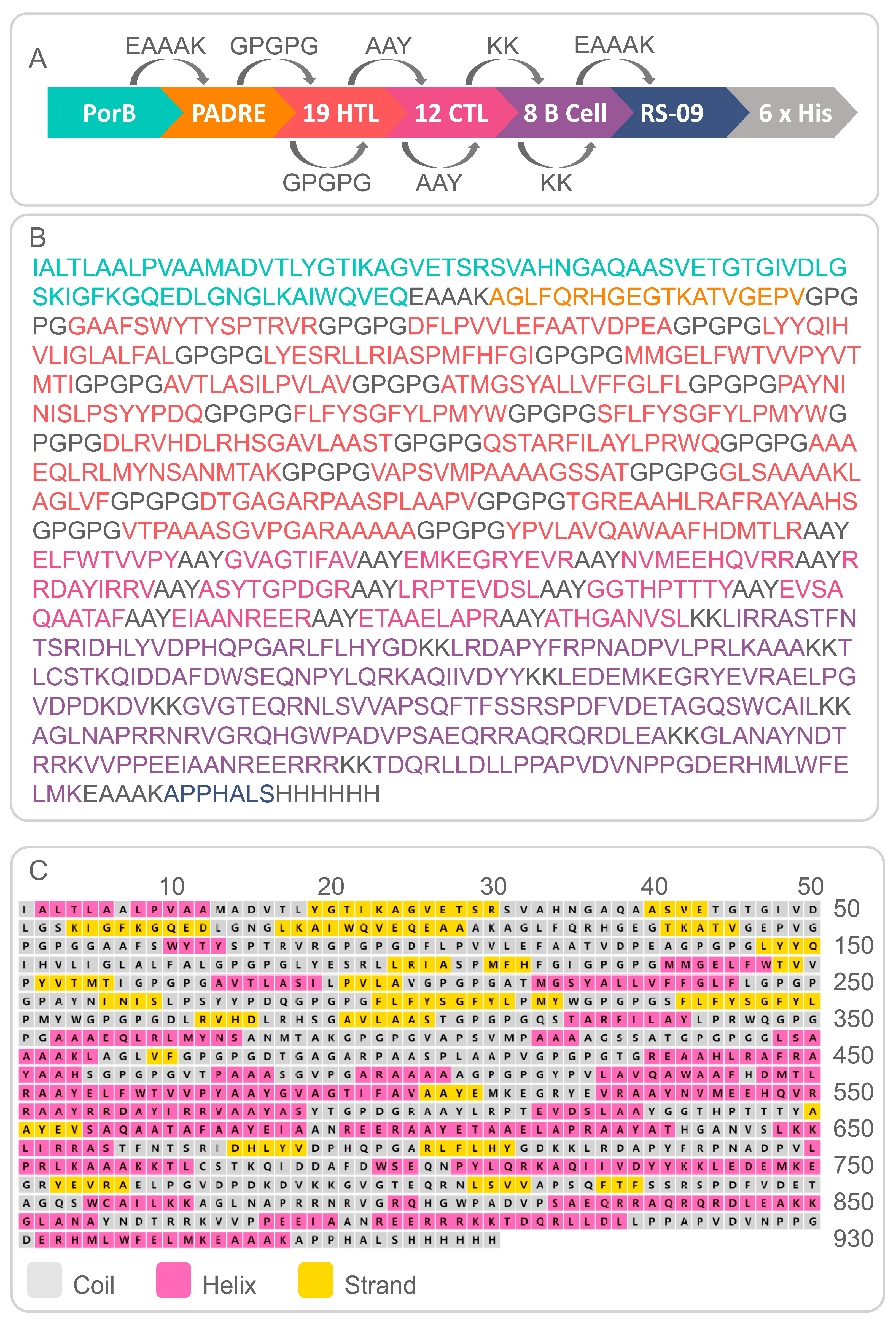
2.1.4. Analysis of Population Coverage of HLA-I and HLA-II Alleles and Construction of MEV
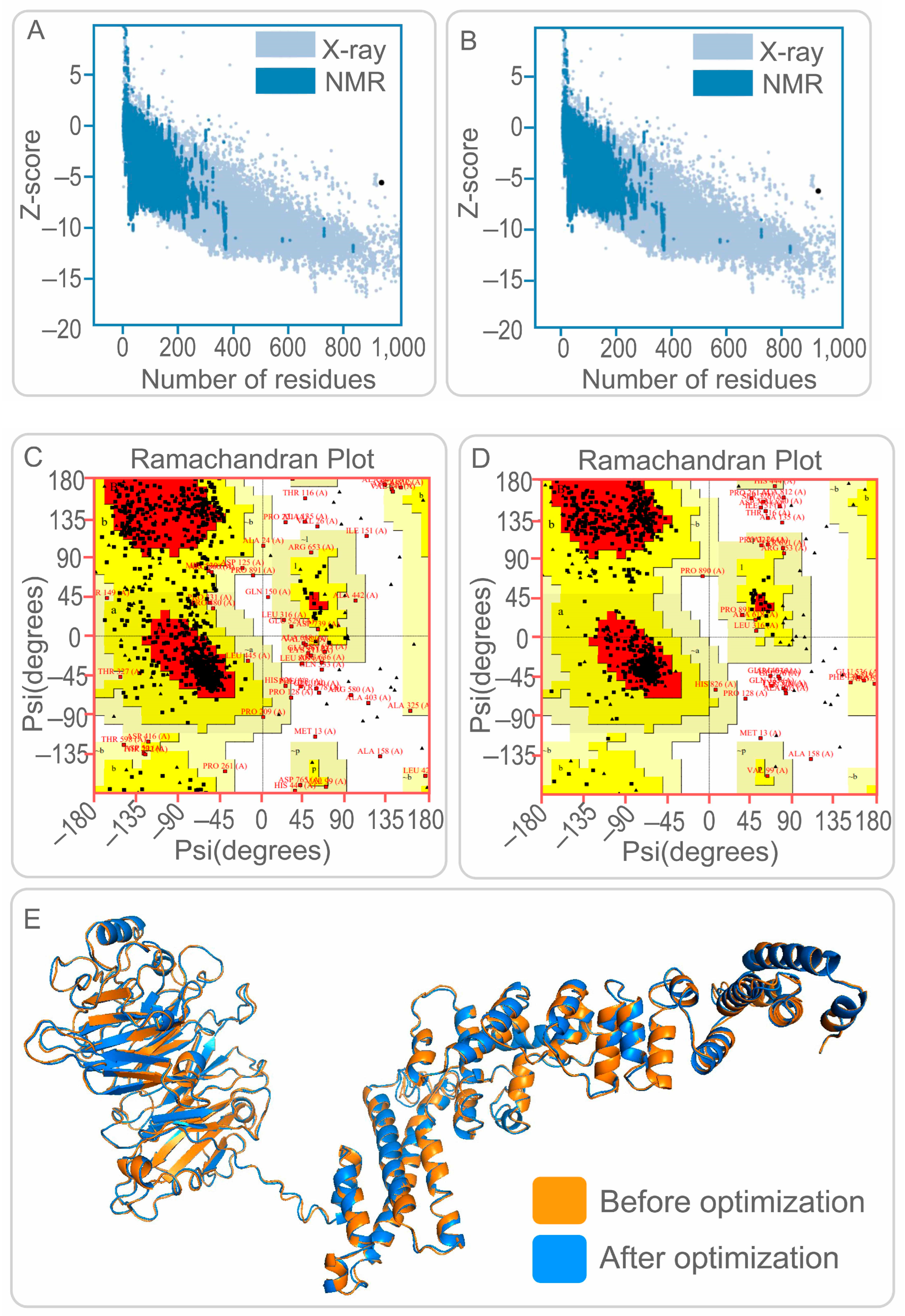
2.1.5. Prediction of Physicochemical Properties and Secondary/Tertiary Structure of the PP19128 Vaccine
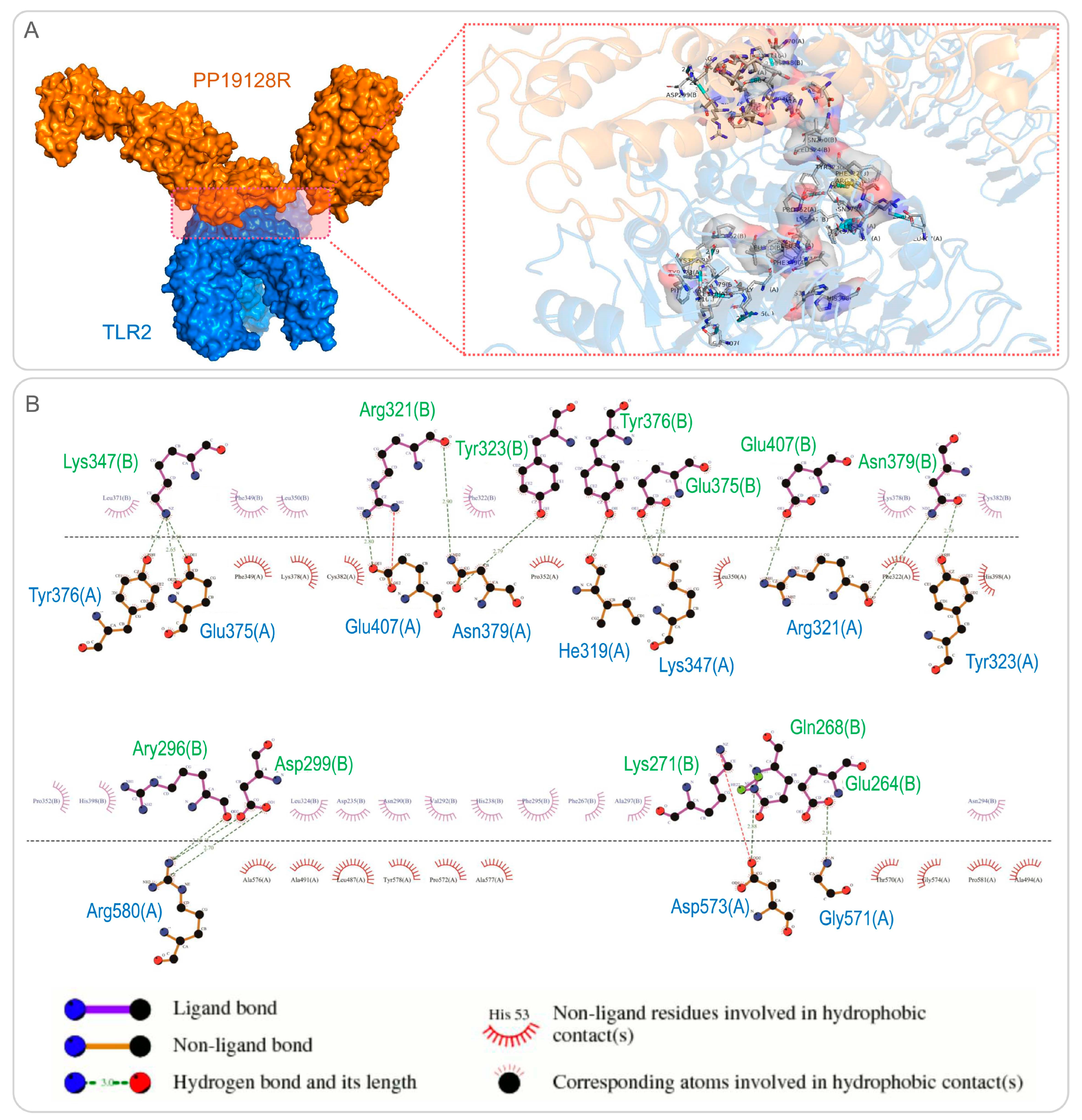
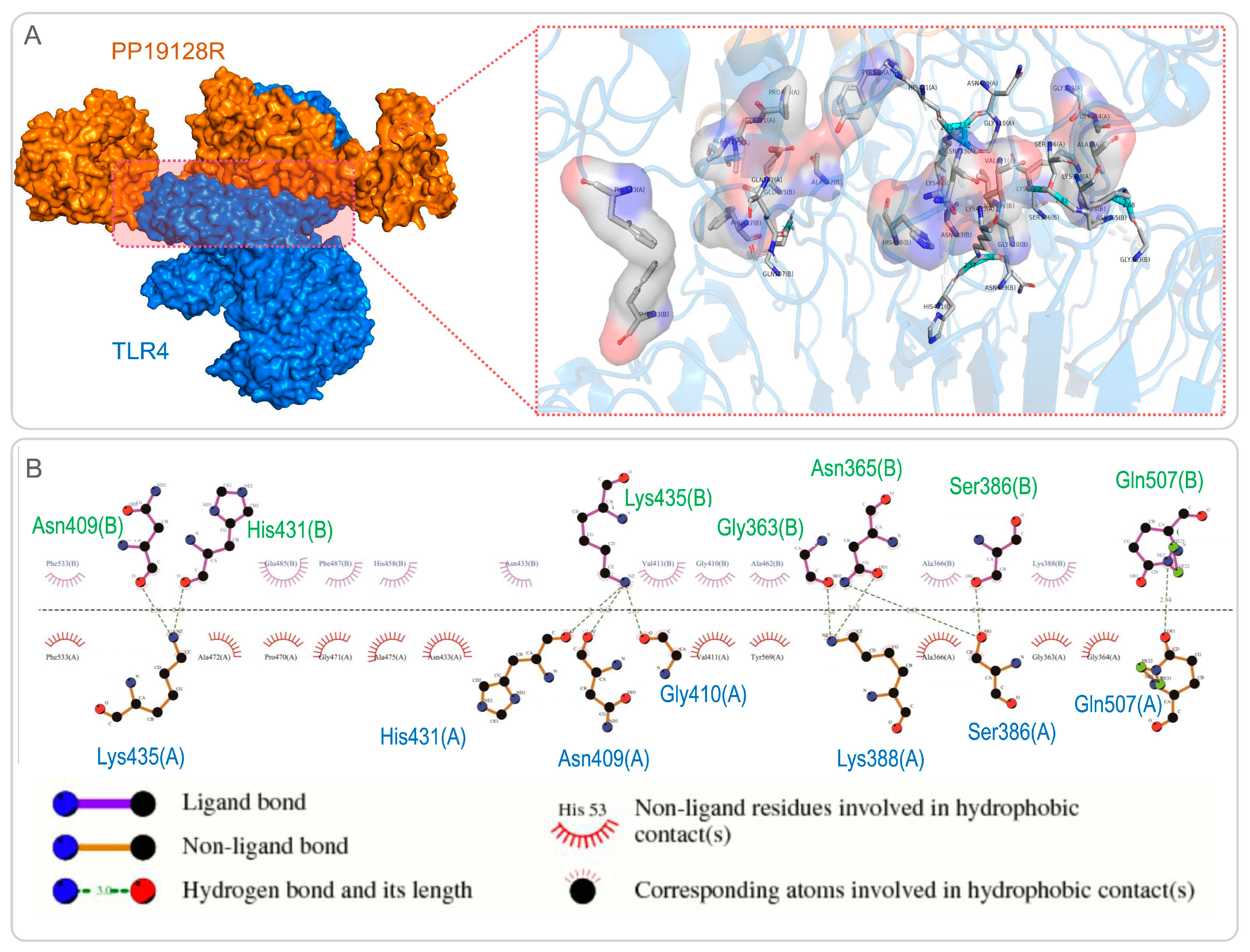
2.1.6. Molecular Docking between the PP19128 Vaccine and TLRs and Their Simulated Immune Responses
2.2. In Vitro Experiments
2.2.1. Ethics and Experimental Subjects
2.2.2. Construction of Recombinant Plasmid and Expression of the PP19128R Vaccine In Vitro
2.2.3. ELISpot
2.2.4. Cytometric Bead Assay (CBA)
2.2.5. Data Collation and Statistical Analysis
3. Results
3.1. Identification of the Immunodominant Epitopes of HTL, CTL, and B-Cells and Analysis of the Population Coverage
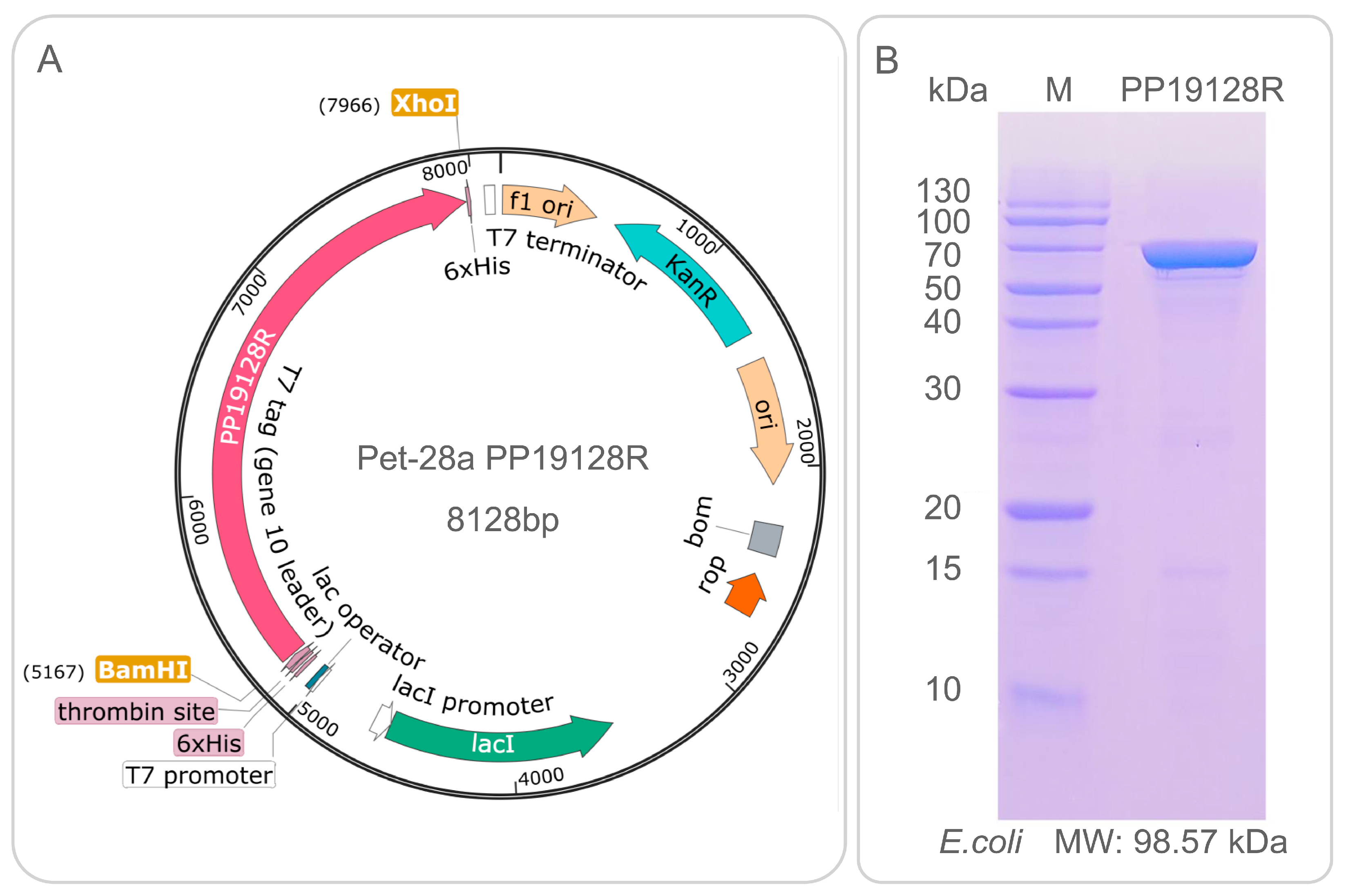
3.2. Construction of PP19128R Vaccine and Prediction of Its Physicochemical Properties and Secondary/Tertiary Structure
3.3. The PP19128R Vaccine Binds Tightly to TLR2 and TLR4
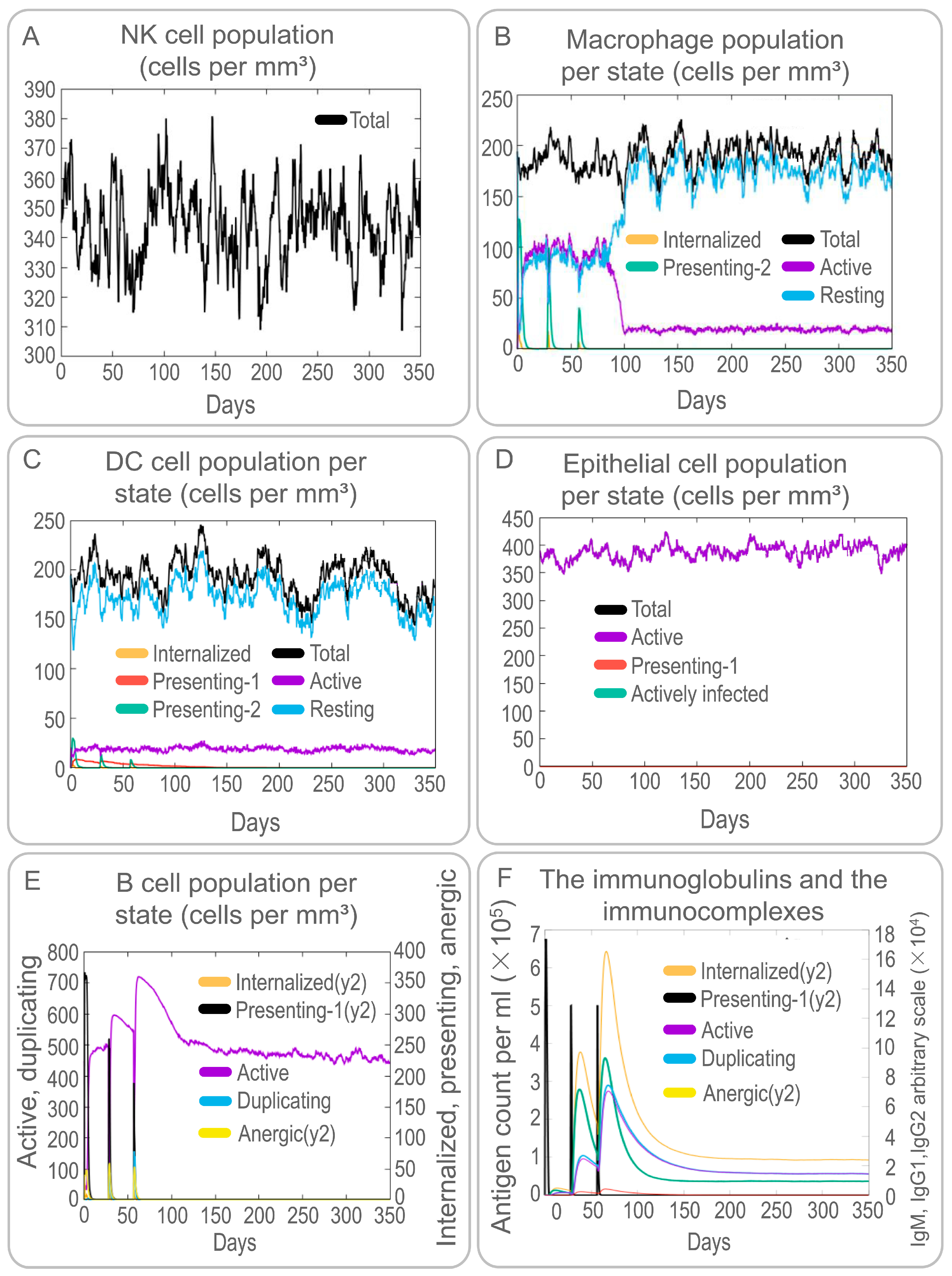
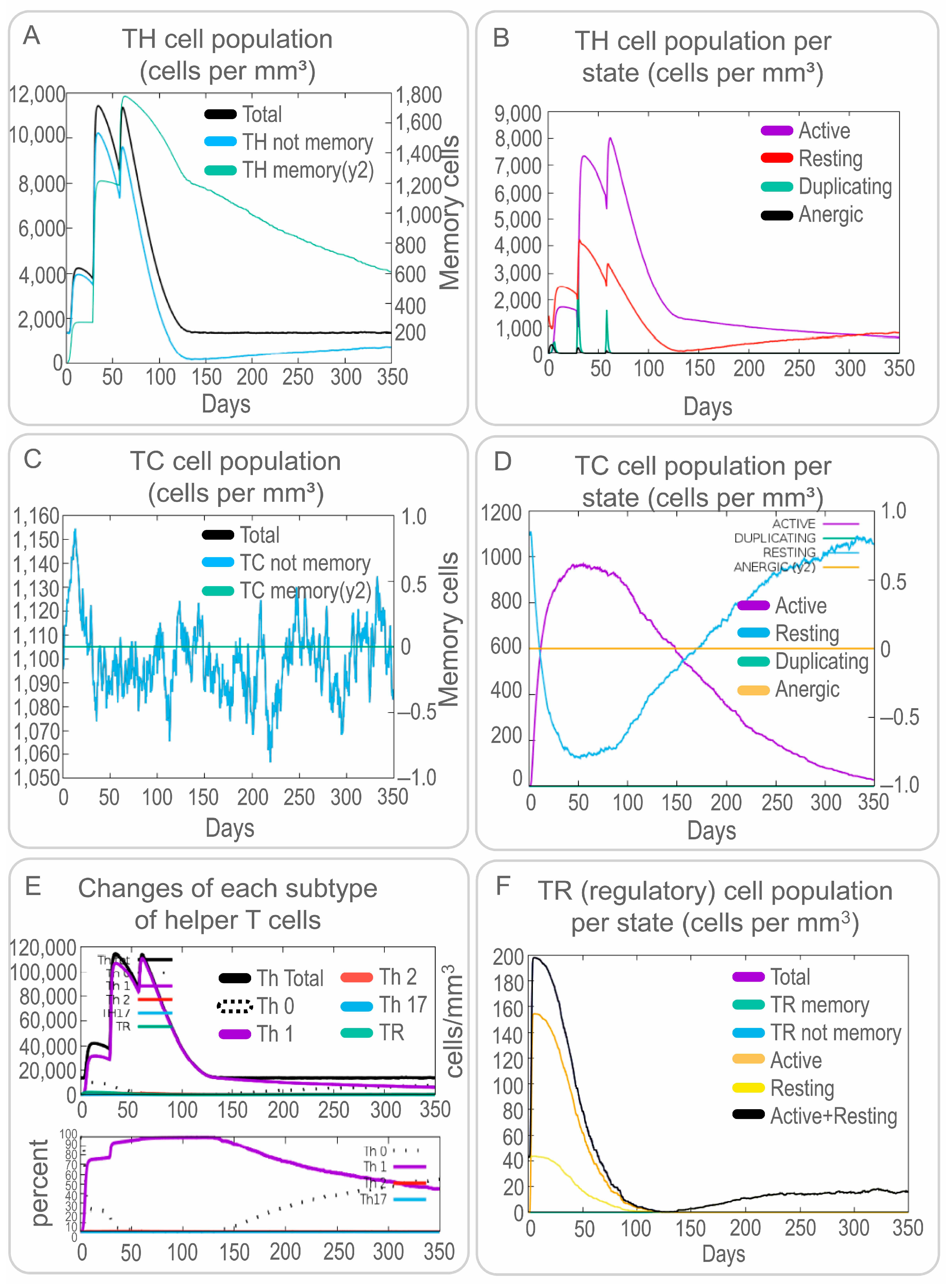
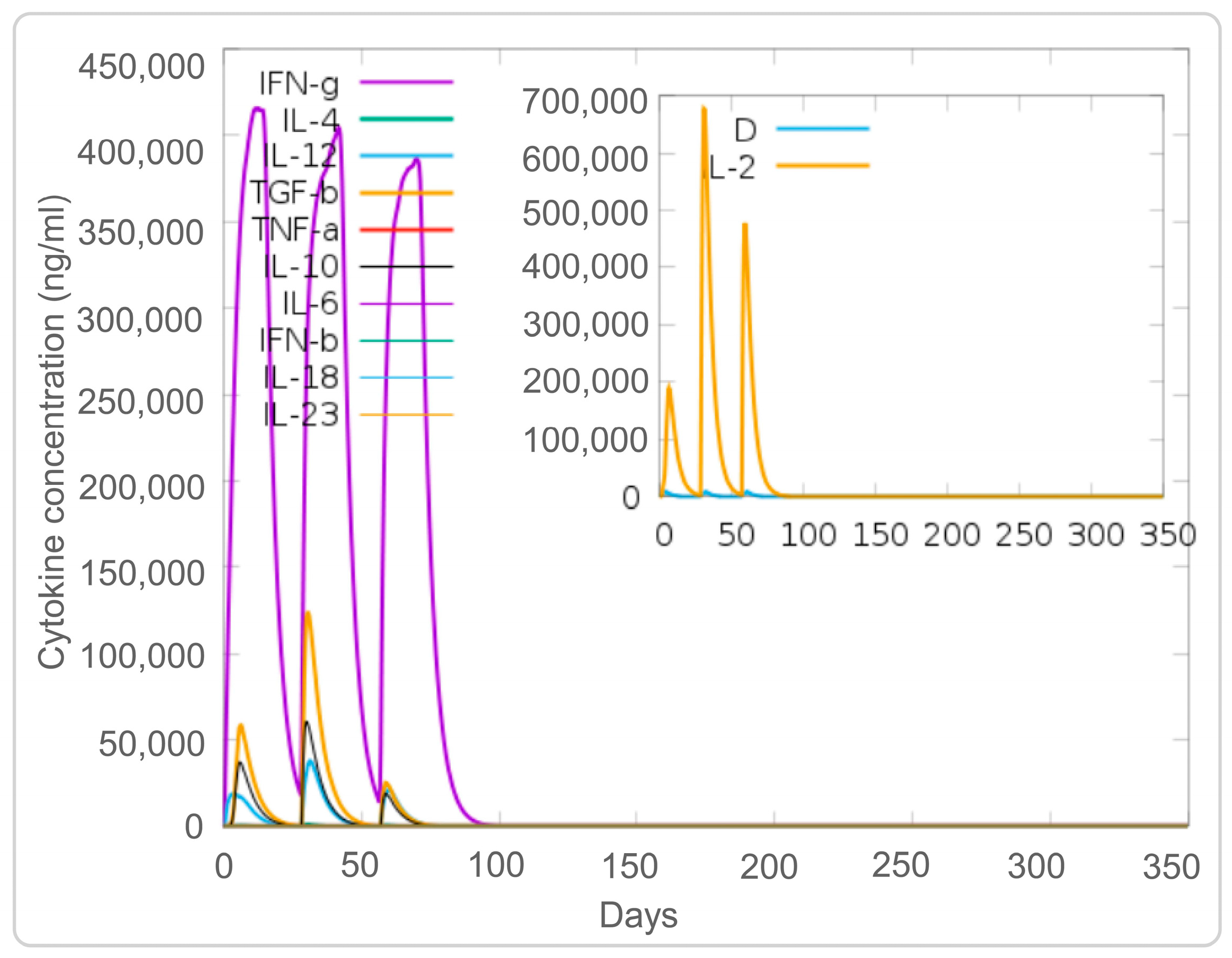
3.4. The PP19128R Vaccine Induced a Robust Immune Response In Silico
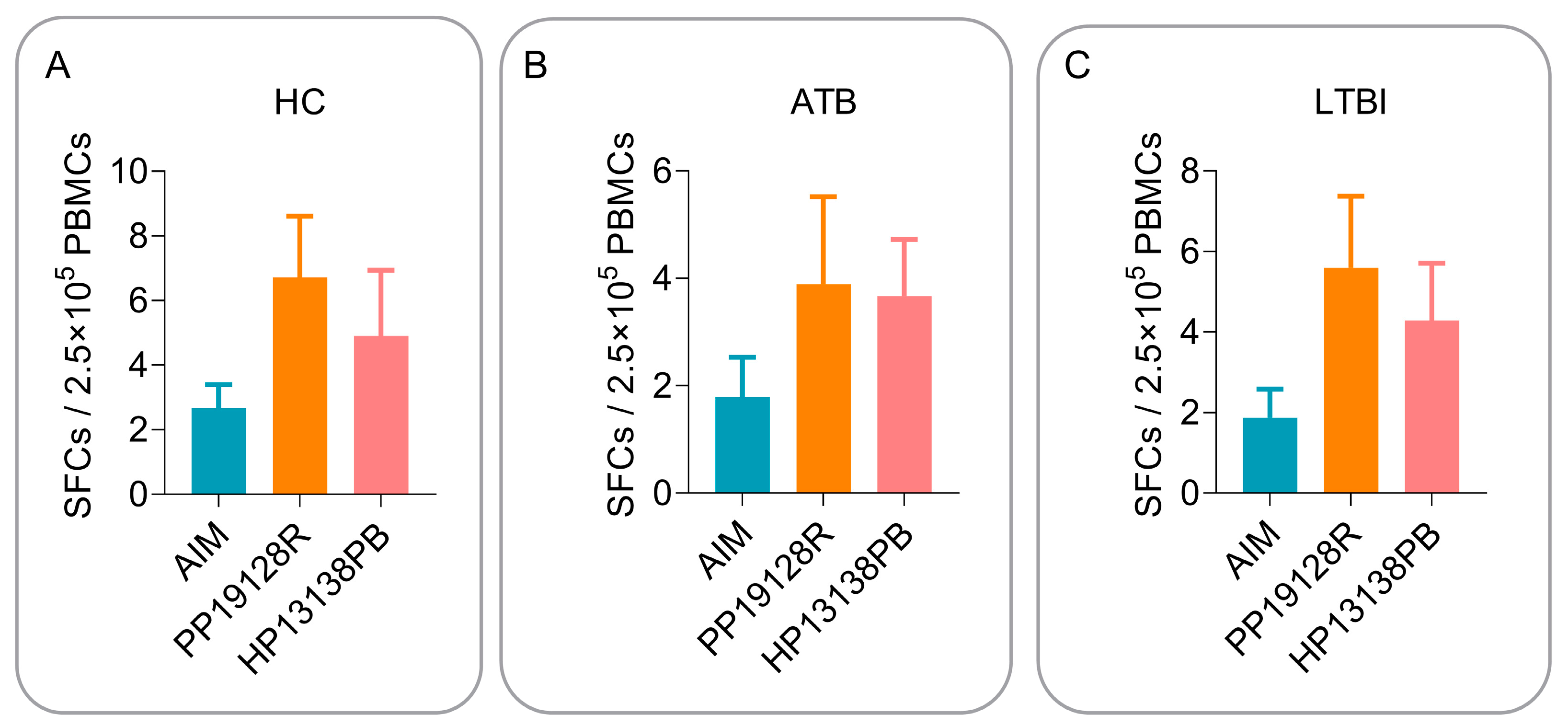
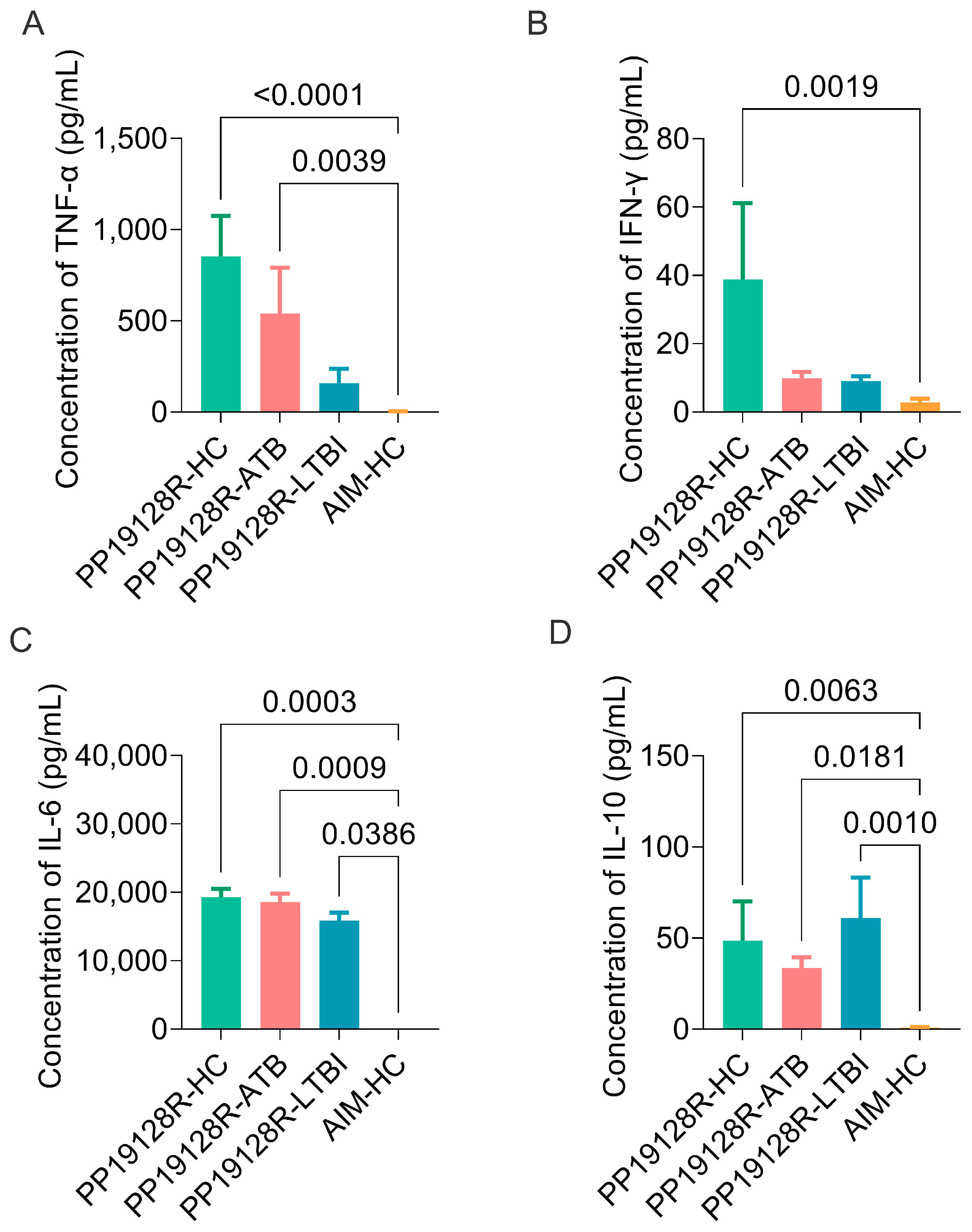
3.5. The PP19128R Vaccine Induced Significantly Higher Levels of Immune Responses In Vitro
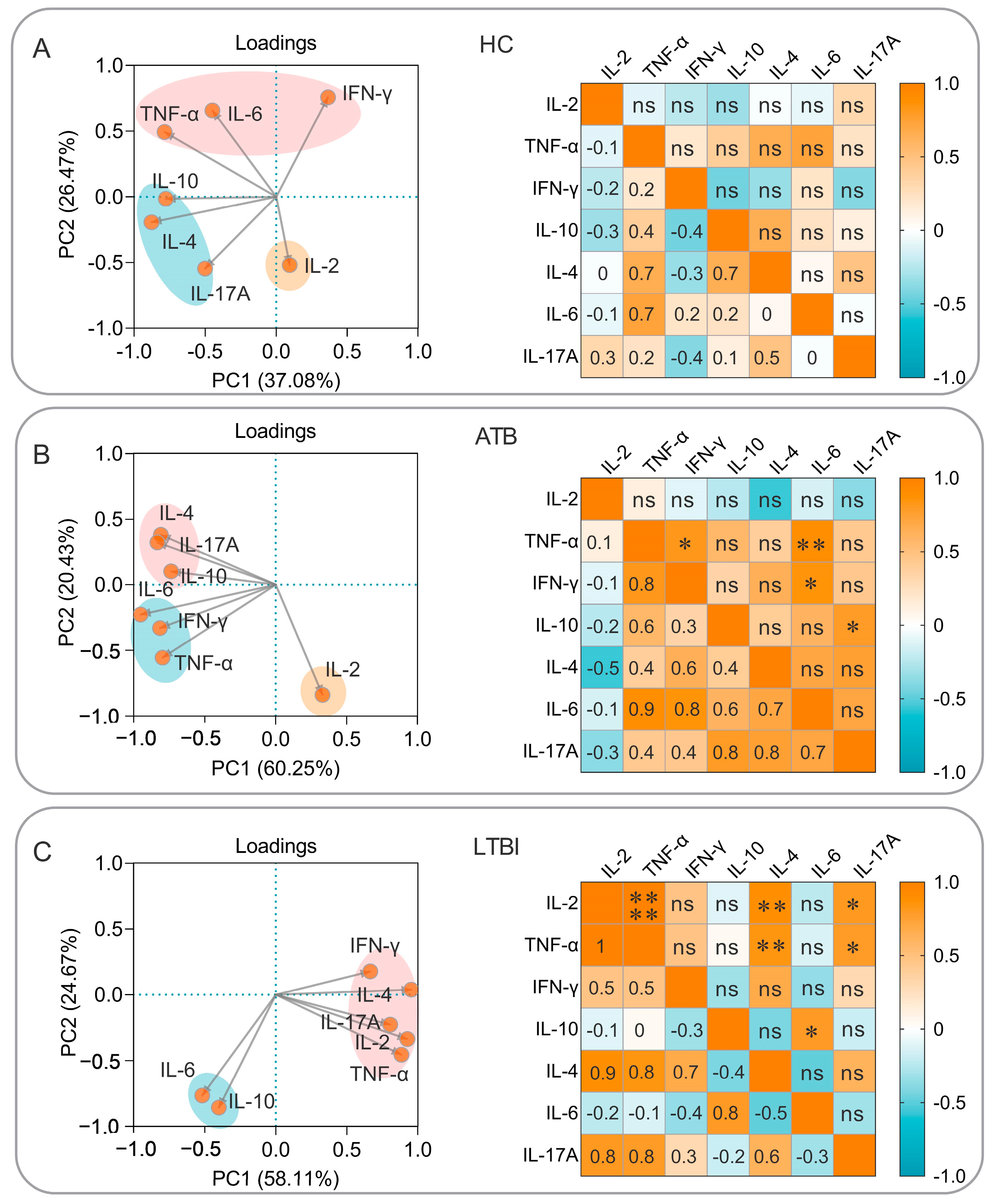
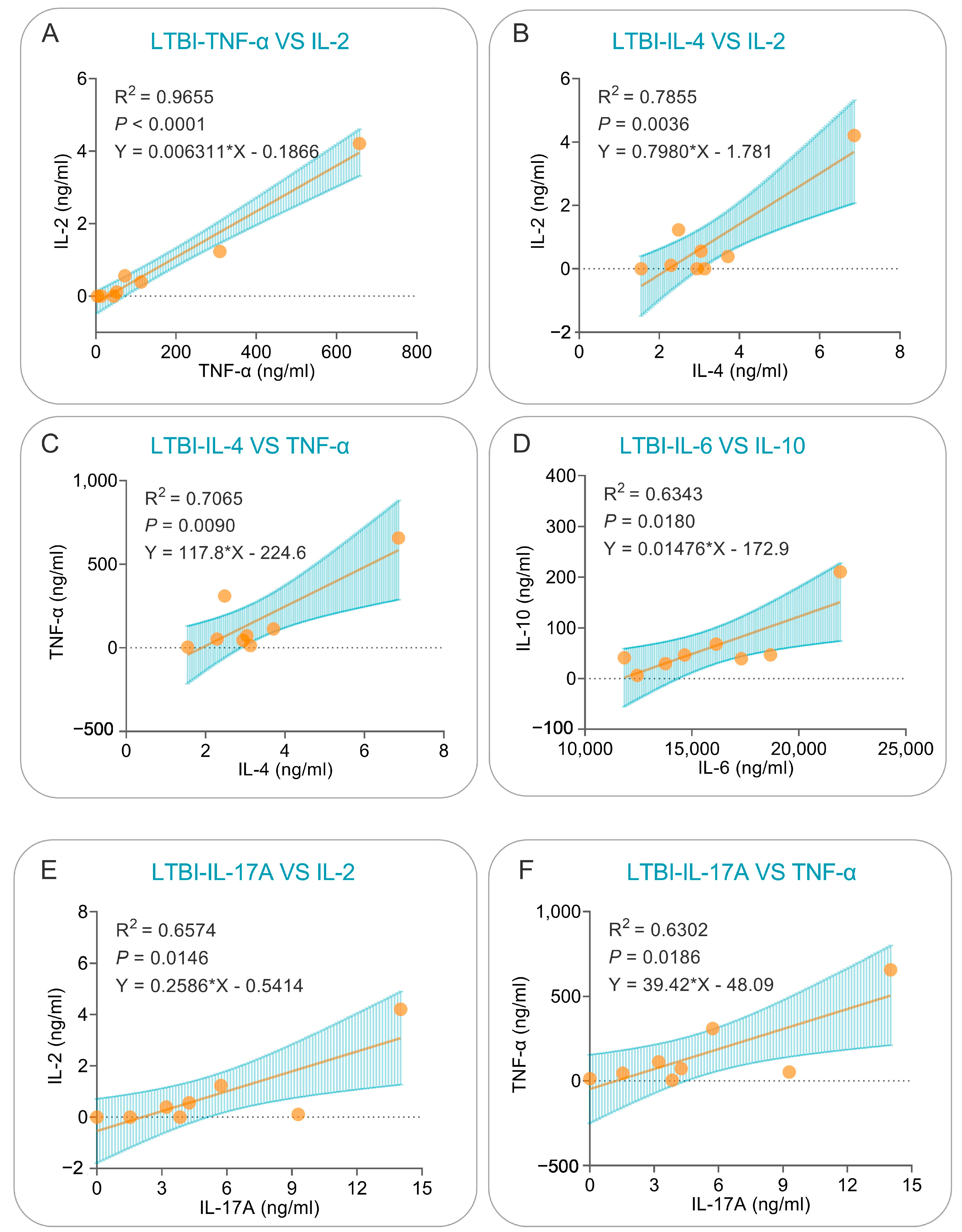
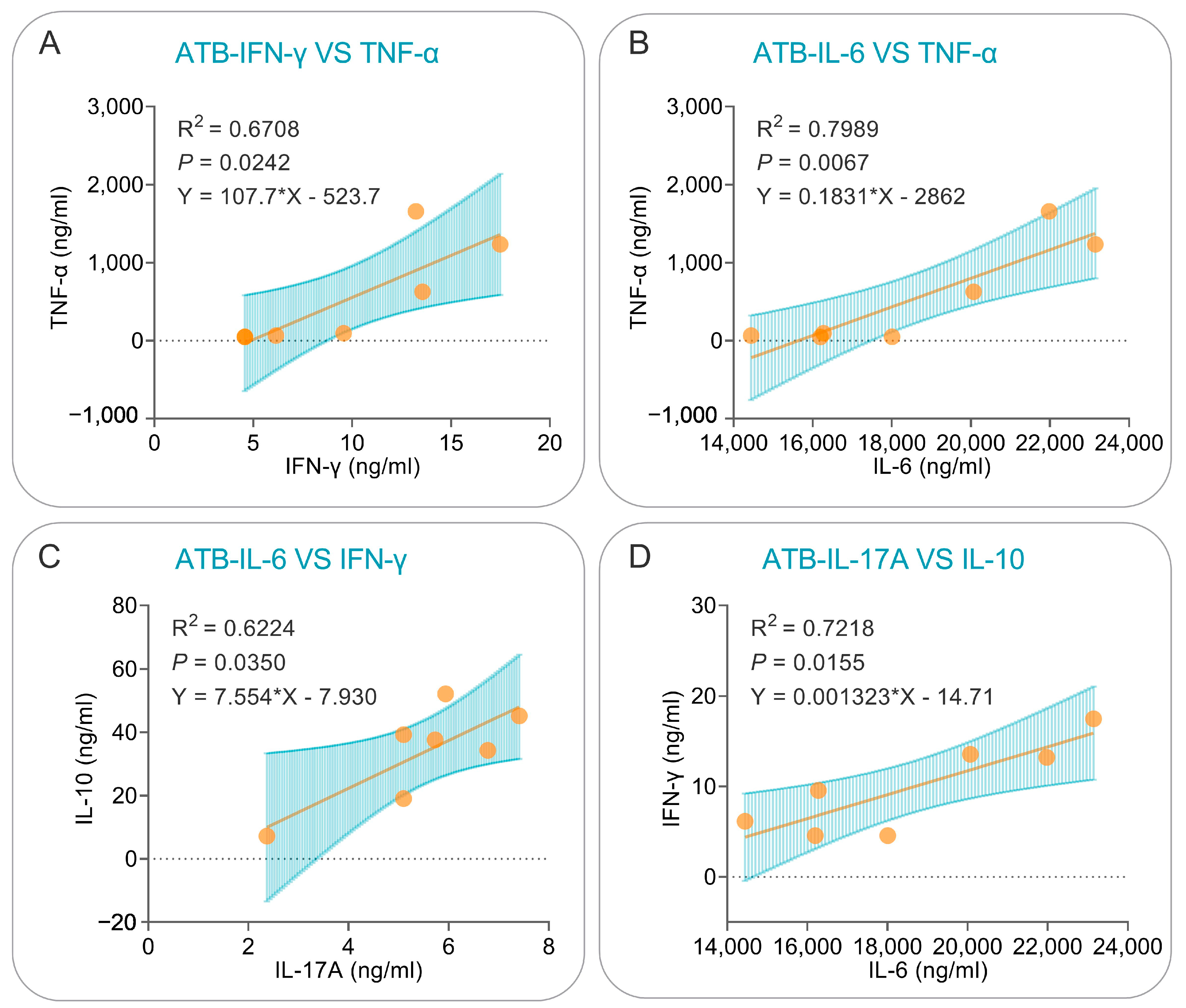
3.6. Correlation Analysis and Simple Linear Regression Analysis of Cytokines Induced by the PP19128R Vaccine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
References
- Zhai, W.; Wu, F.; Zhang, Y.; Fu, Y.; Liu, Z. The immune escape mechanisms of Mycobacterium tuberculosis. Int. J. Mol. Sci. 2019, 20, 340. [Google Scholar] [CrossRef] [PubMed]
- Angelidou, A.; Diray-Arce, J.; Conti, M.G.; Smolen, K.K.; van Haren, S.D.; Dowling, D.J.; Husson, R.N.; Levy, O. BCG as a case study for precision vaccine development: Lessons from vaccine heterogeneity, trained immunity, and immune ontogeny. Front. Microbiol. 2020, 11, 332. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Tuberculosis Report 2022; World Health Organization: Geneva, Switzerland, 2022.
- Gong, W.; Pan, C.; Cheng, P.; Wang, J.; Zhao, G.; Wu, X. Peptide-based vaccines for tuberculosis. Front Immunol. 2022, 13, 830497. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; An, H.; Wang, J.; Cheng, P.; Qi, Y. The natural effect of BCG vaccination on COVID-19: The debate continues. Front. Immunol. 2022, 13, 953228. [Google Scholar] [CrossRef] [PubMed]
- Mangtani, P.; Abubakar, I.; Ariti, C.; Beynon, R.; Pimpin, L.; Fine, P.E.; Rodrigues, L.C.; Smith, P.G.; Lipman, M.; Whiting, P.F.; et al. Protection by BCG vaccine against tuberculosis: A systematic review of randomized controlled trials. Clin. Infect. Dis. 2014, 58, 470–480. [Google Scholar] [CrossRef]
- Andersen, P.; Doherty, T.M. The success and failure of BCG—Implications for a novel tuberculosis vaccine. Nat. Rev. Microbiol. 2005, 3, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Tang, F.; Wang, J.; Cheng, P.; Wang, L.; Gong, W. Immunoinformatic-based multi-epitope vaccine design for co-Infection of Mycobacterium tuberculosis and SARS-CoV-2. J. Pers. Med. 2023, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.; Hawkridge, T.; Hanekom, W. Childhood tuberculosis: Old and new vaccines. Paediatr. Respir. Rev. 2007, 8, 148–154. [Google Scholar] [CrossRef]
- WHO. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020.
- Schrager, L.K.; Vekemens, J.; Drager, N.; Lewinsohn, D.M.; Olesen, O.F. The status of tuberculosis vaccine development. Lancet Infect. Dis. 2020, 20, e28–e37. [Google Scholar] [CrossRef]
- Tait, D.R.; Hatherill, M.; Van Der Meeren, O.; Ginsberg, A.M.; Van Brakel, E.; Salaun, B.; Scriba, T.J.; Akite, E.J.; Ayles, H.M.; Bollaerts, A.; et al. Final analysis of a trial of M72/AS01(E) vaccine to prevent tuberculosis. N. Engl. J. Med. 2019, 381, 2429–2439. [Google Scholar] [CrossRef]
- Lewinsohn, D.M.; Leonard, M.K.; LoBue, P.A.; Cohn, D.L.; Daley, C.L.; Desmond, E.; Keane, J.; Lewinsohn, D.A.; Loeffler, A.M.; Mazurek, G.H.; et al. Official american thoracic society/infectious diseases society of america/centers for disease control and prevention clinical practice guidelines: Diagnosis of tuberculosis in adults and children. Clin. Infect. Dis. 2017, 64, e1–e33. [Google Scholar] [CrossRef]
- Behr, M.A.; Kaufmann, E.; Duffin, J.; Edelstein, P.H.; Ramakrishnan, L. Latent tuberculosis: Two centuries of confusion. Am. J. Respir. Crit. Care Med. 2021, 204, 142–148. [Google Scholar] [CrossRef]
- Houben, R.M.; Dodd, P.J. The global burden of latent tuberculosis infection: A re-estimation using mathematical modelling. PLoS Med. 2016, 13, e1002152. [Google Scholar] [CrossRef]
- De Groot, A.S.; Moise, L.; Terry, F.; Gutierrez, A.H.; Hindocha, P.; Richard, G.; Hoft, D.F.; Ross, T.M.; Noe, A.R.; Takahashi, Y.; et al. Better epitope discovery, precision immune engineering, and accelerated vaccine design using immunoinformatics tools. Front. Immunol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed]
- Oli, A.N.; Obialor, W.O.; Ifeanyichukwu, M.O.; Odimegwu, D.C.; Okoyeh, J.N.; Emechebe, G.O.; Adejumo, S.A.; Ibeanu, G.C. Immunoinformatics and vaccine development: An overview. Immunotargets Ther. 2020, 9, 13–30. [Google Scholar] [CrossRef]
- Urrutia-Baca, V.H.; Gomez-Flores, R.; De La Garza-Ramos, M.A.; Tamez-Guerra, P.; Lucio-Sauceda, D.G.; Rodríguez-Padilla, M.C. Immunoinformatics approach to design a novel epitope-based oral vaccine against Helicobacter pylori. J. Comput. Biol. 2019, 26, 1177–1190. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Mo, Y.; Wang, Y.; Wu, P.; Zhang, Y.; Xiong, F.; Guo, C.; Wu, X.; Li, Y.; Li, X.; et al. Neoantigen vaccine: An emerging tumor immunotherapy. Mol. Cancer 2019, 18, 128. [Google Scholar] [CrossRef] [PubMed]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Skwarczynski, M.; Chandrudu, S.; Rigau-Planella, B.; Islam, M.T.; Cheong, Y.S.; Liu, G.; Wang, X.; Toth, I.; Hussein, W.M. Progress in the development of subunit vaccines against malaria. Vaccines 2020, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Azuar, A.; Jin, W.; Mukaida, S.; Hussein, W.M.; Toth, I.; Skwarczynski, M. Recent advances in the development of peptide vaccines and their delivery systems against group a streptococcus. Vaccines 2019, 7, 58. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, L.; Gong, W. In silico analysis of peptide-based biomarkers for the diagnosis and prevention of latent tuberculosis infection. Front. Microbiol. 2022, 13, 947852. [Google Scholar] [CrossRef]
- Gong, W.; Liang, Y.; Wang, J.; Liu, Y.; Xue, Y.; Mi, J.; Li, P.; Wang, X.; Wang, L.; Wu, X. Prediction of Th1 and cytotoxic T lymphocyte epitopes of Mycobacterium tuberculosis and evaluation of their potential in the diagnosis of tuberculosis in a mouse model and in humans. Microbiol. Spectr. 2022, 10, e0143822. [Google Scholar] [CrossRef]
- Gong, W.; Wu, X. Differential diagnosis of latent tuberculosis infection and active tuberculosis: A key to a successful tuberculosis control strategy. Front. Microbiol. 2021, 12, 745592. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Liu, Y.; Xue, Y.; Cheng, P.; Wang, J.; Lian, J.; Gong, W. Developing a multiepitope vaccine for the prevention of SARS-CoV-2 and monkeypox virus co-infection: A reverse vaccinology analysis. Int. Immunopharmacol. 2023, 115, 109728. [Google Scholar] [CrossRef]
- Gong, W.; Liang, Y.; Mi, J.; Jia, Z.; Xue, Y.; Wang, J.; Wang, L.; Zhou, Y.; Sun, S.; Wu, X. Peptides-based vaccine MP3RT induced protective immunity against Mycobacterium tuberculosis infection in a humanized mouse model. Front. Immunol. 2021, 12, 666290. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, I.; Bangov, I.; Flower, D.R.; Doytchinova, I. AllerTOP v.2--a server for in silico prediction of allergens. J. Mol. Model. 2014, 20, 2278. [Google Scholar] [CrossRef]
- Dimitrov, I.; Naneva, L.; Doytchinova, I.; Bangov, I. AllergenFP: Allergenicity prediction by descriptor fingerprints. Bioinformatics 2014, 30, 846–851. [Google Scholar] [CrossRef]
- Cheng, P.; Xue, Y.; Wang, J.; Jia, Z.; Wang, L.; Gong, W. Evaluation of the consistence between the results of immunoinformatics predictions and real-world animal experiments of a new tuberculosis vaccine MP3RT. Front. Cell Infect. Microbiol. 2022, 12, 1047306. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Yuen, R.; Kuniholm, J.; Lisk, C.; Wetzler, L.M. Neisserial PorB immune enhancing activity and use as a vaccine adjuvant. Hum. Vaccin. Immunother. 2019, 15, 2778–2781. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, H.; Liang, H.; Zhao, X.; Tang, W.; Wei, M.; Li, Y.; Zhang, J.; Yu, X.; Chen, G.; et al. Lactobacillus plantarum RS-09 induces M1-type macrophage immunity against Salmonella typhimurium challenge via the TLR2/NF-κB signalling pathway. Front. Pharmacol. 2022, 13, 832245. [Google Scholar] [CrossRef]
- Soltan, M.A.; Elbassiouny, N.; Gamal, H.; Elkaeed, E.B.; Eid, R.A.; Eldeen, M.A.; Al-Karmalawy, A.A. In silico prediction of a multitope vaccine against Moraxella catarrhalis: Reverse vaccinology and immunoinformatics. Vaccines 2021, 9, 669. [Google Scholar] [CrossRef]
- Magnan, C.N.; Zeller, M.; Kayala, M.A.; Vigil, A.; Randall, A.; Felgner, P.L.; Baldi, P. High-throughput prediction of protein antigenicity using protein microarray data. Bioinformatics 2010, 26, 2936–2943. [Google Scholar] [CrossRef]
- Calis, J.J.; Maybeno, M.; Greenbaum, J.A.; Weiskopf, D.; De Silva, A.D.; Sette, A.; Keşmir, C.; Peters, B. Properties of MHC class I presented peptides that enhance immunogenicity. PLoS Comput. Biol. 2013, 9, e1003266. [Google Scholar] [CrossRef]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P. In silico approach for predicting toxicity of peptides and proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [PubMed]
- Barman, A.; Deb, B.; Chakraborty, S. Prediction of potential epitopes for peptide vaccine formulation against teschovirus a using immunoinformatics. Int. J. Pept. Res. Ther. 2020, 26, 1137–1146. [Google Scholar] [CrossRef]
- Cheng, J.; Randall, A.Z.; Sweredoski, M.J.; Baldi, P. SCRATCH: A protein structure and structural feature prediction server. Nucleic Acids Res. 2005, 33, W72-76. [Google Scholar] [CrossRef]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED protein structure prediction server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef]
- Long, S.; Tian, P. Protein secondary structure prediction with context convolutional neural network. RSC Adv. 2019, 9, 38391–38396. [Google Scholar] [CrossRef]
- Behera, S.K.; Sabarinath, T.; Mishra, P.K.K.; Deneke, Y.; Kumar, A.; ChandraSekar, S.; Senthilkumar, K.; Verma, M.; Ganesh, B.; Gurav, A.; et al. Immunoinformatic study of recombinant liga/bcon1-5 antigen and evaluation of its diagnostic potential in primary and secondary binding tests for serodiagnosis of porcine leptospirosis. Pathogens 2021, 10, 1082. [Google Scholar] [CrossRef]
- Haron, F.N.; Azazi, A.; Chua, K.H.; Lim, Y.A.L.; Lee, P.C.; Chew, C.H. In silico structural modeling and quality assessment of Plasmodium knowlesi apical membrane antigen 1 using comparative protein models. Trop. Biomed. 2022, 39, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Muccee, F.; Ghazanfar, S.; Ajmal, W.; Al-Zahrani, M. In-silico characterization of estrogen reactivating β-glucuronidase enzyme in git associated microbiota of normal human and breast cancer patients. Genes 2022, 13, 1545. [Google Scholar] [CrossRef] [PubMed]
- Setlur, A.S.; Pandey, S.; Sarkar, M.; Niranjan, V. Comprehensive molecular interaction studies to construe the repellent/kill activity of geraniol during binding event against Aedes aegypti proteins. Mol. Biotechnol. 2022, 65, 726–740. [Google Scholar] [CrossRef]
- Vajda, S.; Yueh, C.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Kozakov, D. New additions to the ClusPro server motivated by CAPRI. Proteins 2017, 85, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Rapin, N.; Lund, O.; Bernaschi, M.; Castiglione, F. Computational immunology meets bioinformatics: The use of prediction tools for molecular binding in the simulation of the immune system. PLoS ONE 2010, 5, e9862. [Google Scholar] [CrossRef]
- Cheng, P.; Jiang, F.; Wang, G.; Wang, J.; Xue, Y.; Wang, L.; Gong, W. Bioinformatics analysis and consistency verification of a novel tuberculosis vaccine candidate HP13138PB. Front Immunol. 2023, 14, 1102578. [Google Scholar] [CrossRef]
- Wang, J.; Mi, J.; Liang, Y.; Wu, X.Q.; Zhang, J.X.; Liu, Y.P.; Wang, L.; Xue, Y.; Shi, Y.C.; Gong, W.P. Transcriptomic analysis of tuberculosis peptide-based vaccine MP3RT in humanized mice. Zhonghua Jie He He Hu Xi Za Zhi 2022, 45, 894–903. [Google Scholar]
- Kumar Nathella, P.; Babu, S. Influence of diabetes mellitus on immunity to human tuberculosis. Immunology 2017, 152, 13–24. [Google Scholar] [CrossRef]
- Khademi, F.; Taheri, R.A.; Momtazi-Borojeni, A.A.; Farnoosh, G.; Johnston, T.P.; Sahebkar, A. Potential of cationic liposomes as adjuvants/delivery systems for tuberculosis subunit vaccines. Rev. Physiol. Biochem. Pharmacol. 2018, 175, 47–69. [Google Scholar] [CrossRef]
- Saraav, I.; Singh, S.; Sharma, S. Outcome of Mycobacterium tuberculosis and Toll-like receptor interaction: Immune response or immune evasion? Immunol. Cell Biol. 2014, 92, 741–746. [Google Scholar] [CrossRef]
- Wani, B.A.; Shehjar, F.; Shah, S.; Koul, A.; Yusuf, A.; Farooq, M.; Mir, M.I.; Singh, R.; Afroze, D. Role of genetic variants of Vitamin D receptor, Toll-like receptor 2 and Toll-like receptor 4 in extrapulmonary tuberculosis. Microb. Pathog. 2021, 156, 104911. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.; Dietzold, J.; Salgame, P. Vaccine-mediated immunity to experimental Mycobacterium tuberculosis is not impaired in the absence of Toll-like receptor 9. Cell Immunol. 2016, 302, 11–18. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.; Salgame, P. Toll-like receptor 2 in host defense against Mycobacterium tuberculosis: To be or not to be-that is the question. Curr. Opin. Immunol. 2016, 42, 76–82. [Google Scholar] [CrossRef]
- Sepehri, Z.; Kiani, Z.; Kohan, F.; Ghavami, S. Toll-like receptor 4 as an immune receptor against Mycobacterium tuberculosis: A systematic review. Lab. Med. 2019, 50, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- Khan, M.; Khan, S.; Ali, A.; Akbar, H.; Sayaf, A.M.; Khan, A.; Wei, D.Q. Immunoinformatics approaches to explore Helicobacter Pylori proteome (virulence factors) to design B and T cell multi-epitope subunit vaccine. Sci. Rep. 2019, 9, 13321. [Google Scholar] [CrossRef] [PubMed]
- Shamriz, S.; Ofoghi, H.; Moazami, N. Effect of linker length and residues on the structure and stability of a fusion protein with malaria vaccine application. Comput. Biol. Med. 2016, 76, 24–29. [Google Scholar] [CrossRef]
- Abdollahi, S.; Raoufi, Z.; Fakoor, M.H. Physicochemical and structural characterization, epitope mapping and vaccine potential investigation of a new protein containing Tetratrico Peptide Repeats of Acinetobacter baumannii: An in-silico and in-vivo approach. Mol. Immunol. 2021, 140, 22–34. [Google Scholar] [CrossRef]
- Agrawal, S.; Jana, U.K.; Kango, N. Heterologous expression and molecular modelling of L-asparaginase from Bacillus subtilis ETMC-2. Int. J. Biol. Macromol. 2021, 192, 28–37. [Google Scholar] [CrossRef]
- Qing, R.; Hao, S.; Smorodina, E.; Jin, D.; Zalevsky, A.; Zhang, S. Protein design: From the aspect of water solubility and stability. Chem. Rev. 2022, 122, 14085–14179. [Google Scholar] [CrossRef]
- Bhatnager, R.; Bhasin, M.; Arora, J.; Dang, A.S. Epitope based peptide vaccine against SARS-COV2: An immune-informatics approach. J. Biomol. Struct. Dyn. 2021, 39, 5690–5705. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Dalpiaz, A.; Varani, K.; Gilli, P.; Gilli, G. Can thermodynamic measurements of receptor binding yield information on drug affinity and efficacy? Biochem. Pharmacol. 2000, 60, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Csapó, E.; Bogár, F.; Juhász, Á.; Sebők, D.; Szolomájer, J.; Tóth, G.K.; Majláth, Z.; Vécsei, L.; Dékány, I. Determination of binding capacity and adsorption enthalpy between Human Glutamate Receptor (GluR1) peptide fragments and kynurenic acid by surface plasmon resonance experiments. Part 2: Interaction of GluR1270-300 with KYNA. Colloid Surf. B 2015, 133, 66–72. [Google Scholar] [CrossRef]
- Mayer-Barber, K.D.; Barber, D.L. Innate and adaptive cellular immune responses to Mycobacterium tuberculosis infection. Cold Spring Harb. Perspect. Med. 2015, 5, a018424. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.S.; Conti, B.J.; Fraga-Silva, T.F.C.; Almeida, F.; Bonato, V.L.D. Interplay between alveolar epithelial and dendritic cells and Mycobacterium tuberculosis. J. Leukoc. Biol. 2020, 108, 1139–1156. [Google Scholar] [CrossRef]
- Kim, H.; Shin, S.J. Pathological and protective roles of dendritic cells in Mycobacterium tuberculosis infection: Interaction between host immune responses and pathogen evasion. Front. Cell Infect. Microbiol. 2022, 12, 891878. [Google Scholar] [CrossRef] [PubMed]
- De Libero, G.; Singhal, A.; Lepore, M.; Mori, L. Nonclassical T cells and their antigens in tuberculosis. Csh. Perspect. Med. 2014, 4, a018473. [Google Scholar] [CrossRef]
- Lin, P.L.; Flynn, J.L. CD8 T cells and Mycobacterium tuberculosis infection. Semin. Immunopathol. 2015, 37, 239–249. [Google Scholar] [CrossRef]
- Khan, N.; Vidyarthi, A.; Amir, M.; Mushtaq, K.; Agrewala, J.N. T-cell exhaustion in tuberculosis: Pitfalls and prospects. Crit. Rev. Microbiol. 2017, 43, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.J.; Barreira-Silva, P.; Boyce, S.; Powers, J.; Cavallo, K.; Behar, S.M. CD4 T cell help prevents CD8 T cell exhaustion and promotes control of Mycobacterium tuberculosis infection. Cell. Rep. 2021, 36, 109696. [Google Scholar] [CrossRef]
- Mpande, C.A.M.; Rozot, V.; Mosito, B.; Musvosvi, M.; Dintwe, O.B.; Bilek, N.; Hatherill, M.; Scriba, T.J.; Nemes, E. Immune profiling of Mycobacterium tuberculosis-specific T cells in recent and remote infection. EBioMedicine 2021, 64, 103233. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Allele | Peptide | Adjusted Rank | Antigenicity Score | IFN-γ Score | Immunogenicity Score | ABC Score | Allergenicity * | Toxin Pred | |
|---|---|---|---|---|---|---|---|---|---|---|
| HTL | ||||||||||
| Rv1736c | HLA-DQA1*02:01, DQB1*05:02 | GAAFSWYTYSPTRVR | 0.14 | 0.9205 | 0.43799388 | NA | NA | NO | NO | NT |
| HLA-DQA1*05:01, DQB1*02:01 | DFLPVVLEFAATVDPEA | 0.31 | 1.0044 | 0.60486444 | NA | NA | NO | NO | NT | |
| HLA-DRB1*15:02 | LYYQIHVLIGLALFAL | 0.3 | 0.9211 | 0.43948812 | NA | NA | NO | NO | NT | |
| HLA-DRB1*14:04 | LYESRLLRIASPMFHFGI | 0.38 | 0.7205 | 0.33801445 | NA | NA | NO | NO | NT | |
| HLA-DRB1*15:02 | MMGELFWTVVPYVTMTI | 0.39 | 0.7048 | 0.34250203 | NA | NA | NO | NO | NT | |
| Rv1737c | HLA-DQA1*06:01, DQB1*03:03 | AVTLASILPVLAV | 0.34 | 0.8199 | 0.38907504 | NA | NA | NO | NO | NT |
| HLA-DRB1*15:02 | ATMGSYALLVFFGLFL | 0.46 | 0.856 | 0.72004245 | NA | NA | NO | NO | NT | |
| Rv1980c | HLA-DRB3*02:02 | PAYNINISLPSYYPDQ | 0.08 | 1.3614 | 2 | NA | NA | NO | NO | NT |
| Rv1981c | HLA-DRB1*15:02 | FLFYSGFYLPMYW | 0.08 | 1.3283 | 1 | NA | NA | NO | NO | NT |
| HLA-DQA1*02:01, DQB1*05:02 | 0.12 | 1.3283 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DQA1*01:01, DQB1*05:01 | 0.16 | 1.3283 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DPA1*01, DPB1*04:01 | 0.19 | 1.3283 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DRB1*15:02 | SFLFYSGFYLPMYW | 0.06 | 1.072 | 1 | NA | NA | NO | NO | NT | |
| HLA-DPA1*01, DPB1*04:01 | 0.14 | 1.072 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DQA1*02:01, DQB1*05:02 | 0.16 | 1.072 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DQA1*01:01, DQB1*05:01 | 0.19 | 1.072 | 1 | NA | NA | NO | NO | NT | ||
| HLA-DPA1*01:03, DPB1*02:01 | 0.31 | 1.072 | 1 | NA | NA | NO | NO | NT | ||
| Rv2659c | HLA-DQA1*05:01, DQB1*03:01 | DLRVHDLRHSGAVLAAST | 0.44 | 0.9292 | 2 | NA | NA | NO | NO | NT |
| Rv3429 | HLA-DQA1*02:01, DQB1*05:02 | QSTARFILAYLPRWQ | 0.28 | 0.7639 | 0.13606656 | NA | NA | NO | NO | NT |
| HLA-DRB3*02:02 | AAAEQLRLMYNSANMTAK | 0.41 | 0.7112 | 0.54792199 | NA | NA | NO | NO | NT | |
| Rv3873 | HLA-DQA1*03:01, DQB1*06:01 | VAPSVMPAAAAGSSAT | 0.31 | 1.0173 | 0.23735279 | NA | NA | NO | NO | NT |
| HLA-DQA1*03:01, DQB1*06:01 | GLSAAAAKLAGLVF | 0.43 | 0.7126 | 0.67278691 | NA | NA | NO | NO | NT | |
| HLA-DQA1*05:01, DQB1*03:01 | DTGAGARPAASPLAAPV | 0.15 | 0.8414 | 0.82589324 | NA | NA | NO | NO | NT | |
| Rv3879 | HLA-DQA1*02:01, DQB1*05:02 | TGREAAHLRAFRAYAAHS | 0.03 | 0.7317 | 1.4848492 | NA | NA | NO | NO | NT |
| HLA-DQA1*05:01, DQB1*03:01 | VTPAAASGVPGARAAAAA | 0.47 | 0.8376 | 1.1311669 | NA | NA | NO | NO | NT | |
| HLA-DQA1*02:01, DQB1*05:02 | YPVLAVQAWAAFHDMTLR | 0.44 | 0.7948 | 0.84318227 | NA | NA | NO | NO | NT | |
| CTL | ||||||||||
| Rv1736c | HLA-B*15:02, HLA-B*15:11 | ELFWTVVPY | NA | 1.3999 | NA | 0.36591 | NA | NO | NO | NT |
| Rv1737c | HLA-A*02:01 | GVAGTIFAV | NA | 0.9924 | NA | 0.33154 | NA | NO | NO | NT |
| Rv2031c | HLA-A*33:03 | EMKEGRYEVR | NA | 1.678 | NA | 0.21929 | NA | NO | NO | NT |
| Rv2626c | HLA-A*33:03 | NVMEEHQVRR | NA | 0.8767 | NA | 0.12879 | NA | NO | NO | NT |
| Rv2656c | HLA-C*06:02, HLA-C*07:02 | RRDAYIRRV | NA | 0.9325 | NA | 0.24217 | NA | NO | NO | NT |
| Rv2659c | HLA-A*11:01 | ASYTGPDGR | NA | 1.0059 | NA | 0.09894 | NA | NO | NO | NT |
| Rv1511 | HLA-C*07:02, HLA-C*06:02 | LRPTEVDSL | NA | 1.2094 | NA | 0.09388 | NA | NO | NO | NT |
| Rv1980c | HLA-B*15:01 | GGTHPTTTY | NA | 1.7475 | NA | 0.12633 | NA | NO | NO | NT |
| Rv3872 | HLA-B*15:11 | EVSAQAATAF | NA | 0.8759 | NA | 0.00342 | NA | NO | NO | NT |
| Rv3425 | HLA-A*33:03 | EIAANREER | NA | 1.0439 | NA | 0.23749 | NA | NO | NO | NT |
| Rv3878 | HLA-A*33:03 | ETAAELAPR | NA | 1.0877 | NA | 0.16567 | NA | NO | NO | NT |
| Rv3879 | HLA-C*03:04, HLA-C*01:02, HLA-A*30:01 | ATHGANVSL | NA | 1.8216 | NA | 0.01479 | NA | NO | NO | NT |
| B cell | ||||||||||
| Rv1511 | NA | LIRRASTFNTSRIDHLYVDPHQPGARLFLHYGD | NA | NA | NA | NA | 0.77 | NO | NO | NT |
| Rv1737c | NA | LRDAPYFRPNADPVLPRLKAAA | NA | NA | NA | NA | 0.78 | NO | NO | NT |
| Rv1981c | NA | TLCSTKQIDDAFDWSEQNPYLQRKAQIIVDYY | NA | NA | NA | NA | 0.81 | NO | NO | NT |
| Rv2031c | NA | LEDEMKEGRYEVRAELPGVDPDKDV | NA | NA | NA | NA | 0.94 | NO | NO | NT |
| Rv2660c | NA | GVGTEQRNLSVVAPSQFTFSSRSPDFVDETAGQSWCAIL | NA | NA | NA | NA | 0.86 | NO | NO | NT |
| Rv2653c | NA | AGLNAPRRNRVGRQHGWPADVPSAEQRRAQRQRDLEA | NA | NA | NA | NA | 0.92 | NO | NO | NT |
| Rv3425 | NA | GLANAYNDTRRKVVPPEEIAANREERRR | NA | NA | NA | NA | 0.86 | NO | NO | NT |
| Rv3879 | NA | TDQRLLDLLPPAPVDVNPPGDERHMLWFELMK | NA | NA | NA | NA | 0.82 | NO | NO | NT |
| Population/Area | Class I | Class II | ||||
|---|---|---|---|---|---|---|
| Coverage a | Average_hit b | Pc90 c | Coverage a | Average_hit b | Pc90 c | |
| Central Africa | 57.38% | 1.32 | 0.23 | 82.75% | 2.79 | 0.58 |
| East Africa | 61.10% | 1.2 | 0.26 | 89.22% | 3.63 | 0.93 |
| East Asia | 85.63% | 2.29 | 0.7 | 73.85% | 2.72 | 0.38 |
| Europe | 82.00% | 1.76 | 0.56 | 99.06% | 4.41 | 1.4 |
| North Africa | 65.52% | 1.42 | 0.29 | 80.01% | 4.04 | 0.5 |
| North America | 82.17% | 1.88 | 0.56 | 99.93% | 5.05 | 2.94 |
| Northeast Asia | 92.53% | 2.66 | 1.12 | 97.97% | 5.07 | 2.19 |
| Oceania | 76.33% | 1.38 | 0.42 | 97.55% | 4.78 | 2.05 |
| South America | 65.56% | 1.29 | 0.29 | 98.12% | 4.69 | 2.37 |
| South Asia | 70.59% | 1.83 | 0.34 | 97.75% | 5.68 | 1.65 |
| Southeast Asia | 91.15% | 2.63 | 1.05 | 88.86% | 3.84 | 0.9 |
| Southwest Asia | 64.29% | 1.34 | 0.28 | 83.80% | 4.01 | 0.62 |
| West Africa | 59.42% | 1.33 | 0.25 | 88.35% | 4.03 | 0.86 |
| World | 82.24% | 1.88 | 0.56 | 93.71% | 4.16 | 1.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, F.; Peng, C.; Cheng, P.; Wang, J.; Lian, J.; Gong, W. PP19128R, a Multiepitope Vaccine Designed to Prevent Latent Tuberculosis Infection, Induced Immune Responses In Silico and In Vitro Assays. Vaccines 2023, 11, 856. https://doi.org/10.3390/vaccines11040856
Jiang F, Peng C, Cheng P, Wang J, Lian J, Gong W. PP19128R, a Multiepitope Vaccine Designed to Prevent Latent Tuberculosis Infection, Induced Immune Responses In Silico and In Vitro Assays. Vaccines. 2023; 11(4):856. https://doi.org/10.3390/vaccines11040856
Chicago/Turabian StyleJiang, Fan, Cong Peng, Peng Cheng, Jie Wang, Jianqi Lian, and Wenping Gong. 2023. "PP19128R, a Multiepitope Vaccine Designed to Prevent Latent Tuberculosis Infection, Induced Immune Responses In Silico and In Vitro Assays" Vaccines 11, no. 4: 856. https://doi.org/10.3390/vaccines11040856