
Genetic Diversity and Evolutionary Kinetics of Influenza A Virus H3N2 Subtypes Circulating in Riyadh, Saudi Arabia
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Detection, Typing, and Sequencing of IAV
2.2. Sequence Data Analysis and Phylogenetic Analysis
2.3. Statistical Analysis
3. Results
3.1. Detection of IAV and Subtyping
3.2. Sequence Analysis of HA and NA Genes of H3N2
3.2.1. Nucleotide and AA Sequence Analysis of HA Gene of H3N2 Subtype Study Strains
3.2.2. Sequence Analysis of NA Gene of H3N2 Strains
3.3. N- and O-Glycosylation Site Analysis in the AA Sequence
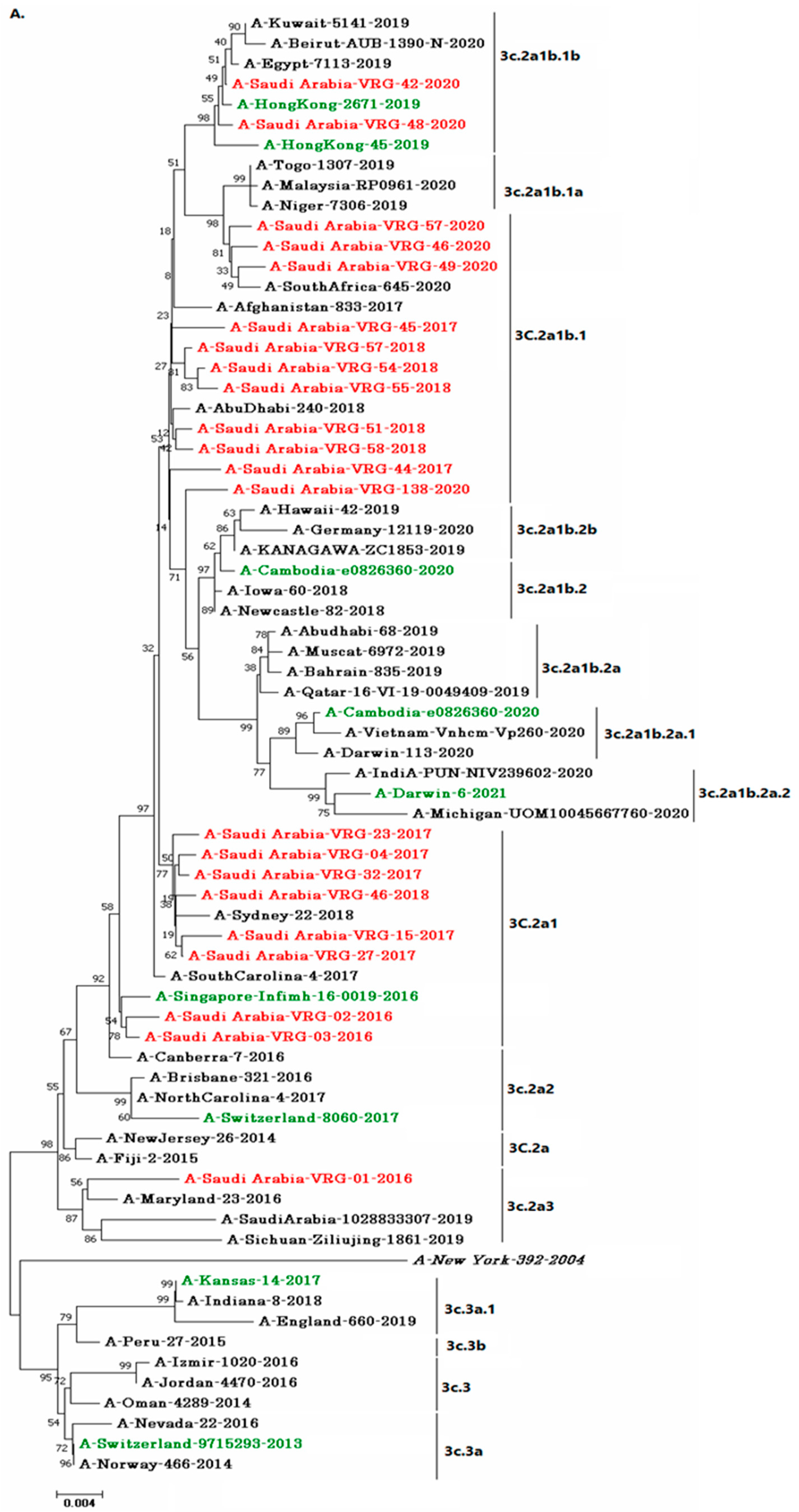
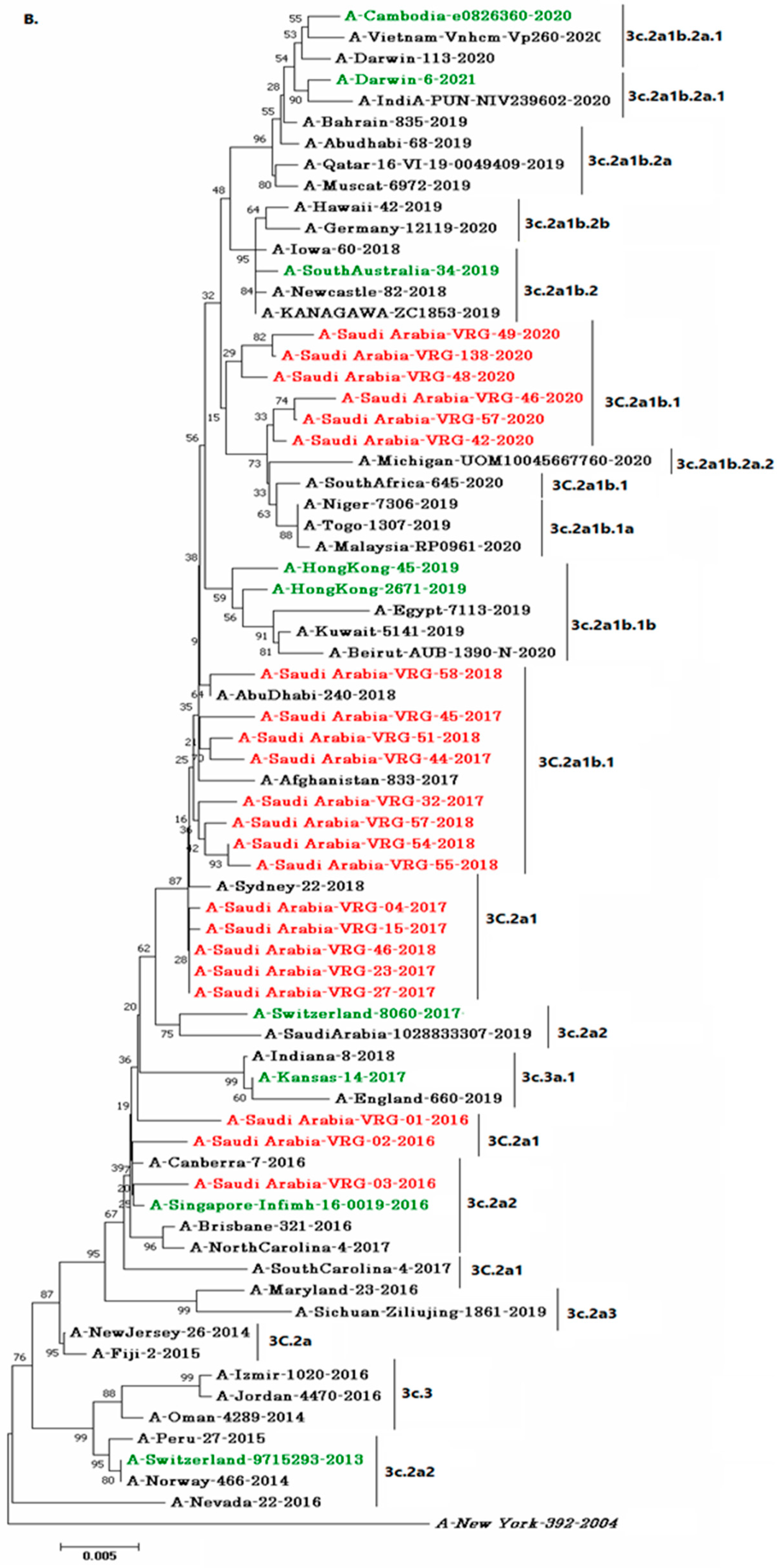
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- de Zwart, A.; Riezebos-Brilman, A.; Lunter, G.; Vonk, J.; Glanville, A.R.; Gottlieb, J.; Permpalung, N.; Kerstjens, H.; Alffenaar, J.W.; Verschuuren, E. Respiratory syncytial virus, human metapneumovirus, and parainfluenza virus infections in lung transplant recipients: A systematic review of outcomes and treatment strategies. Clin. Infect. Dis. 2022, 74, 2252–2260. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, K. The role of molecular diagnosis in acute respiratory tract infection. Respir. Investig. 2019, 57, 511. [Google Scholar] [CrossRef] [PubMed]
- Hamrin, J. Viral Respiratory Tract Infections in Children. Ph.D. Thesis, Karolinska Institutet, Solna, Sweden, 2021. [Google Scholar]
- Govers, C.; Calder, P.C.; Savelkoul, H.F.J.; Albers, R.; van Neerven, R.J.J. Ingestion, Immunity, and Infection: Nutrition and Viral Respiratory Tract Infections. Front. Immunol. 2022, 13, 632. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, C.S.; Allen, J.L.Y.; Gkrania-Klotsas, E. Influenza: Epidemiology and hospital management. Medicine 2021, 49, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Noda, T. Native Morphology of Influenza Virions. Front. Microbiol. 2012, 2, 269. [Google Scholar] [CrossRef] [Green Version]
- To, K.; Ng, K.H.; Que, T.L.; Chan, J.; Tsang, K.-Y.; Tsang, A.K.; Chen, H.; Yuen, K.-Y. Avian influenza A H5N1 virus: A continuous threat to humans. Emerg. Microbes Infect. 2012, 1, 1–12. [Google Scholar] [CrossRef]
- Li, Y.-T.; Linster, M.; Mendenhall, I.H.; Su, Y.C.F.; Smith, G.J.D. Avian influenza viruses in humans: Lessons from past outbreaks. Br. Med Bull. 2019, 132, 81–95. [Google Scholar] [CrossRef]
- Mineev, K.; Lyukmanova, E.N.; Krabben, L.; Serebryakova, M.; Shulepko, M.; Arseniev, A.S.; Kordyukova, L.V.; Veit, M. Structural investigation of influenza virus hemagglutinin membrane-anchoring peptide. Protein Eng. Des. Sel. 2013, 26, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Bakar, U.A.; Amrani, L.; Kamarulzaman, F.; Karsani, S.A.; Hassandarvish, P.; Khairat, J.E. Avian Influenza Virus Tropism in Humans. Preprints 2023, 2023020081. [Google Scholar] [CrossRef]
- Merced-Morales, A.; Daly, P.; Abd Elal, A.I.; Ajayi, N.; Annan, E.; Budd, A.; Barnes, J.; Colon, A.; Cummings, C.N.; Iuliano, A.D.; et al. Influenza Activity and Composition of the 2022–23 Influenza Vaccine—United States, 2021—22 Season. Morb. Mortal. Wkly. Rep. 2022, 71, 2019–2020. [Google Scholar] [CrossRef]
- Benkouiten, S.; Al-Tawfiq, J.A.; Memish, Z.A.; Albarrak, A.; Gautret, P. Clinical respiratory infections and pneumonia during the Hajj pilgrimage: A systematic review. Travel Med. Infect. Dis. 2018, 28, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. The beginning and ending of a respiratory viral pandemic-lessons from the Spanish flu. Microb. Biotechnol. 2022, 15, 1301–1317. [Google Scholar] [CrossRef]
- World Health Organisation. WHO Information for the Molecular Detection of Influenza Viruses; World Health Organisation: Geneva, Switzerland, 2017; Available online: http://www.who.int/influenza/gisrs_laboratory/molecular_diagnosis/en (accessed on 8 February 2022).
- Agustiningsih, A.; Trimarsanto, H.; Setiawaty, V.; Artika, I.M.; Muljono, D.H. Primer development to obtain complete coding sequence of HA and NA genes of influenza A/H3N2 virus. BMC Res. Notes 2016, 9, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-Glycosylation Sites in Human Proteins. 2004. Available online: https://www.hsls.pitt.edu/obrc/index.php?page=URL1043959999 (accessed on 30 January 2022).
- Julenius, K.; Mølgaard, A.; Gupta, R.; Brunak, S. Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites. Glycobiology 2004, 15, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Regional Office for Europe Guidance for Influenza Surveillance in Humans. 2009. Available online: https://apps.who.int/iris/handle/10665/107265 (accessed on 8 February 2022).
- Beauté, J.; Zucs, P.; Korsun, N.; Bragstad, K.; Enouf, V.; Kossyvakis, A.; Griškevičius, A.; Olinger, C.M.; Meijer, A.; Guiomar, R.; et al. Age-specific differences in influenza virus type and subtype distribution in the 2012/2013 season in 12 European countries. Epidemiol. Infect. 2015, 143, 2950–2958. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.; Ahmed, A.; Zeitounie, R.; Khandekar, R. Impact of influenza vaccine in reduction of incidence and severity of influenza-like illness. East. Mediterr. Health J. 2021, 27, 884–891. [Google Scholar] [CrossRef]
- Elhakim, M.; Rasooly, M.H.; Fahim, M.; Ali, S.S.; Haddad, N.; Cherkaoui, I.; Hjaija, D.; Nadeem, S.; Assiri, A.; Aljifri, A.; et al. Epidemiology of severe cases of influenza and other acute respiratory infections in the Eastern Mediterranean Region, July 2016 to June 2018. J. Infect. Public Health 2019, 13, 423–429. [Google Scholar] [CrossRef]
- Althaqafi, A.; Farahat, F.; Alsaedi, A.; Alshamrani, M.; Alsaeed, M.S.; AlhajHussein, B.; El-Kafrawy, S.A.; Azhar, E.I. Molecular Detection of Influenza A and B Viruses in Four Consecutive Influenza Seasons 2015–16 to 2018–19 in a Tertiary Center in Western Saudi Arabia. J. Epidemiology Glob. Health 2021, 11, 208–215. [Google Scholar] [CrossRef]
- Cobbin, J.C.A.; Alfelali, M.; Barasheed, O.; Taylor, J.; Dwyer, D.E.; Kok, J.; Booy, R.; Holmes, E.C.; Rashid, H. Multiple Sources of Genetic Diversity of Influenza A Viruses during the Hajj. J. Virol. 2017, 91, e00096-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, M.R.; Badra, R.; El Rifay, A.S.; Kandeil, A.; Kamel, M.N.; Shama, N.M.A.; El-Shesheny, R.; Barakat, A.B.; Ali, M.A.; Kayali, G. Incidence and seroprevalence of seasonal influenza a viruses in Egypt: Results of a community-based cohort study. Influenza Other Respir. Viruses 2022, 16, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.R.; Simonsen, L.; Edelson, P.J.; Morse, S.S. Epidemiological evidence of an early wave of the 1918 influenza pandemic in New York City. Proc. Natl. Acad. Sci. USA 2005, 102, 11059–11063. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.P.; Ponomarenko, A. System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality. Infect. Dis. Rep. 2022, 14, 287–309. [Google Scholar] [CrossRef]
- Tuckerman, J.; Harper, K.; Sullivan, T.R.; Cuthbert, A.R.; Fereday, J.; Couper, J.; Smith, N.; Tai, A.; Kelly, A.; Couper, R.; et al. Short Message Service Reminder Nudge for Parents and Influenza Vaccination Uptake in Children and Adolescents with Special Risk Medical Conditions: The Flutext-4U Randomized Clinical Trial. JAMA Pediatr. 2023. [Google Scholar] [CrossRef] [PubMed]
- Grohskopf, L.A.; Sokolow, L.Z.; Broder, K.R.; Walter, E.B.; Fry, A.M.; Jernigan, D.B. Prevention and control of seasonal influenza with vaccines: Recommendations of the Advisory Committee on Immunization Practices—United States, 2018–2019 influenza season. MMWR Recomm. Rep. 2018, 67, 1. [Google Scholar] [CrossRef]
- Aoun, T.; Borrow, R.; Arkwright, P.D. Immunogenicity and safety of seasonal influenza vaccines in children under 3 years of age. Expert Rev. Vaccines 2023, 22. [Google Scholar] [CrossRef]
- Barr, I.G.; Jelley, L.L. The coming era of quadrivalent human influenza vaccines: Who will benefit? Drugs 2012, 72, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wharton, S.A.; Whittaker, L.; Dai, M.; Ermetal, B.; Lo, J.; Pontoriero, A.; Baumeister, E.; Daniels, R.S.; McCauley, J.W. The characteristics and antigenic properties of recently emerged subclade 3C.3a and 3C.2a human influenza A(H3N2) viruses passaged in MDCK cells. Influenza Other Respir. Viruses 2017, 11, 263–274. [Google Scholar] [CrossRef]
- Melidou, A.; Gioula, G.; Exindari, M.; Ioannou, E.; Gkolfinopoulou, K.; Georgakopoulou, T.; Tsiodras, S.; Papa, A. Influenza A(H3N2) genetic variants in vaccinated patients in northern Greece. J. Clin. Virol. 2017, 94, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Kissling, E.; Pozo, F.; Buda, S.; Vilcu, A.-M.; Rizzo, C.; Gherasim, A.; Horváth, J.K.; Brytting, M.; Domegan, L.; Meijer, A.; et al. Effectiveness of influenza vaccine against influenza A in Europe in seasons of different A(H1N1)pdm09 and the same A(H3N2) vaccine components (2016–17 and 2017–18). Vaccine X 2019, 3, 100042. [Google Scholar] [CrossRef]
- Chambers, B.S.; Parkhouse, K.; Ross, T.; Alby, K.; Hensley, S.E. Identification of Hemagglutinin Residues Responsible for H3N2 Antigenic Drift during the 2014–2015 Influenza Season. Cell Rep. 2015, 12, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronski, D.M.; Sabaiduc, S.; Chambers, C.; Eshaghi, A.; Gubbay, J.B.; Krajden, M.; Drews, S.J.; Martineau, C.; De Serres, G.; Dickinson, J.; et al. Mutations acquired during cell culture isolation may affect antigenic characterisation of influenza A(H3N2) clade 3C.2a viruses. Eurosurveillance 2016, 21, 30112. [Google Scholar] [CrossRef]
- Kim, P.; Jang, Y.H.; Bin Kwon, S.; Lee, C.M.; Han, G.; Seong, B.L. Glycosylation of Hemagglutinin and Neuraminidase of Influenza A Virus as Signature for Ecological Spillover and Adaptation among Influenza Reservoirs. Viruses 2018, 10, 183. [Google Scholar] [CrossRef] [Green Version]
- Klenk, H.-D.; Wagner, R.; Heuer, D.; Wolff, T. Importance of hemagglutinin glycosylation for the biological functions of influenza virus. Virus Res. 2001, 82, 73–75. [Google Scholar] [CrossRef]
- Kumari, R.; Sharma, S.D.; Kumar, A.; Ende, Z.; Mishina, M.; Wang, Y.; Falls, Z.; Samudrala, R.; Pohl, J.; Knight, P.R.; et al. Antiviral Approaches against Influenza Virus. Clin. Microbiol. Rev. 2023, e00040-22. [Google Scholar] [CrossRef]
- Kawakami, C.; Yamayoshi, S.; Akimoto, M.; Nakamura, K.; Miura, H.; Fujisaki, S.; Pattinson, D.J.; Shimizu, K.; Ozawa, H.; Momoki, T.; et al. Genetic and antigenic characterisation of influenza A (H3N2) viruses isolated in Yokohama during the 2016/17 and 2017/18 influenza seasons. Eurosurveillance 2019, 24, 1800467. [Google Scholar] [CrossRef] [Green Version]
- Shiteng, H.; Ruijun, Y.; Lei, L.; Xufu, C.; Chenghua, Y.; Sheng, W. Molecular characteristics of hemagglutinin gene of influenza A (H3N2) virus in Quzhou, 2014–2015. Dis. Surveill. 2018, 33, 483–488. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Primer Description | Type/Subtype | Gene | Primer Name | Sequence 5′-3′ | Product Size (bp) | Ref. |
|---|---|---|---|---|---|---|
| Primers used for detection | IAV | M30F2/08 | ATGAGYCTTYTAACCGAGGTCGAAACG | 244 | [14] | |
| M264R3/08 | TGGACAAANCGTCTACGCTGCAG | |||||
| Primers used for typing | (H1N1) Pdm09 | HKU-SWF | TGAGCTCAGTGTCATCATTTGA | 174 | [14] | |
| HKU-SWR | TGCTGAGCTTTGGGTATGAA | |||||
| H3N2 | H3A1F6 | AAGCAGGGGATAATTCTATTAACC | 1127 | |||
| H3A1R1 | GTCTATCATTCCCTCCCAACCATT | |||||
| Primers used for sequencing | H3N2 | HA | H3A1F6 | AAGCAGGGGATAATTCTATTAACC | 1127 | [14] |
| H3A1R1 | GTCTATCATTCCCTCCCAACCATT | |||||
| HA828F | ACGAAGTGGGAAAAGCTCAATA | 934 | [15] | |||
| HA1778R | AGTAGAAACAAGGGTGTTTT | |||||
| H3N2 | NA | NA-1F | GAGCAAAAGCAGGAGTAAAG | 807 | ||
| NA787R | TGACAATGTGCTAGTATGAAC | |||||
| NA636F | AGATAGTGTTGTTTCATGGTC | 830 | ||||
| NA1413Rn | AGTAGAAACAAGGAGTTTTT |
| No. of Samples N (%) | Positive for Influenza A Virus N (%) | Positive for | |||
|---|---|---|---|---|---|
| H1N1, N (%) | H3N2, N (%) | ||||
| Total | 311 | 88 (28.3) | 45 (51.2) | 43 (48.8) | |
| Season | 2014–2015 | 24 (7.7) | - | - | - |
| 2015–2016 | 14 (4.5) | 14 (100) | 11 (78.5) | 3 (21.5) | |
| 2016–2017 | 42 (13.5) | 42 (100) | 22 (52.4) | 20 (47.6) | |
| 2017–2018 | 15 (4.8) | 15 (100) | 1 (6.7) | 14 (93.3) | |
| 2019–2020 | 216 (69.5) | 17 (7.9) | 11 (64.7) | 6 (35.3) | |
| Gender | Male | 155 (49.8) | 43 (27.7) | 23 (53.5) | 20 (46.5) |
| Female | 156 (50.2) | 45 (28.8) | 22 (48.9) | 23 (51.1) | |
| Age (Y) | 0–4 | 35 (11.3) | 8 (22.9) | 8 (100) | - |
| 5–14 | 8 (2.6) | 6 (75) a | 4 (66.6) | 2 (33.4) | |
| 15–64 | 200 (64.3) | 65 (32.5) b | 30 (46.2) | 35 (53.8) | |
| ≥65 | 68 (21.8) | 9 (13.2) | 3 (33.3) | 6 (66.7) | |
| No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Mutation sites | 5 | 126 | 212 | 220 | 303 | 329 | 344 | 463 | 468 |
| A-Singapore-Infimh-16-0019-2016 | Q | P | I | K | V | N | E | D | H |
| A-SaudiArabia-VRG-42-2020 | Q | L | V | N | I | S | K | D | P |
| A-SaudiArabia-VRG-46-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-48-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-49-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-57-2020 | K | L | V | N | I | S | K | D | P |
| A-SaudiArabia-VRG-138-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-46-2018 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-51-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-54-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-55-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-57-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-58-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-04-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-15-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-23-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-27-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-32-2017 | Q | P | A | N | I | S | E | D | H |
| A-SaudiArabia-VRG-44-2017 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-45-2017 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-01-2016 | Q | P | V | K | V | T | E | D | H |
| A-SaudiArabia-VRG-02-2016 | Q | P | V | K | V | N | E | D | H |
| A-SaudiArabia-VRG-03-2016 | Q | P | V | K | V | N | E | D | H |
| No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Mutation sites | 5 | 126 | 212 | 220 | 303 | 329 | 344 | 463 | 468 |
| A-Singapore-Infimh-16-0019-2016 | Q | P | I | K | V | N | E | D | H |
| A-SaudiArabia-VRG-42-2020 | Q | L | V | N | I | S | K | D | P |
| A-SaudiArabia-VRG-46-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-48-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-49-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-57-2020 | K | L | V | N | I | S | K | D | P |
| A-SaudiArabia-VRG-138-2020 | K | L | V | N | I | S | K | N | P |
| A-SaudiArabia-VRG-46-2018 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-51-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-54-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-55-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-57-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-58-2018 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-04-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-15-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-23-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-27-2017 | Q | P | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-32-2017 | Q | P | A | N | I | S | E | D | H |
| A-SaudiArabia-VRG-44-2017 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-45-2017 | Q | L | V | N | I | S | E | D | H |
| A-SaudiArabia-VRG-01-2016 | Q | P | V | K | V | T | E | D | H |
| A-SaudiArabia-VRG-02-2016 | Q | P | V | K | V | N | E | D | H |
| A-SaudiArabia-VRG-03-2016 | Q | P | V | K | V | N | E | D | H |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dudin, G.A.; Aziz, I.M.; Alzayed, R.M.; Ahmed, A.; Hussain, T.; Somily, A.M.; Alsaadi, M.M.; Almajhdi, F.N. Genetic Diversity and Evolutionary Kinetics of Influenza A Virus H3N2 Subtypes Circulating in Riyadh, Saudi Arabia. Vaccines 2023, 11, 702. https://doi.org/10.3390/vaccines11030702
Dudin GA, Aziz IM, Alzayed RM, Ahmed A, Hussain T, Somily AM, Alsaadi MM, Almajhdi FN. Genetic Diversity and Evolutionary Kinetics of Influenza A Virus H3N2 Subtypes Circulating in Riyadh, Saudi Arabia. Vaccines. 2023; 11(3):702. https://doi.org/10.3390/vaccines11030702
Chicago/Turabian StyleDudin, Gani Asa, Ibrahim M. Aziz, Rasha M. Alzayed, Anwar Ahmed, Tajamul Hussain, Ali M. Somily, Muslim M. Alsaadi, and Fahad N. Almajhdi. 2023. "Genetic Diversity and Evolutionary Kinetics of Influenza A Virus H3N2 Subtypes Circulating in Riyadh, Saudi Arabia" Vaccines 11, no. 3: 702. https://doi.org/10.3390/vaccines11030702