
Heterologous Vector—mRNA Based SARS-CoV-2 Vaccination Strategy Appears Superior to a Homologous Vector—Based Vaccination Scheme in German Healthcare Workers Regarding Humoral SARS-CoV-2 Response Indicating a High Boosting Effect by mRNA Vaccines
 , ,
, ,
Abstract
:1. Introduction
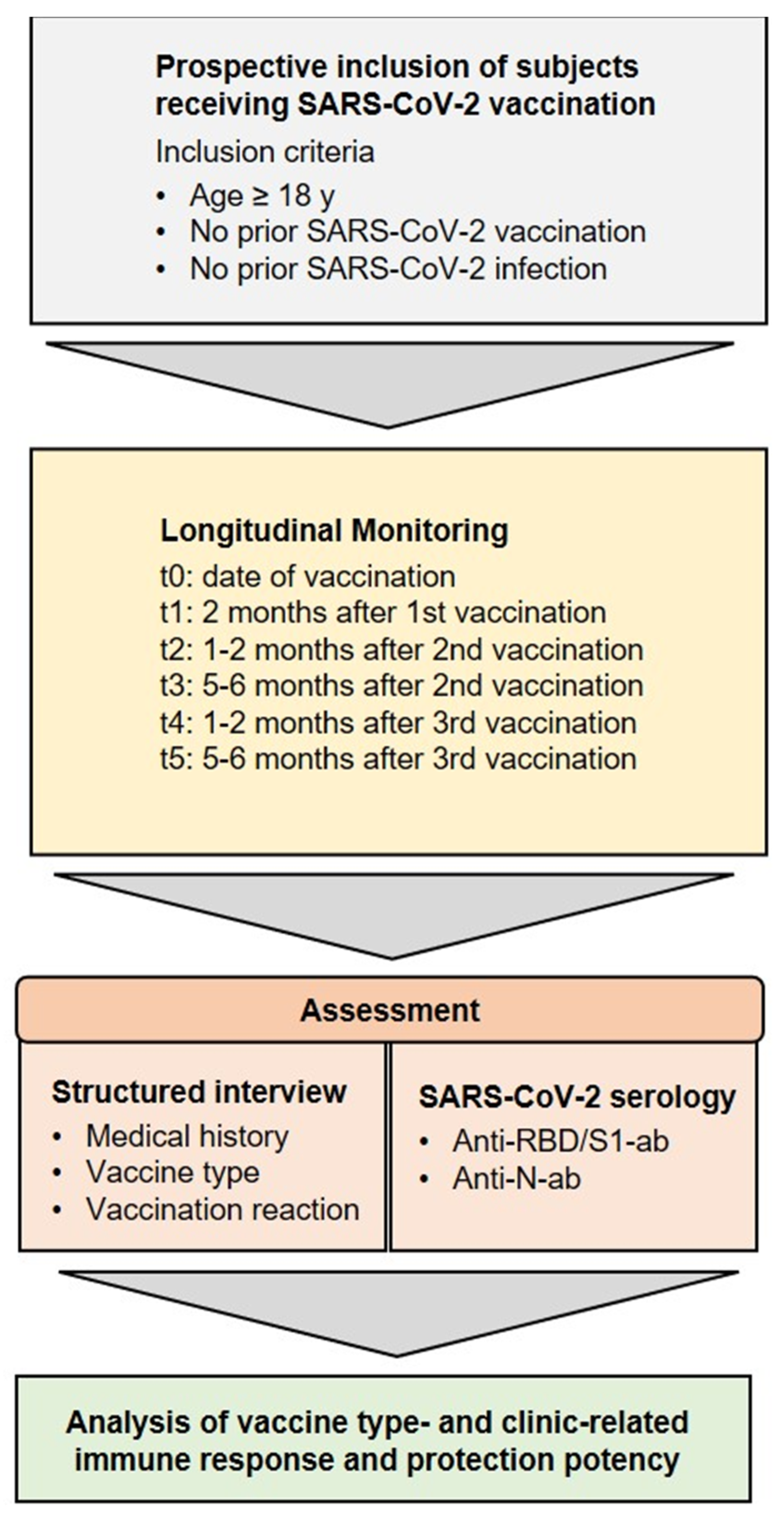
2. Materials and Methods
2.1. Participant Recruitment and Sample Collection
2.2. Anti-SARS-CoV-2 Antibody Detection
2.3. General Data Analysis
3. Results
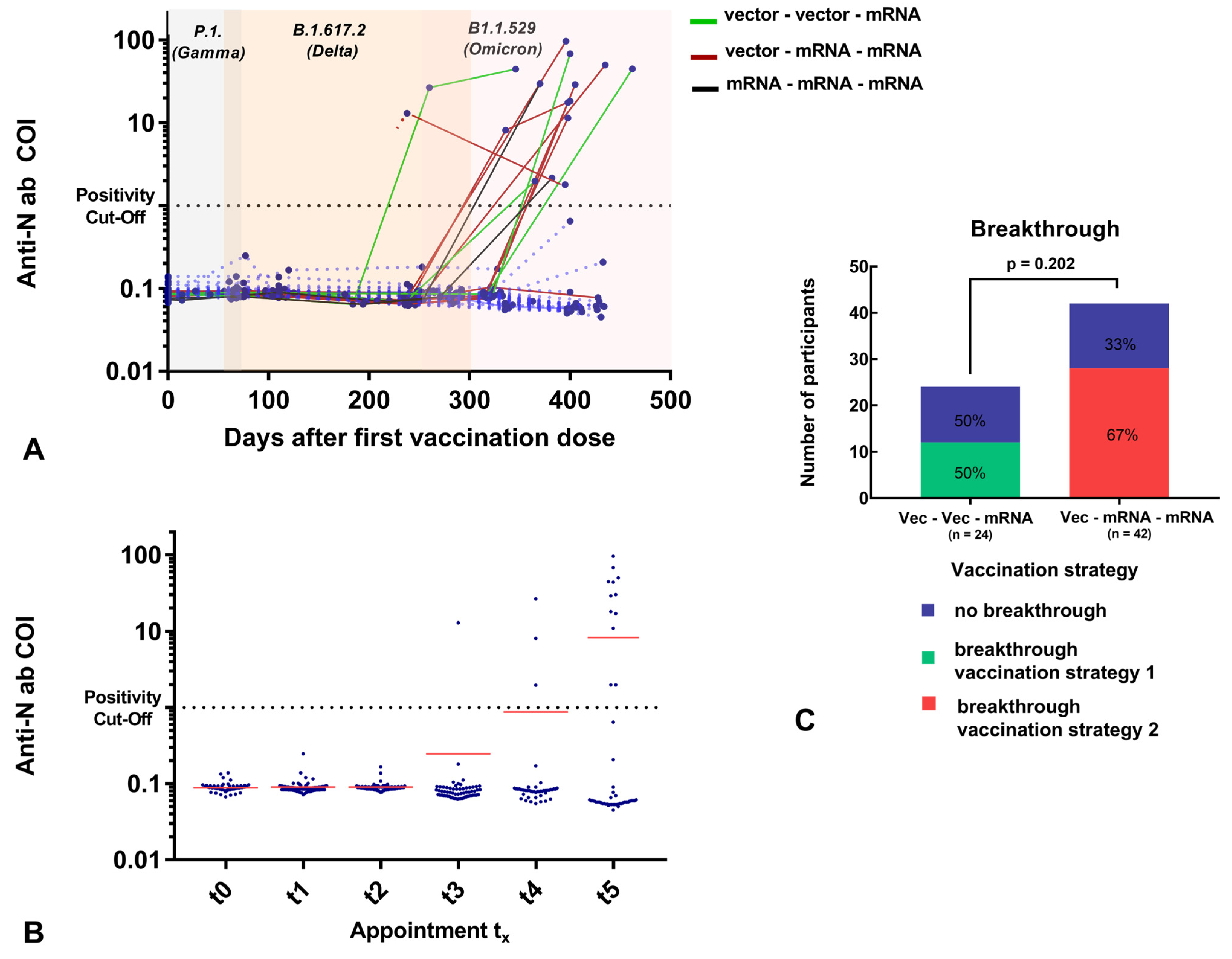
3.1. Demographics and Vaccination Strategies
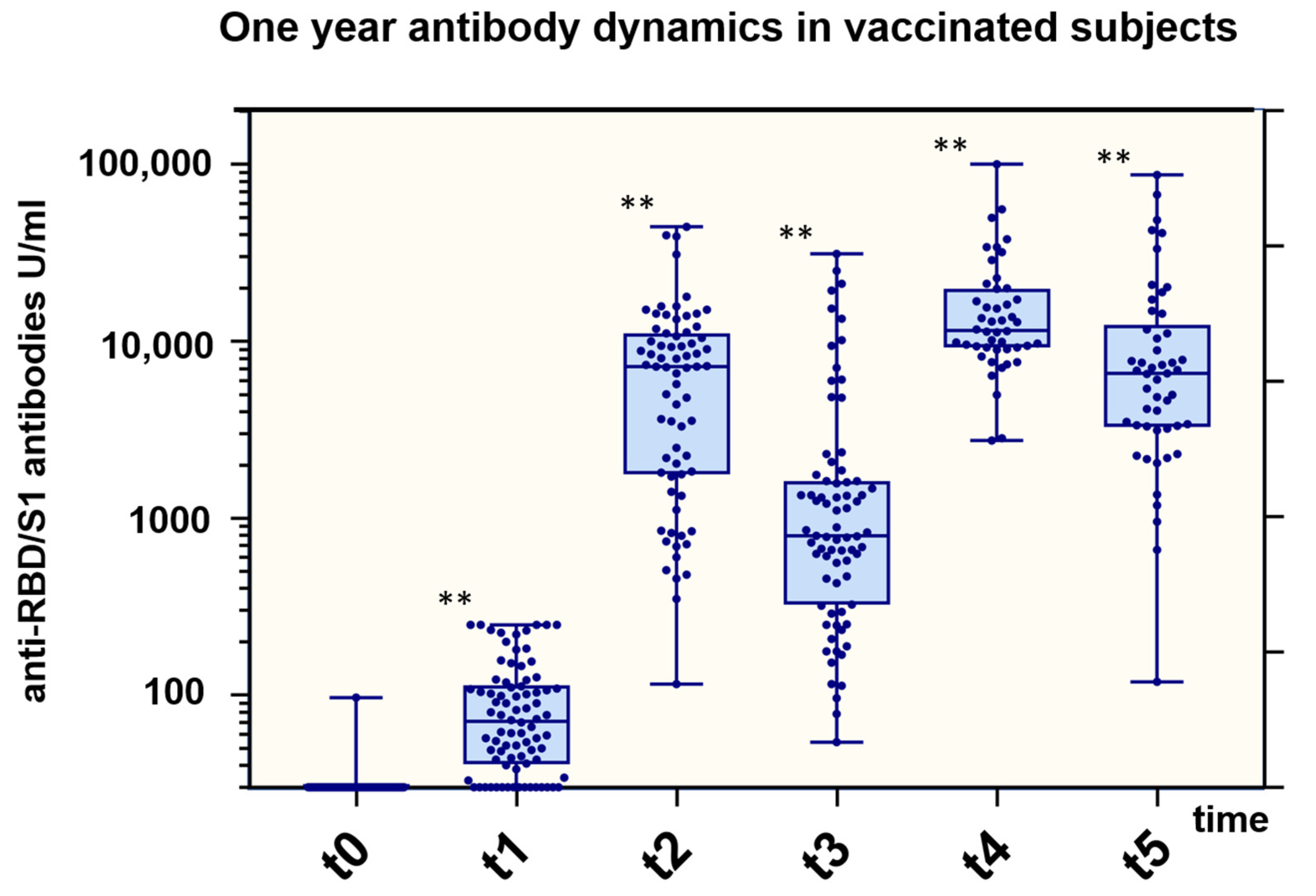
3.2. Overall Antibody Trends
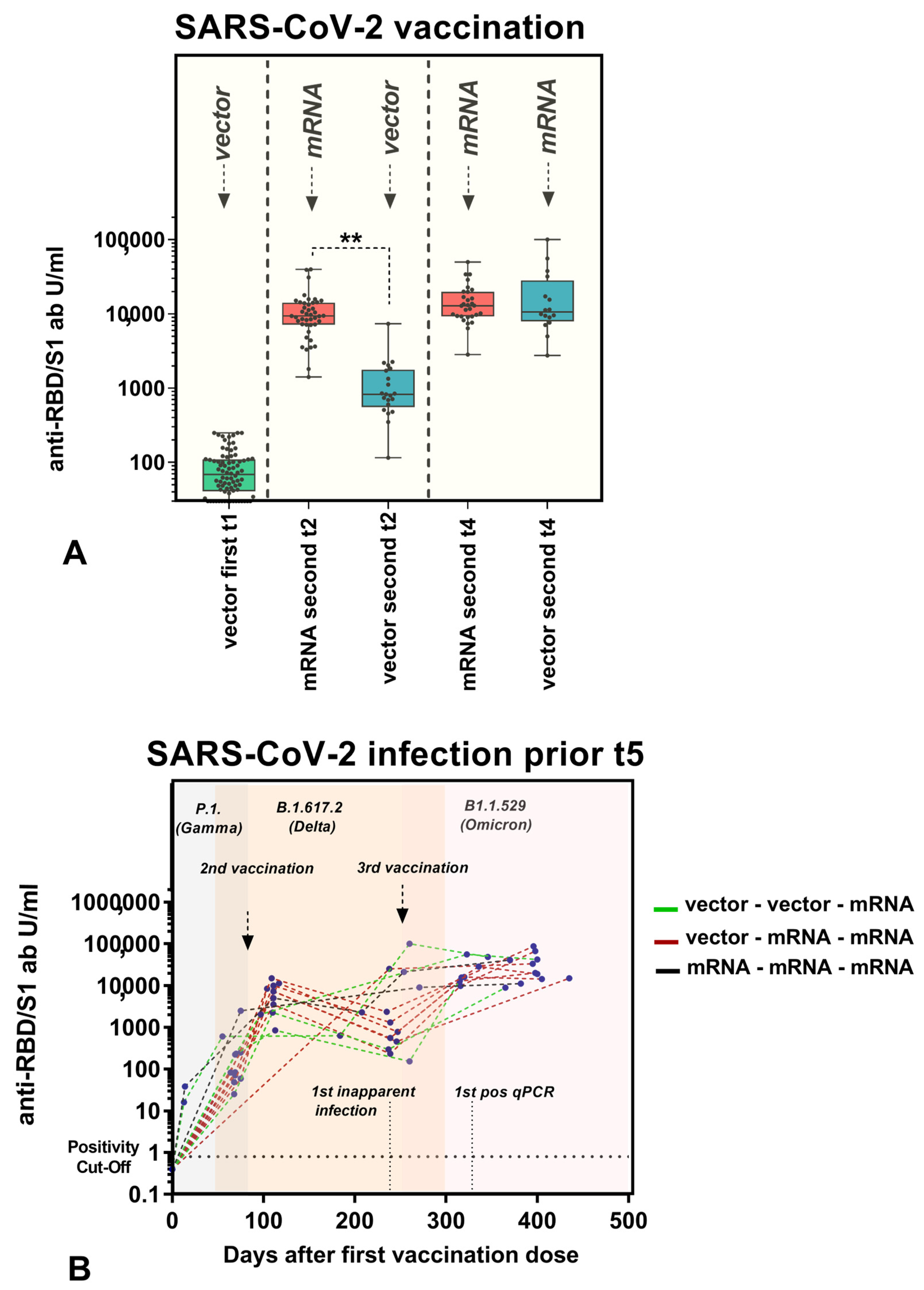
3.3. Anti-RBD/S1 Antibody Dynamics Correlated with Vaccination Strategy
3.4. Anti-RBD/S1 Antibody Dynamics Correlated with Vaccination Reaction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ab | Antibody |
| BMI | Body mass index |
| CE | Communauté Européenne |
| COI | Cut-off index |
| COVID-19 | Coronavirus disease 2019 |
| ECLIA | Electrochemiluminescence immunoassay |
| FDA | Food and Drug Administration |
| Ig | Immunoglobulin |
| ISO | International Organization for Standardization |
| qPCR | Quantitative polymerase chain reaction |
| RBD | Receptor-binding domain |
| REDCap | Research Electronic Data Capture |
| mRNA | Messenger ribonucleic acid |
| S | Spike protein |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
References
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, S.; Ou, J.; Zhang, J.; Lan, W.; Guan, W.; Wu, X.; Yan, Y.; Zhao, W.; Wu, J.; et al. COVID-19: Coronavirus Vaccine Development Updates. Front. Immunol. 2020, 11, 602256. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, A.M.; Petrover, D.R.; McMahon, T.M.; Libman, R.B.; Giliberto, L.; Clouston, S.A.P.; Castillo, M.; Kirsch, C. Retrospective review COVID-19 vaccine induced thrombotic thrombocytopenia and cerebral venous thrombosis-what can we learn from the immune response. Clin. Imaging 2022, 90, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.P.; Jonsdottir, H.R.; Brigger, D.; Damonti, L.; Suter-Riniker, F.; Endrich, O.; Froehlich, T.K.; Fiedler, M.; Largiadèr, C.R.; Marschall, J.; et al. Serological testing for SARS-CoV-2 antibodies in clinical practice: A comparative diagnostic accuracy study. Allergy 2022, 77, 2090–2103. [Google Scholar] [CrossRef]
- Fernandes-Matano, L.; Salas-Lais, A.G.; Grajales-Muñiz, C.; Hernández-Ávila, M.; Garfias-Becerra, Y.O.; Rodríguez-Sepúlveda, M.C.; Segura-Sánchez, C.; Montes-Herrera, D.; Mendoza-Sánchez, D.; Angeles-Martínez, J.; et al. Longevity and Neutralizing Capacity of IgG Antibodies against SARS-CoV-2 Generated by the Application of BNT162b2, AZD1222, Convidecia, Sputnik V, and CoronaVac Vaccines: A Cohort Study in the Mexican Population. Microbiol. Spectr. 2023, 11, e02376-22. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Torriani, G.; Yerly, S.; Mazza, L.; Calame, A.; Arm-Vernez, I.; Zimmer, G.; Agoritsas, T.; Stirnemann, J.; Spechbach, H.; et al. Validation of a commercially available SARS-CoV-2 serological Immunoassay. Clin. Microbiol. Infect. 2020, 26, 1386–1394. [Google Scholar] [CrossRef]
- Bonelli, F.; Sarasini, A.; Zierold, C.; Calleri, M.; Bonetti, A.; Vismara, C.; Blocki, F.A.; Pallavicini, L.; Chinali, A.; Campisi, D.; et al. Clinical and Analytical Performance of an Automated Serological Test That Identifies S1/S2-Neutralizing IgG in COVID-19 Patients Semiquantitatively. J. Clin. Microbiol. 2020, 58, e01224-20. [Google Scholar] [CrossRef]
- Glöckner, S.; Hornung, F.; Baier, M.; Weis, S.; Pletz, M.W.; Deinhardt-Emmer, S.; Löffler, B.; the CoNAN Study Group. Robust Neutralizing Antibody Levels Detected after Either SARS-CoV-2 Vaccination or One Year after Infection. Viruses 2021, 13, 2003. [Google Scholar] [CrossRef]
- Jara, A.; Undurraga, E.A.; Zubizarreta, J.R.; González, C.; Pizarro, A.; Acevedo, J.; Leo, K.; Paredes, F.; Bralic, T.; Vergara, V.; et al. Effectiveness of homologous and heterologous booster doses for an inactivated SARS-CoV-2 vaccine: A large-scale prospective cohort study. Lancet Glob. Health 2022, 10, e798–e806. [Google Scholar] [CrossRef]
- Wanlapakorn, N.; Suntronwong, N.; Phowatthanasathian, H.; Yorsaeng, R.; Vichaiwattana, P.; Thongmee, T.; Auphimai, C.; Srimuan, D.; Thatsanatorn, T.; Assawakosri, S.; et al. Safety and immunogenicity of heterologous and homologous inactivated and adenoviral-vectored COVID-19 vaccine regimens in healthy adults: A prospective cohort study. Hum. Vaccines Immunother. 2022, 18, 2029111. [Google Scholar] [CrossRef]
- Gerhards, C.; Thiaucourt, M.; Kittel, M.; Becker, C.; Ast, V.; Hetjens, M.; Neumaier, M.; Haselmann, V. Longitudinal assessment of anti-SARS-CoV-2 antibody dynamics and clinical features following convalescence from a COVID-19 infection. Int. J. Infect. Dis. 2021, 107, 221–227. [Google Scholar] [CrossRef] [PubMed]
- R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org (accessed on 1 October 2021).
- Mason, T.F.D.; Whitston, M.; Hodgson, J.; Watkinson, R.E.; Lau, Y.-S.; Abdulrazeg, O.; Sutton, M. Effects of BNT162b2 mRNA vaccine on COVID-19 infection and hospitalisation amongst older people: Matched case control study for England. BMC Med. 2021, 19, 275. [Google Scholar] [CrossRef] [PubMed]
- Mostaghimi, D.; Valdez, C.N.; Larson, H.T.; Kalinich, C.C.; Iwasaki, A. Prevention of host-to-host transmission by SARS-CoV-2 vaccines. Lancet Infect. Dis. 2022, 22, e52–e58. [Google Scholar] [CrossRef] [PubMed]
- Fadul, A.; ELMustafa, A.; Abdelmahmuod, E.; Abdulgayoom, M.; Ali, E.; Al-warqi, A.; Al-yahary, H. COVID-19 Vaccine-Induced Cerebral Sinus Thrombosis: Coincidence vs. Cause? Cureus 2022, 14, e26436. [Google Scholar] [CrossRef]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef]
- Spencer, A.J.; McKay, P.F.; Belij-Rammerstorfer, S.; Ulaszewska, M.; Bissett, C.D.; Hu, K.; Samnuan, K.; Blakney, A.K.; Wright, D.; Sharpe, H.R.; et al. Heterologous vaccination regimens with self-amplifying RNA and adenoviral COVID vaccines induce robust immune responses in mice. Nat. Commun. 2021, 12, 2893. [Google Scholar] [CrossRef]
- Sritipsukho, P.; Khawcharoenporn, T.; Siribumrungwong, B.; Damronglerd, P.; Suwantarat, N.; Satdhabudha, A.; Chaiyakulsil, C.; Sinlapamongkolkul, P.; Tangsathapornpong, A.; Bunjoungmanee, P.; et al. Comparing real-life effectiveness of various COVID-19 vaccine regimens during the delta variant-dominant pandemic: A test-negative case-control study. Emerg. Microbes Infect. 2022, 11, 585–592. [Google Scholar] [CrossRef]
- Larkin, H.D. New Data on Heterologous COVID-19 Vaccine Combinations. JAMA 2022, 328, 916. [Google Scholar] [CrossRef]
- Kim, D.-I.; Lee, S.J.; Park, S.; Kim, P.; Lee, S.M.; Lee, N.; Shum, D.; Kim, D.H.; Kim, E.H. Immunogenicity and Durability of Antibody Responses to Homologous and Heterologous Vaccinations with BNT162b2 and ChAdOx1 Vaccines for COVID-19. Vaccines 2022, 10, 1864. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Salles, T.; Prieto-Alhambra, D. Heterologous vaccine regimens against COVID-19. Lancet 2021, 398, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Serebro, D. COVID-19 Vaccine Financing and Purchasing in Africa: Wherefrom the Money? 2021. Available online: https://www.cabri-sbo.org/en/blog/2021/covid-19-vaccine-financing-and-purchasing-in-africa-wherefrom-the-money (accessed on 14 February 2023).
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Homologous and Heterologous COVID-19 Booster Vaccinations. N. Engl. J. Med. 2022, 386, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Wanlapakorn, N.; Suntronwong, N.; Phowatthanasathian, H.; Yorsaeng, R.; Thongmee, T.; Vichaiwattana, P.; Auphimai, C.; Wongsrisang, L.; Klinfueng, S.; Sudhinaraset, N.; et al. Immunogenicity of heterologous inactivated and adenoviral-vectored COVID-19 vaccine: Real-world data. Vaccine 2022, 40, 3203–3209. [Google Scholar] [CrossRef]
- Adjobimey, T.; Meyer, J.; Sollberg, L.; Bawolt, M.; Berens, C.; Kovačević, P.; Trudić, A.; Parcina, M.; Hoerauf, A. Comparison of IgA, IgG, and Neutralizing Antibody Responses Following Immunization with Moderna, BioNTech, AstraZeneca, Sputnik-V, Johnson and Johnson, and Sinopharm’s COVID-19 Vaccines. Front. Immunol. 2022, 13, 917905. [Google Scholar] [CrossRef] [PubMed]
- Coggins, S.A.; Laing, E.D.; Olsen, C.H.; Goguet, E.; Moser, M.; Jackson-Thompson, B.M.; Samuels, E.C.; Pollett, S.D.; Tribble, D.R.; Davies, J.; et al. Adverse Effects and Antibody Titers in Response to the BNT162b2 mRNA COVID-19 Vaccine in a Prospective Study of Healthcare Workers. Open Forum Infect. Dis. 2022, 9, ofab575. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Saez, F.; Peiró, S.; Cuenca, L.; Vanaclocha, H.; Limón, R.; Salas, D.; Burgos, J.S.; Sánchez-Payá, J.; Meneu, R.; Díez, J.; et al. Side effects during the week after first dose vaccination with four COVID-19 vaccines. Results of the ProVaVac Survey Study with 13,837 people in Spain. Vaccine 2022, 40, 5942–5949. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Heterologous SARS-CoV-2 Booster Vaccinations—Preliminary Report. medRxiv 2021. [Google Scholar] [CrossRef]
- Sugiyama, K.; Suzuki, C.; Aoyama, M.; Toyota, N.; Nakagawa, N.; Shozu, M.; Nakai, K.; Iwano, K. Long-term observation of antibody titers against SARS-CoV-2 following vaccination. Public Health Pract. 2022, 4, 100297. [Google Scholar] [CrossRef]
- Irsara, C.; Egger, A.E.; Prokop, W.; Nairz, M.; Loacker, L.; Sahanic, S.; Pizzini, A.; Sonnweber, T.; Holzer, B.; Mayer, W.; et al. Clinical validation of the Siemens quantitative SARS-CoV-2 spike IgG assay (sCOVG) reveals improved sensitivity and a good correlation with virus neutralization titers. Clin. Chem. Lab. Med. CCLM 2021, 59, 1453–1462. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Zhong, W.; Roberts, A.D.; Woodland, D.L. Antibody-Independent Antiviral Function of Memory CD4+ T Cells In Vivo Requires Regulatory Signals from CD8+ Effector T Cells. J. Immunol. 2001, 167, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Alshukairi, A.N.; Baharoon, S.A.; Ahmed, W.A.; Bokhari, A.A.; Nehdi, A.M.; Layqah, L.A.; Alghamdi, M.G.; Al Gethamy, M.M.; Dada, A.M.; et al. Recovery from the Middle East respiratory syndrome is associated with antibody and T cell responses. Sci. Immunol. 2017, 2, eaan5393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meschi, S.; Matusali, G.; Colavita, F.; Lapa, D.; Bordi, L.; Puro, V.; Leoni, B.D.; Galli, C.; Capobianchi, M.R.; Castilletti, C. Predicting the protective humoral response to a SARS-CoV-2 mRNA vaccine. Clin. Chem. Lab. Med. CCLM 2021, 59, 2010–2018. [Google Scholar] [CrossRef]
- Kittel, M.; Eichner, R.; Aida, S.; Bode, A.; Ast, V.; Kessler, A.; Neumaier, M.; Wölfel, R.; Haselmann, V. Results of a European-Wide External Quality Assessment (EQA) Scheme for Serological Detection of Anti-SARS-CoV-2 (CoVimm)—Pitfalls of Routine Application. Viruses 2022, 14, 1662. [Google Scholar] [CrossRef]
- Roche Diagnostics Deutschland GmbH. ElecsysT Anti-SARS-CoV-2 S. Available online: https://assets.cwp.roche.com/f/94122/x/37b1d9536f/factsheet-elecsys-anti-sars-cov-2-s_v3.pdf (accessed on 21 February 2023).
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Methot, N.; Yu, E.D.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; Wang, E.; et al. Impact of SARS-CoV-2 variants on the total CD4+ and CD8+ T cell reactivity in infected or vaccinated individuals. Cell Rep. Med. 2021, 2, 100355. [Google Scholar] [CrossRef] [PubMed]
- Naranbhai, V.; Nathan, A.; Kaseke, C.; Berrios, C.; Khatri, A.; Choi, S.; Getz, M.A.; Tano-Menka, R.; Ofoman, O.; Gayton, A.; et al. T cell reactivity to the SARS-CoV-2 Omicron variant is preserved in most but not all prior infected and vaccinated individuals. medRxiv 2022. [Google Scholar] [CrossRef]
- Tartof, S.Y.; Slezak, J.M.; Fischer, H.; Hong, V.; Ackerson, B.K.; Ranasinghe, O.N.; Frankland, T.B.; Ogun, O.A.; Zamparo, J.M.; Gray, S.; et al. Effectiveness of mRNA BNT162b2 COVID-19 vaccine up to 6 months in a large integrated health system in the USA: A retrospective cohort study. Lancet 2021, 398, 1407–1416. [Google Scholar] [CrossRef]
- Takita, M.; Yoshida, T.; Tsuchida, T.; Nakagama, Y.; Kido, Y.; Suzuki, S.; Imamura, M.; Kawahata, K.; Shimizu, G.; Yoshida, H.; et al. Low SARS-CoV-2 antibody titers may be associated with poor clinical outcomes for patients with severe COVID-19. Sci. Rep. 2022, 12, 9147. [Google Scholar] [CrossRef]
- Amjadi, M.F.; O’Connell, S.E.; Armbrust, T.; Mergaert, A.M.; Narpala, S.R.; Halfmann, P.J.; Bashar, S.J.; Glover, C.R.; Heffron, A.S.; Taylor, A.; et al. Specific COVID-19 Symptoms Correlate with High Antibody Levels against SARS-CoV-2. ImmunoHorizons 2021, 5, 466–476. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Overall |
|---|---|
| n = 103 | |
| Demographics | |
| Sex F/M (%) | 78/25 (76.7/24.3) |
| Age (mean (SD)) | 39.64 (14.81) |
| BMI (mean (SD)) | 25.32 (5.10) |
| Smoking | 20/81 (24.7%) |
| 1–5 cigarettes/day | 4 (4.9%) |
| 6–10 cigarettes/day | 6 (7.4%) |
| 11–20 cigarettes/day | 7 (8.6%) |
| 31–40 cigarettes/day | 3 (3.7%) |
| Pre-existing disease | |
| Pulmonary disease | 10/81 (12.3%) |
| Vascular disease | 6/81 (7.4%) |
| Autoimmune disease | 3/81 (3.7%) |
| Cancer (in remission) | 2/81 (2.5%) |
| Diabetes | 1/81 (1.2%) |
| Other disease | 17/81 (21.0%) |
| Hypothyroidism | 6/81 (7.4%) |
| Allergy | 37/81 (45.7%) |
| Medication | |
| Any medication | 33/81 (40.7%) |
| Immunosuppression | 0/81 (0%) |
| Antiallergic medication | 9/81 (11.1%) |
| Blood type | |
| Unknown | 31 (38.3%) |
| 0+ | 17 (21.0%) |
| A+ | 15 (18.5%) |
| B+ | 3 (3.7%) |
| AB+ | 4 (4.9%) |
| 0- | 4 (4.9%) |
| A- | 6 (7.4%) |
| AB- | 1 (1.2%) |
| SARS-CoV-2 anamnesis | |
| Contact (1st vaccination) | 13/82 (15.9%) |
| Previous infection | 2/103 (1.9%) |
| SARS-CoV-2 vaccination | |
| First dose | |
| ChAdOx1 nCoV-19 (AstraZeneca) | 85 (94.4%) |
| BNT162b2 (Pfizer-BioNTech) | 4 (4.4%) |
| mRNA-1273 (Moderna) | 1 (1.1%) |
| Second dose | |
| ChAdOx1 nCoV-19 (AstraZeneca) | 27 (32.1%) |
| BNT162b2 (Pfizer-BioNTech) | 55 (65.5%) |
| mRNA-1273 (Moderna) | 2 (2.4%) |
| Third dose | |
| ChAdOx1 nCoV-19 (AstraZeneca) | 0 (0%) |
| BNT162b2 (Pfizer-BioNTech) | 59 (84.5%) |
| mRNA-1273 (Moderna) | 11 (15.5%) |
| Vaccination strategy | |
| First and second dose | |
| Homologous (AstraZeneca) | 26 (31.0%) |
| Heterologous (AstraZeneca/ BioNTech) | 53 (63.1%) |
| Homologous (BioNTech) | 4 (4.8%) |
| Homologous (Moderna) | 1 (1.2%) |
| Second and third dose | |
| Homologous (mRNA vaccines) | 47 (66.2%) |
| Heterologous (mRNA/vector) | 24 (33.8%) |
| Variable | Overall |
|---|---|
| t0 anti-N abs (median (IQR)) | 0.09 (0.09, 0.09) |
| t1 anti-N abs (mean (SD)) | 0.94 (7.70) |
| t2 anti-N abs (median (IQR)) | 0.09 (0.09, 0.09) |
| t3 anti-N abs (median (IQR)) | 0.07 (0.07, 0.09) |
| t4 anti-N abs (median (IQR)) | 0.08 (0.08, 0.09) |
| t5 anti-N abs (median (IQR)) | 0.06 (0.06, 1.50) |
| t0 anti-RBD/S1 (median (IQR)) | 0.40 (0.40, 0.40) |
| t1 anti-RBD/S1 (median (IQR)) | 71.43 (41.76, 111.68) |
| t2 anti-RBD/S1 (median (IQR)) | 7201.00 (1793.00, 10,839.50) |
| t3 anti-RBD/S1 (median (IQR)) | 791.70 (376.95, 1619.00) |
| t4 anti-RBD/S1 (median (IQR)) | 11,490.00 (9239.00, 19,079.50) |
| t5 anti-RBD/S1 (median (IQR)) | 6557.00 (3304.25, 11,506.25) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerhards, C.; Thiaucourt, M.; Hetjens, M.; Haselmann, V.; Neumaier, M.; Kittel, M. Heterologous Vector—mRNA Based SARS-CoV-2 Vaccination Strategy Appears Superior to a Homologous Vector—Based Vaccination Scheme in German Healthcare Workers Regarding Humoral SARS-CoV-2 Response Indicating a High Boosting Effect by mRNA Vaccines. Vaccines 2023, 11, 701. https://doi.org/10.3390/vaccines11030701
Gerhards C, Thiaucourt M, Hetjens M, Haselmann V, Neumaier M, Kittel M. Heterologous Vector—mRNA Based SARS-CoV-2 Vaccination Strategy Appears Superior to a Homologous Vector—Based Vaccination Scheme in German Healthcare Workers Regarding Humoral SARS-CoV-2 Response Indicating a High Boosting Effect by mRNA Vaccines. Vaccines. 2023; 11(3):701. https://doi.org/10.3390/vaccines11030701
Chicago/Turabian StyleGerhards, Catharina, Margot Thiaucourt, Michael Hetjens, Verena Haselmann, Michael Neumaier, and Maximilian Kittel. 2023. "Heterologous Vector—mRNA Based SARS-CoV-2 Vaccination Strategy Appears Superior to a Homologous Vector—Based Vaccination Scheme in German Healthcare Workers Regarding Humoral SARS-CoV-2 Response Indicating a High Boosting Effect by mRNA Vaccines" Vaccines 11, no. 3: 701. https://doi.org/10.3390/vaccines11030701