

A Multistage Antigen Complex Epera013 Promotes Efficient and Comprehensive Immune Responses in BALB/c Mice
 , , and
, , and
Abstract
:1. Introduction
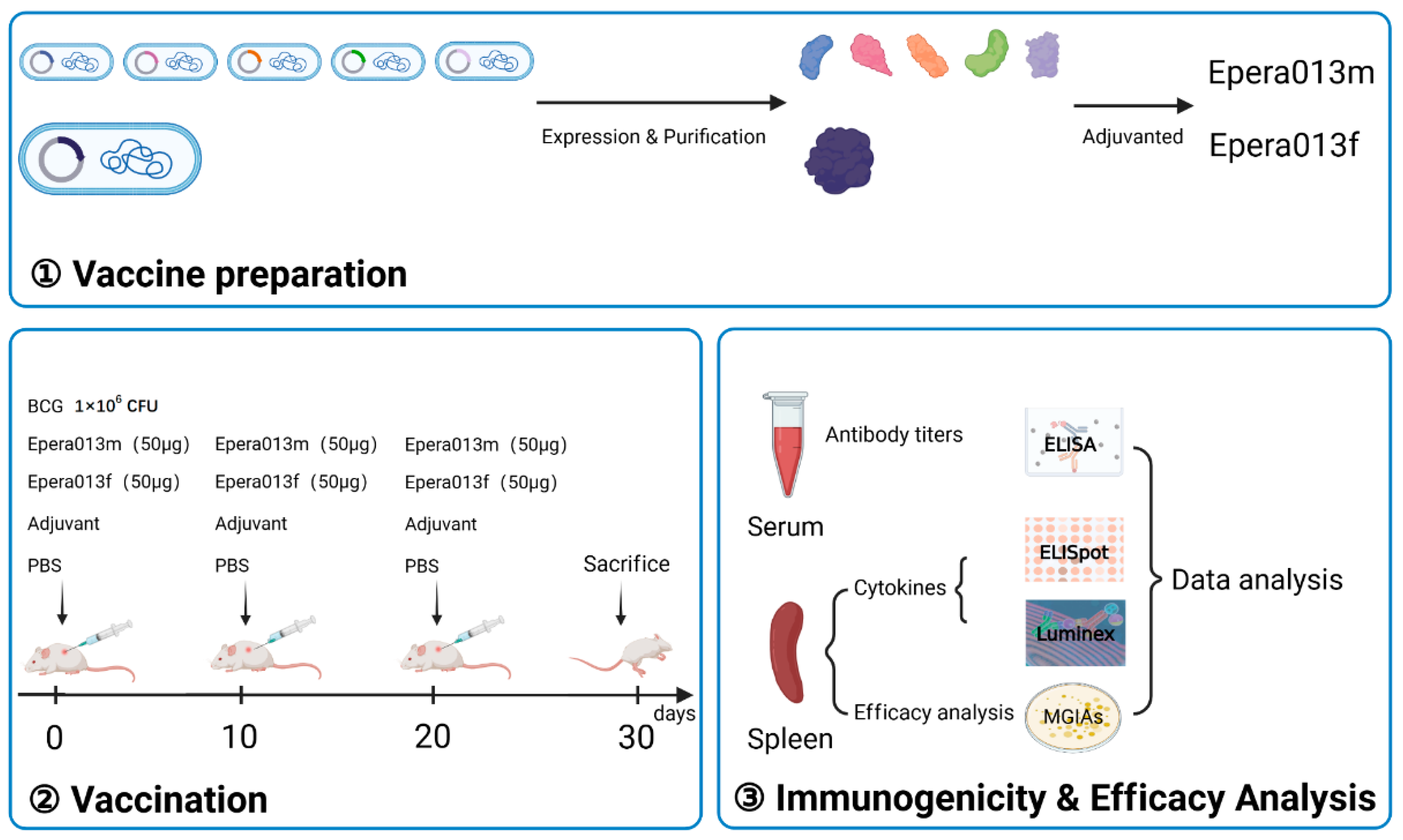
2. Materials and Methods
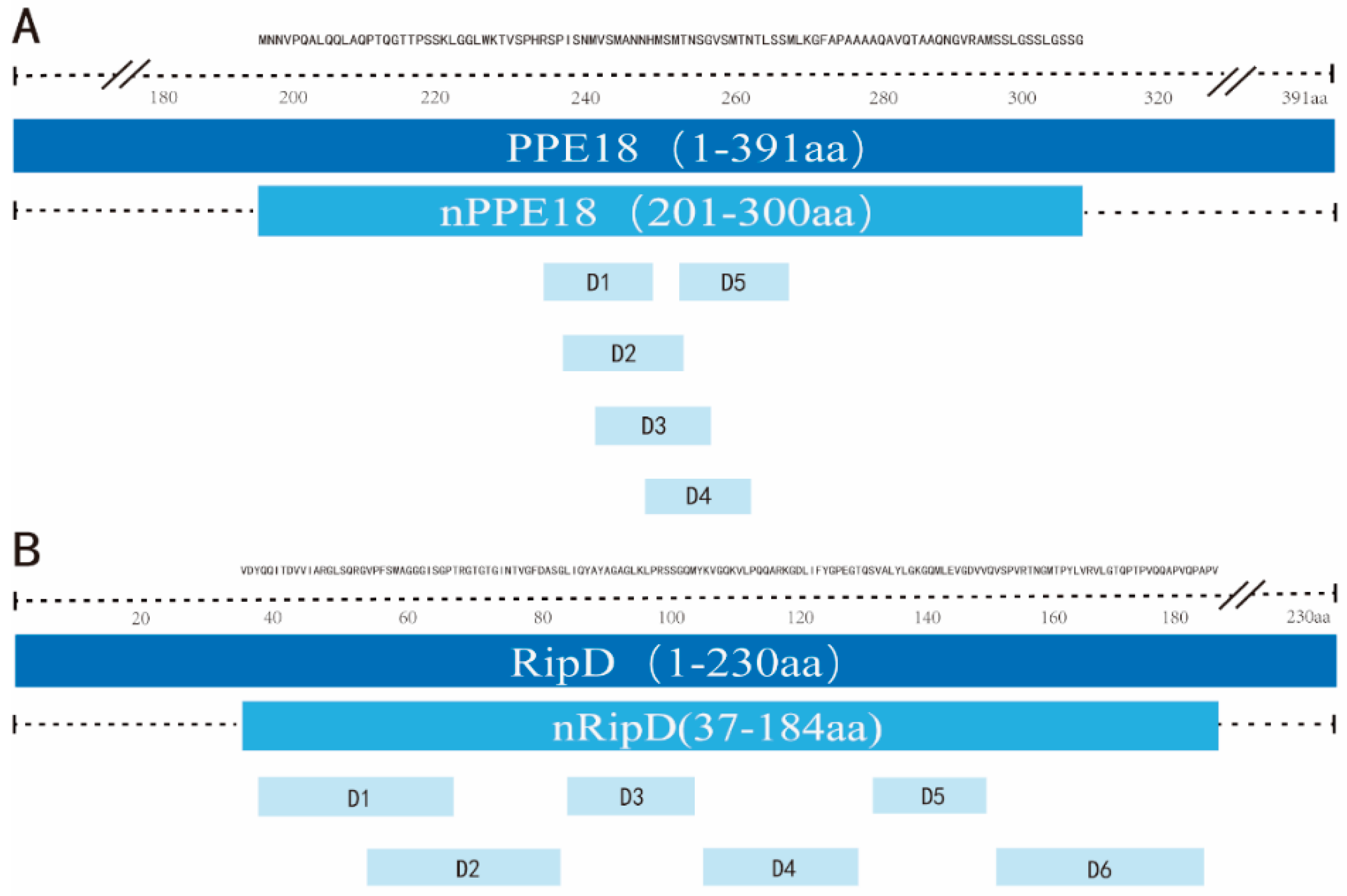
2.1. T-cell Epitope Prediction
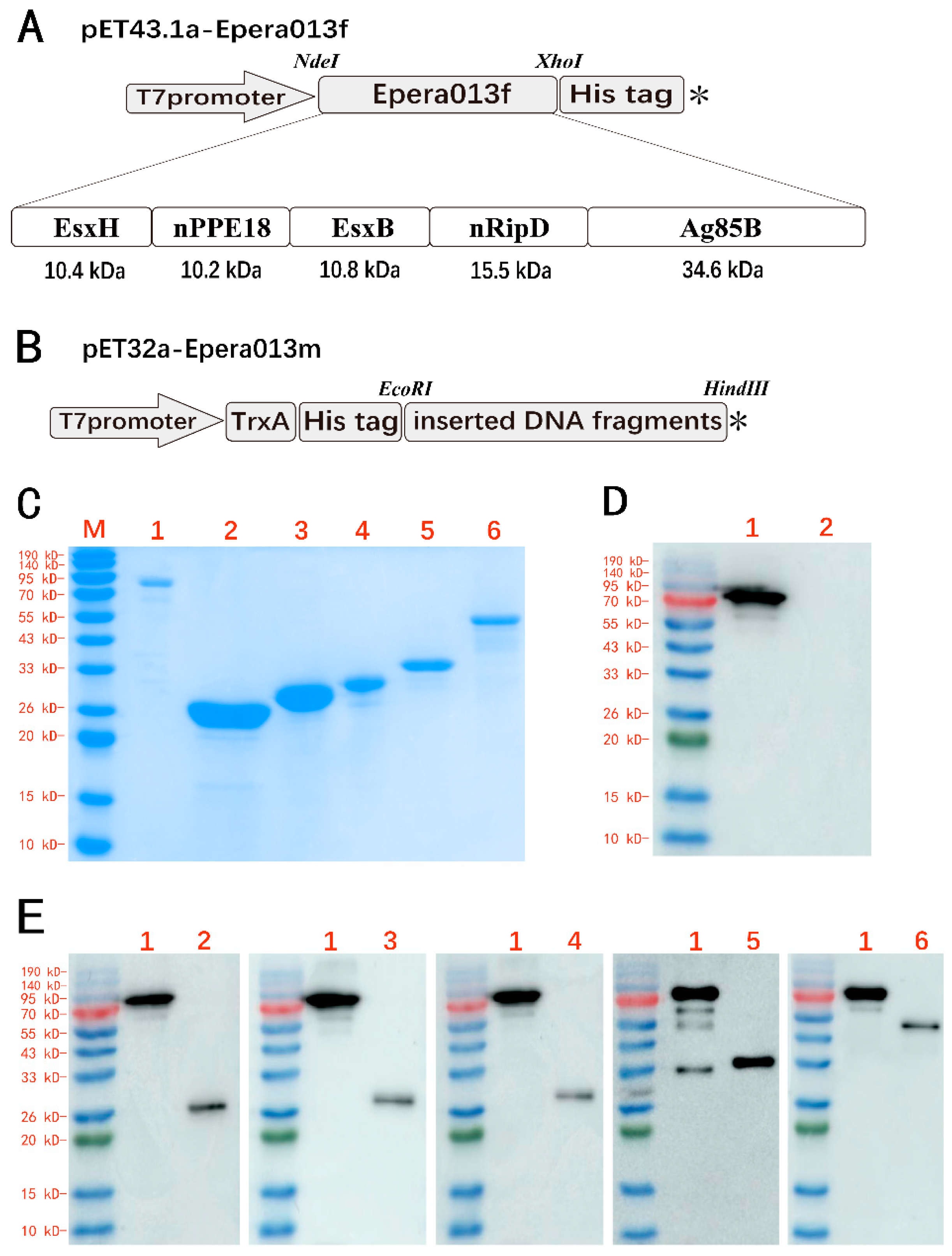
2.2. Cloning and Purification of Epera013f and Five Individual Antigen Components of Epera013m
2.3. Western Blotting Assay
2.4. Culture Condition and Preparation of Mycobacterium Strains
2.5. Animals and Immunization Strategy
2.6. Humoral Immunogenicity Evaluation
2.7. Lymphocyte Preparation
2.8. Antigen Specific Cytokines Determination
2.9. Ex Vivo Mycobacterial Growth Inhibition Assays
2.10. Statistical Analysis
3. Results
3.1. T-cell Epitope Prediction
3.2. Expression and Purification of the Recombinant Proteins
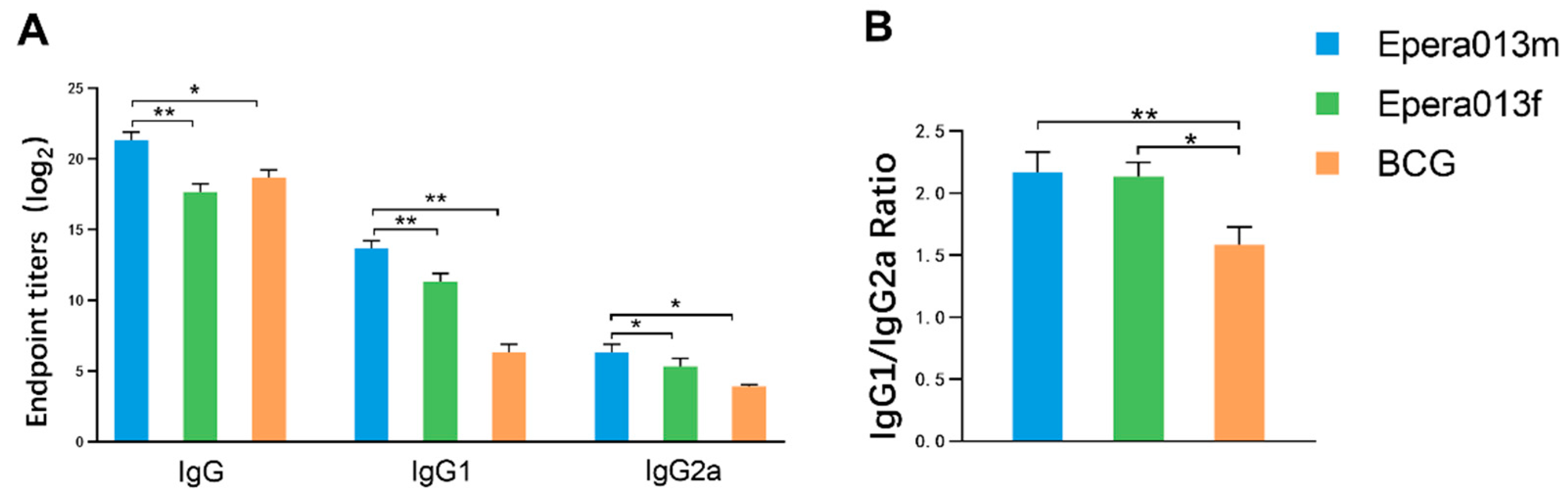
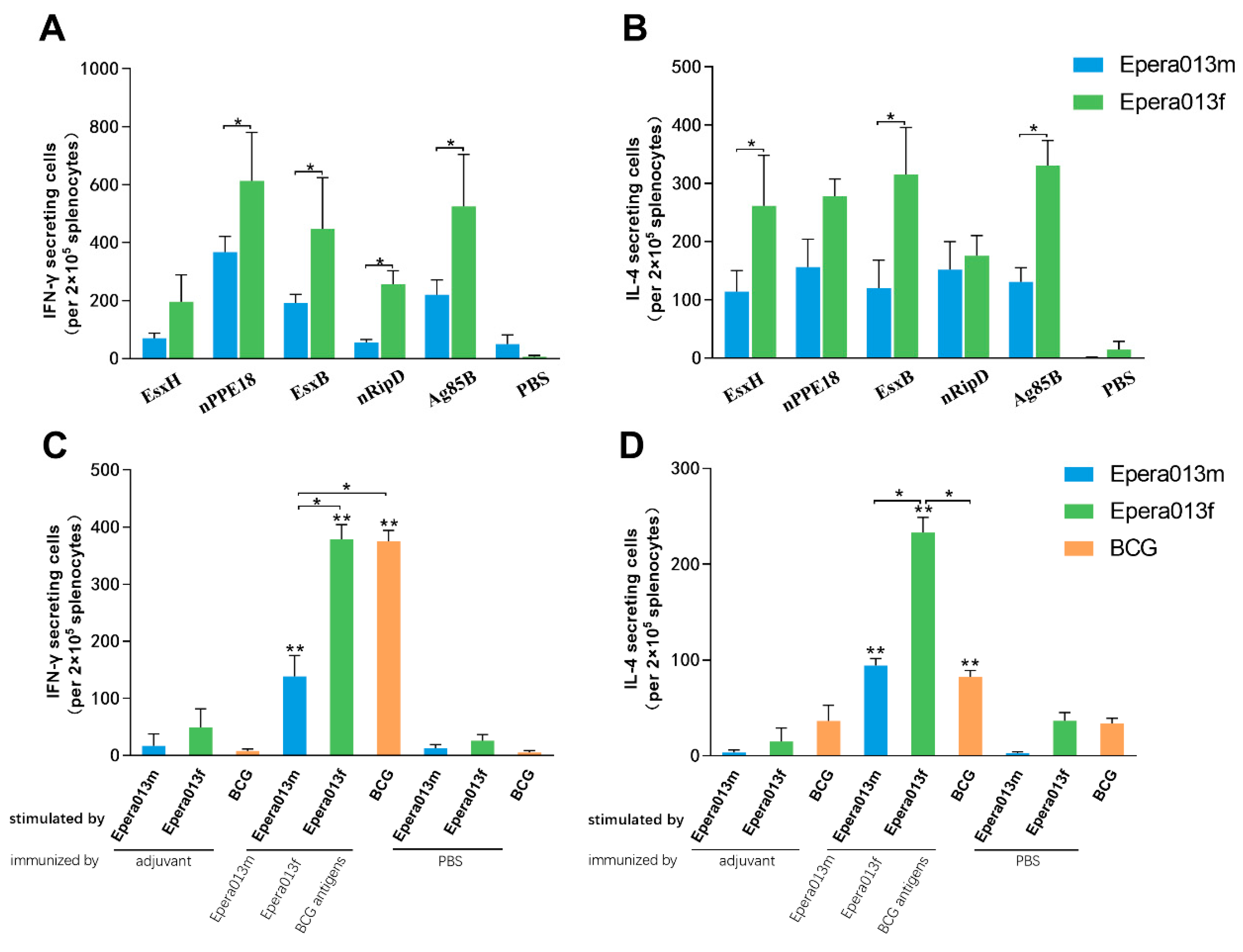
3.3. Robust Humoral Immune Responses Were Induced by Epera013f and Epera013m
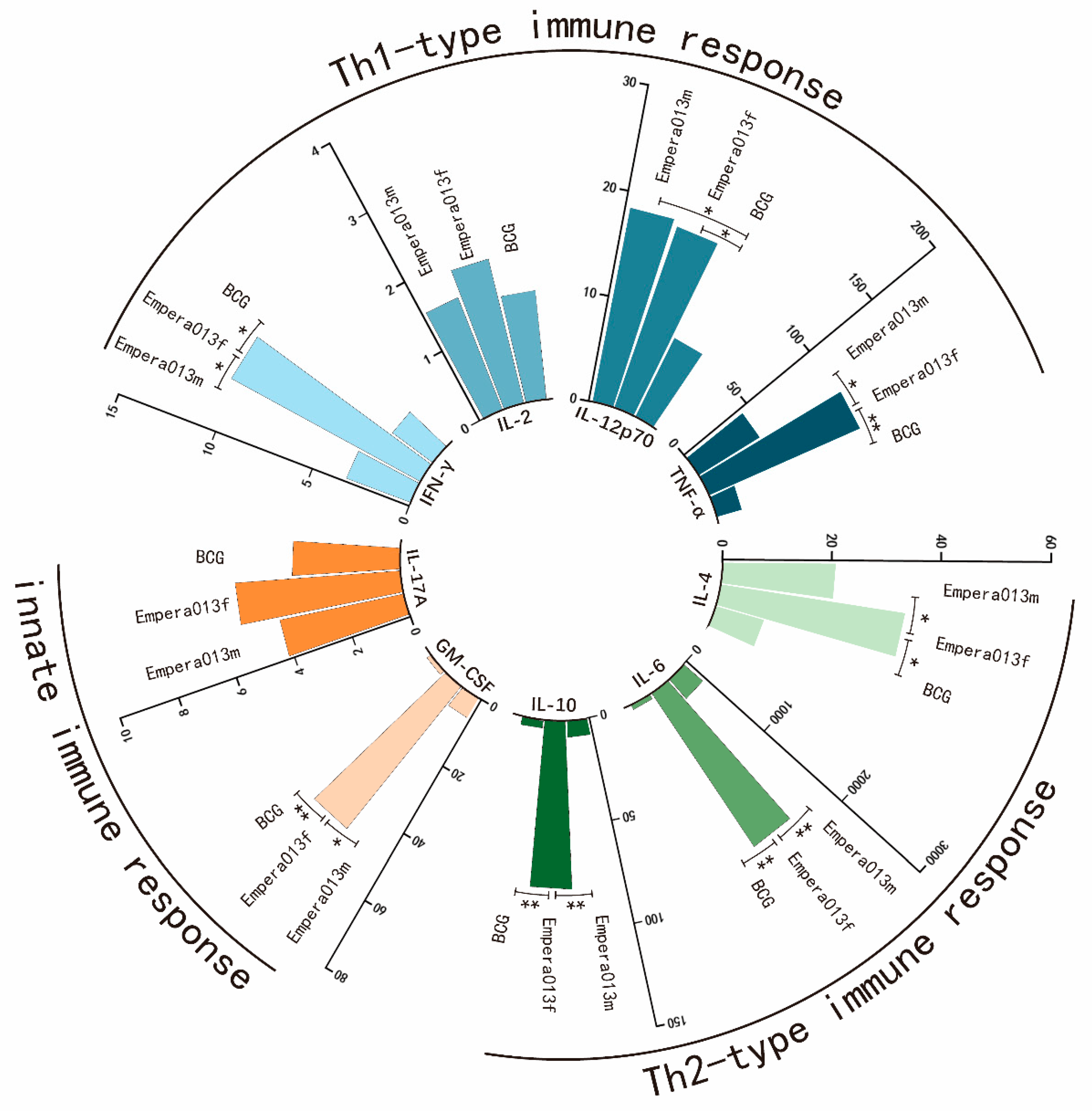
3.4. Th1, Th2 and Innate Immune Response Equilibrated Immune Status Was Generated by Epera013f
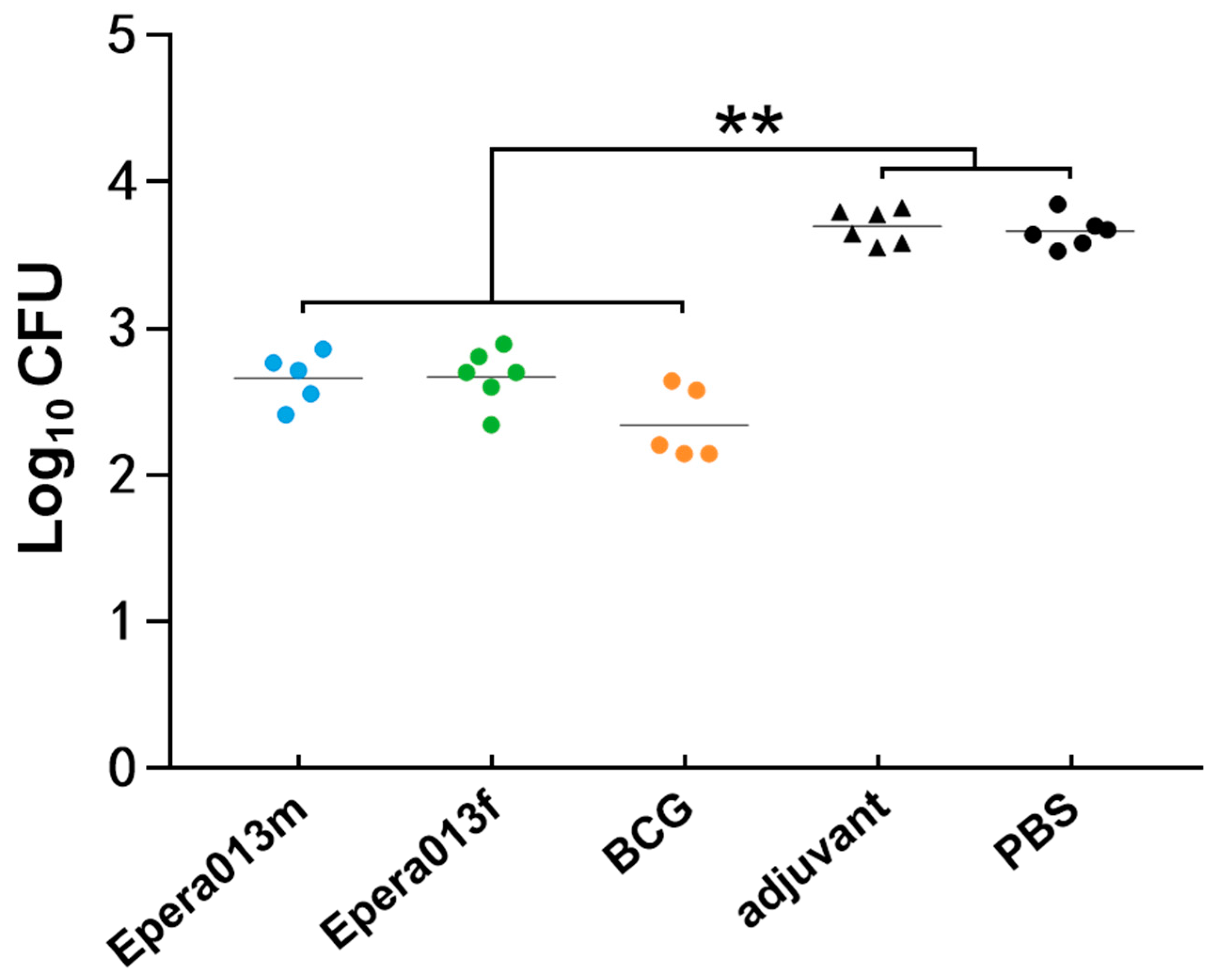
3.5. Mycobacterial Growth Inhibiting Capacity of Murine Splenocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- McShane, H. Tuberculosis vaccines: Beyond bacille Calmette-Guerin. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2782–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.; Dorman, S.E. Latent Tuberculosis Infection. N. Engl. J. Med. 2021, 385, 2271–2280. [Google Scholar] [CrossRef]
- Lange, C.; Chesov, D.; Heyckendorf, J.; Leung, C.C.; Udwadia, Z.; Dheda, K. Drug-resistant tuberculosis: An update on disease burden, diagnosis and treatment. Respirology 2018, 23, 656–673. [Google Scholar] [CrossRef] [PubMed]
- Martinson, N.A.; Barnes, G.L.; Moulton, L.H.; Msandiwa, R.; Hausler, H.; Ram, M.; McIntyre, J.A.; Gray, G.E.; Chaisson, R.E. New regimens to prevent tuberculosis in adults with HIV infection. N. Engl. J. Med. 2011, 365, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Dockrell, H.M.; Ottenhoff, T.H.M.; Evans, T.G.; Zhang, Y. Tuberculosis vaccines: Opportunities and challenges. Respirology 2018, 23, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazier, B.; McShane, H. Towards new TB vaccines. Semin. Immunopathol. 2020, 42, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Garg, S.; Vedi, S.; Kunimoto, D.Y.; Kumar, R.; Agrawal, B. Future Path Toward TB Vaccine Development: Boosting BCG or Re-educating by a New Subunit Vaccine. Front. Immunol. 2018, 9, 2371. [Google Scholar] [CrossRef] [Green Version]
- Bellini, C.; Horvati, K. Recent Advances in the Development of Protein- and Peptide-Based Subunit Vaccines against Tuberculosis. Cells 2020, 9, 2673. [Google Scholar] [CrossRef]
- Jenum, S.; Tonby, K.; Rueegg, C.S.; Ruhwald, M.; Kristiansen, M.P.; Bang, P.; Olsen, I.C.; Sellaeg, K.; Rostad, K.; Mustafa, T.; et al. A Phase I/II randomized trial of H56:IC31 vaccination and adjunctive cyclooxygenase-2-inhibitor treatment in tuberculosis patients. Nat. Commun. 2021, 12, 6774. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C.; Quaife, M.; Weerasuriya, C.; Gomez, G.B.; Sumner, T.; Bozzani, F.; White, R.G. Cost-effectiveness of routine adolescent vaccination with an M72/AS01E-like tuberculosis vaccine in South Africa and India. Nat. Commun. 2022, 13, 602. [Google Scholar] [CrossRef]
- Bettencourt, P.; Muller, J.; Nicastri, A.; Cantillon, D.; Madhavan, M.; Charles, P.D.; Fotso, C.B.; Wittenberg, R.; Bull, N.; Pinpathomrat, N.; et al. Identification of antigens presented by MHC for vaccines against tuberculosis. NPJ Vaccines 2020, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stylianou, E.; Harrington-Kandt, R.; Beglov, J.; Bull, N.; Pinpathomrat, N.; Swarbrick, G.M.; Lewinsohn, D.A.; Lewinsohn, D.M.; McShane, H. Identification and Evaluation of Novel Protective Antigens for the Development of a Candidate Tuberculosis Subunit Vaccine. Infect. Immun. 2018, 86, e00014–e00018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, X.; Fan, X.; Wang, R.; Deng, Y.; Chen, Z.; Li, N.; Yan, Y.; Li, X.; Liu, H.; Li, G.; et al. High Immunogenicity of a T-Cell Epitope-Rich Recombinant Protein Rv1566c-444 From Mycobacterium tuberculosis in Immunized BALB/c Mice, Despite Its Low Diagnostic Sensitivity. Front. Immunol. 2022, 13, 824415. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, S.; Xu, Y.; Zheng, H.; Xiao, T.; Li, Y.; Chen, X.; Huang, M.; Zhang, H.; Fang, X.; et al. Identification and evaluation of the novel immunodominant antigen Rv2351c from Mycobacterium tuberculosis. Emerg. Microbes Infect. 2017, 6, e48. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Jiang, Y.; Dou, X.; Wang, H.; Zhao, X.; Zhang, W.; Wan, L.; Zhang, Z.; Chen, C.; Wan, K. pstS1 polymorphisms of Mycobacterium tuberculosis strains may reflect ongoing immune evasion. Tuberculosis 2013, 93, 475–481. [Google Scholar] [CrossRef]
- Horwitz, M.A.; Lee, B.W.; Dillon, B.J.; Harth, G. Protective immunity against tuberculosis induced by vaccination with major extracellular proteins of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 1995, 92, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, C.; Hoang, T.; Dietrich, J.; Cardona, P.J.; Izzo, A.; Dolganov, G.; Schoolnik, G.K.; Cassidy, J.P.; Billeskov, R.; Andersen, P. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat. Med. 2011, 17, 189–194. [Google Scholar] [CrossRef]
- Sutiwisesak, R.; Hicks, N.D.; Boyce, S.; Murphy, K.C.; Papavinasasundaram, K.; Carpenter, S.M.; Boucau, J.; Joshi, N.; Le Gall, S.; Fortune, S.M.; et al. A natural polymorphism of Mycobacterium tuberculosis in the esxH gene disrupts immunodomination by the TB10.4-specific CD8 T cell response. PLoS Pathog. 2020, 16, e1009000. [Google Scholar] [CrossRef]
- Nair, S.; Ramaswamy, P.A.; Ghosh, S.; Joshi, D.C.; Pathak, N.; Siddiqui, I.; Sharma, P.; Hasnain, S.E.; Mande, S.C.; Mukhopadhyay, S. The PPE18 of Mycobacterium tuberculosis interacts with TLR2 and activates IL-10 induction in macrophage. J. Immunol. 2009, 183, 6269–6281. [Google Scholar] [CrossRef] [Green Version]
- Lundegaard, C.; Lund, O.; Buus, S.; Nielsen, M. Major histocompatibility complex class I binding predictions as a tool in epitope discovery. Immunology 2010, 130, 309–318. [Google Scholar] [CrossRef]
- Zhao, W.; Sher, X. Systematically benchmarking peptide-MHC binding predictors: From synthetic to naturally processed epitopes. PLoS Comput. Biol. 2018, 14, e1006457. [Google Scholar] [CrossRef]
- Mei, S.; Li, F.; Leier, A.; Marquez-Lago, T.T.; Giam, K.; Croft, N.P.; Akutsu, T.; Smith, A.I.; Li, J.; Rossjohn, J.; et al. A comprehensive review and performance evaluation of bioinformatics tools for HLA class I peptide-binding prediction. Brief. Bioinform. 2020, 21, 1119–1135. [Google Scholar] [CrossRef] [PubMed]
- Krejci, J.; Pekarek, J.; Johanovsky, J.; Svejcar, J. Demonstration of the inflammatory activity of the supernatant of hypersensitive lymph node cells incubated with a high dose of antigen. Immunology 1969, 16, 677–684. [Google Scholar]
- Walzl, G.; Ronacher, K.; Hanekom, W.; Scriba, T.J.; Zumla, A. Immunological biomarkers of tuberculosis. Nat. Rev. Immunol. 2011, 11, 343–354. [Google Scholar] [CrossRef]
- Jensen, C.; Lindebo Holm, L.; Svensson, E.; Aagaard, C.; Ruhwald, M. Optimisation of a murine splenocyte mycobacterial growth inhibition assay using virulent Mycobacterium tuberculosis. Sci. Rep. 2017, 7, 2830. [Google Scholar] [CrossRef] [Green Version]
- Ehrt, S.; Schnappinger, D.; Rhee, K.Y. Metabolic principles of persistence and pathogenicity in Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2018, 16, 496–507. [Google Scholar] [CrossRef]
- Voskuil, M.I.; Visconti, K.C.; Schoolnik, G.K. Mycobacterium tuberculosis gene expression during adaptation to stationary phase and low-oxygen dormancy. Tuberculosis 2004, 84, 218–227. [Google Scholar] [CrossRef]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Muttucumaru, D.G.; Roberts, G.; Hinds, J.; Stabler, R.A.; Parish, T. Gene expression profile of Mycobacterium tuberculosis in a non-replicating state. Tuberculosis 2004, 84, 239–246. [Google Scholar] [CrossRef]
- Bertholet, S.; Ireton, G.C.; Ordway, D.J.; Windish, H.P.; Pine, S.O.; Kahn, M.; Phan, T.; Orme, I.M.; Vedvick, T.S.; Baldwin, S.L.; et al. A defined tuberculosis vaccine candidate boosts BCG and protects against multidrug-resistant Mycobacterium tuberculosis. Sci. Transl. Med. 2010, 2, 53ra74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Garra, A.; Redford, P.S.; McNab, F.W.; Bloom, C.I.; Wilkinson, R.J.; Berry, M.P. The immune response in tuberculosis. Annu Rev. Immunol. 2013, 31, 475–527. [Google Scholar] [CrossRef]
- Pai, M.; Denkinger, C.M.; Kik, S.V.; Rangaka, M.X.; Zwerling, A.; Oxlade, O.; Metcalfe, J.Z.; Cattamanchi, A.; Dowdy, D.W.; Dheda, K.; et al. Gamma interferon release assays for detection of Mycobacterium tuberculosis infection. Clin. Microbiol. Rev. 2014, 27, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Trinchieri, G. Interleukin-12: A proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. Annu. Rev. Immunol. 1995, 13, 251–276. [Google Scholar] [CrossRef]
- Alcais, A.; Fieschi, C.; Abel, L.; Casanova, J.L. Tuberculosis in children and adults: Two distinct genetic diseases. J. Exp. Med. 2005, 202, 1617–1621. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Darrah, P.A.; Roederer, M. T-cell quality in memory and protection: Implications for vaccine design. Nat. Rev. Immunol. 2008, 8, 247–258. [Google Scholar] [CrossRef]
- Phuah, J.Y.; Mattila, J.T.; Lin, P.L.; Flynn, J.L. Activated B cells in the granulomas of nonhuman primates infected with Mycobacterium tuberculosis. Am. J. Pathol. 2012, 181, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Maglione, P.J.; Chan, J. How B cells shape the immune response against Mycobacterium tuberculosis. Eur. J. Immunol. 2009, 39, 676–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravesloot-Chavez, M.M.; Van Dis, E.; Stanley, S.A. The Innate Immune Response to Mycobacterium tuberculosis Infection. Annu. Rev. Immunol. 2021, 39, 611–637. [Google Scholar] [CrossRef]
- Higgins, D.M.; Sanchez-Campillo, J.; Rosas-Taraco, A.G.; Lee, E.J.; Orme, I.M.; Gonzalez-Juarrero, M. Lack of IL-10 alters inflammatory and immune responses during pulmonary Mycobacterium tuberculosis infection. Tuberculosis 2009, 89, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Achkar, J.M.; Casadevall, A. Antibody-mediated immunity against tuberculosis: Implications for vaccine development. Cell Host Microbe 2013, 13, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Koeken, V.; Verrall, A.J.; Netea, M.G.; Hill, P.C.; van Crevel, R. Trained innate immunity and resistance to Mycobacterium tuberculosis infection. Clin. Microbiol. Infect. 2019, 25, 1468–1472. [Google Scholar] [CrossRef] [Green Version]
- O′Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef]
- Bryson, B.D.; Rosebrock, T.R.; Tafesse, F.G.; Itoh, C.Y.; Nibasumba, A.; Babunovic, G.H.; Corleis, B.; Martin, C.; Keegan, C.; Andrade, P.; et al. Heterogeneous GM-CSF signaling in macrophages is associated with control of Mycobacterium tuberculosis. Nat. Commun. 2019, 10, 2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counoupas, C.; Ferrell, K.C.; Ashhurst, A.; Bhattacharyya, N.D.; Nagalingam, G.; Stewart, E.L.; Feng, C.G.; Petrovsky, N.; Britton, W.J.; Triccas, J.A. Mucosal delivery of a multistage subunit vaccine promotes development of lung-resident memory T cells and affords interleukin-17-dependent protection against pulmonary tuberculosis. NPJ Vaccines 2020, 5, 105. [Google Scholar] [CrossRef] [PubMed]
- Tanner, R.; Villarreal-Ramos, B.; Vordermeier, H.M.; McShane, H. The Humoral Immune Response to BCG Vaccination. Front. Immunol. 2019, 10, 1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodo, M.J.; Rozot, V.; Nemes, E.; Dintwe, O.; Hatherill, M.; Little, F.; Scriba, T.J. A comparison of antigen-specific T cell responses induced by six novel tuberculosis vaccine candidates. PLoS Pathog. 2019, 15, e1007643. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Teng, X.; Wang, X.; Fan, X.; Wu, Y.; Tian, M.; Zhou, Z.; Li, L. A Multistage Subunit Vaccine Effectively Protects Mice Against Primary Progressive Tuberculosis, Latency and Reactivation. EBioMedicine 2017, 22, 143–154. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Start at | End at | Length | ||
|---|---|---|---|---|
| nPPE18 | Nucleotide position | 601 | 900 | 300 bp |
| Amino acid position | 201 | 300 | 100 aa | |
| nRipD | Nucleotide position | 109 | 552 | 444 bp |
| Amino acid position | 37 | 184 | 148 aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, C.; Fan, X.; Wang, R.; Cao, B.; Yu, J.; Luan, X.; Li, G.; Jiang, Y.; Li, M.; Zhao, X.; et al. A Multistage Antigen Complex Epera013 Promotes Efficient and Comprehensive Immune Responses in BALB/c Mice. Vaccines 2023, 11, 609. https://doi.org/10.3390/vaccines11030609
Qian C, Fan X, Wang R, Cao B, Yu J, Luan X, Li G, Jiang Y, Li M, Zhao X, et al. A Multistage Antigen Complex Epera013 Promotes Efficient and Comprehensive Immune Responses in BALB/c Mice. Vaccines. 2023; 11(3):609. https://doi.org/10.3390/vaccines11030609
Chicago/Turabian StyleQian, Chengyu, Xueting Fan, Ruihuan Wang, Bin Cao, Jinjie Yu, Xiuli Luan, Guilian Li, Yi Jiang, Machao Li, Xiuqin Zhao, and et al. 2023. "A Multistage Antigen Complex Epera013 Promotes Efficient and Comprehensive Immune Responses in BALB/c Mice" Vaccines 11, no. 3: 609. https://doi.org/10.3390/vaccines11030609