
Evaluation of the Association of Recombinant Proteins NanH and PknG from Corynebacterium pseudotuberculosis Using Different Adjuvants as a Recombinant Vaccine in Mice
 , , , and
, , , and 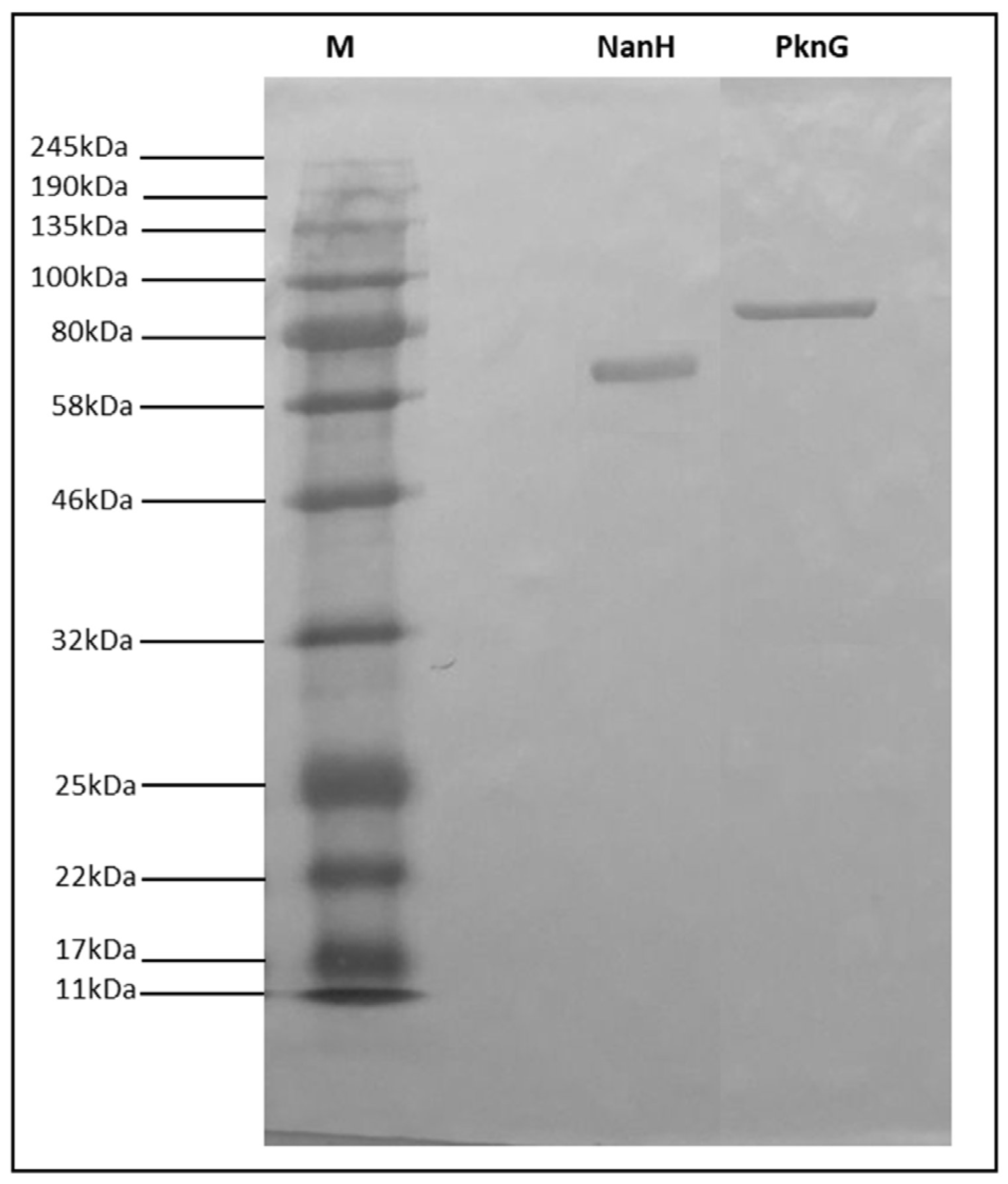{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Cloning, Transformation, and Selection of Recombinant Clones
2.3. Heterologous Expression of Proteins NanH and PknG from C. pseudotuberculosis in E. coli and Protein Purification
2.4. Western Blotting
2.5. Animals and Ethics Statement
2.6. Immunization and Challenge
2.7. Assessment of the Humoral Immune Response
2.8. Statistical Analyses
3. Results
3.1. Expression, Purification, and Identity Confirmation of Recombinant NanH and PknG Proteins
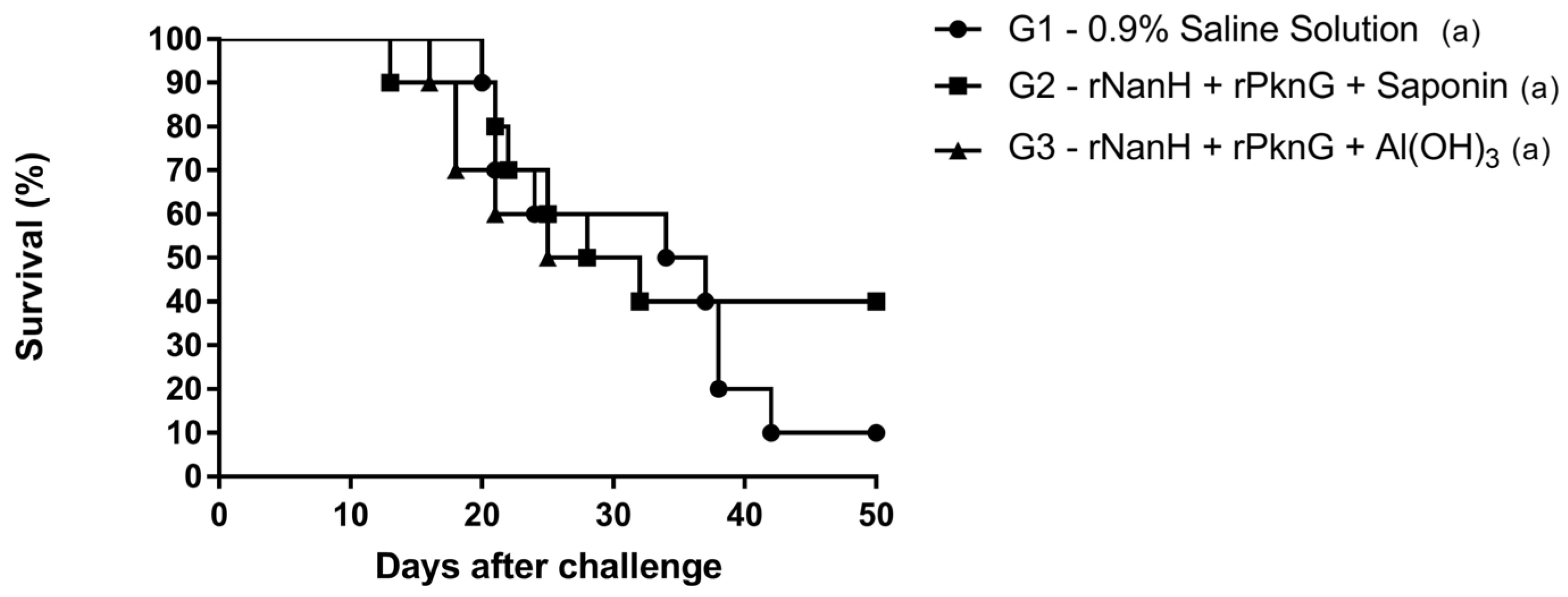
3.2. Evaluation of the Protective Potential of Vaccine Formulations
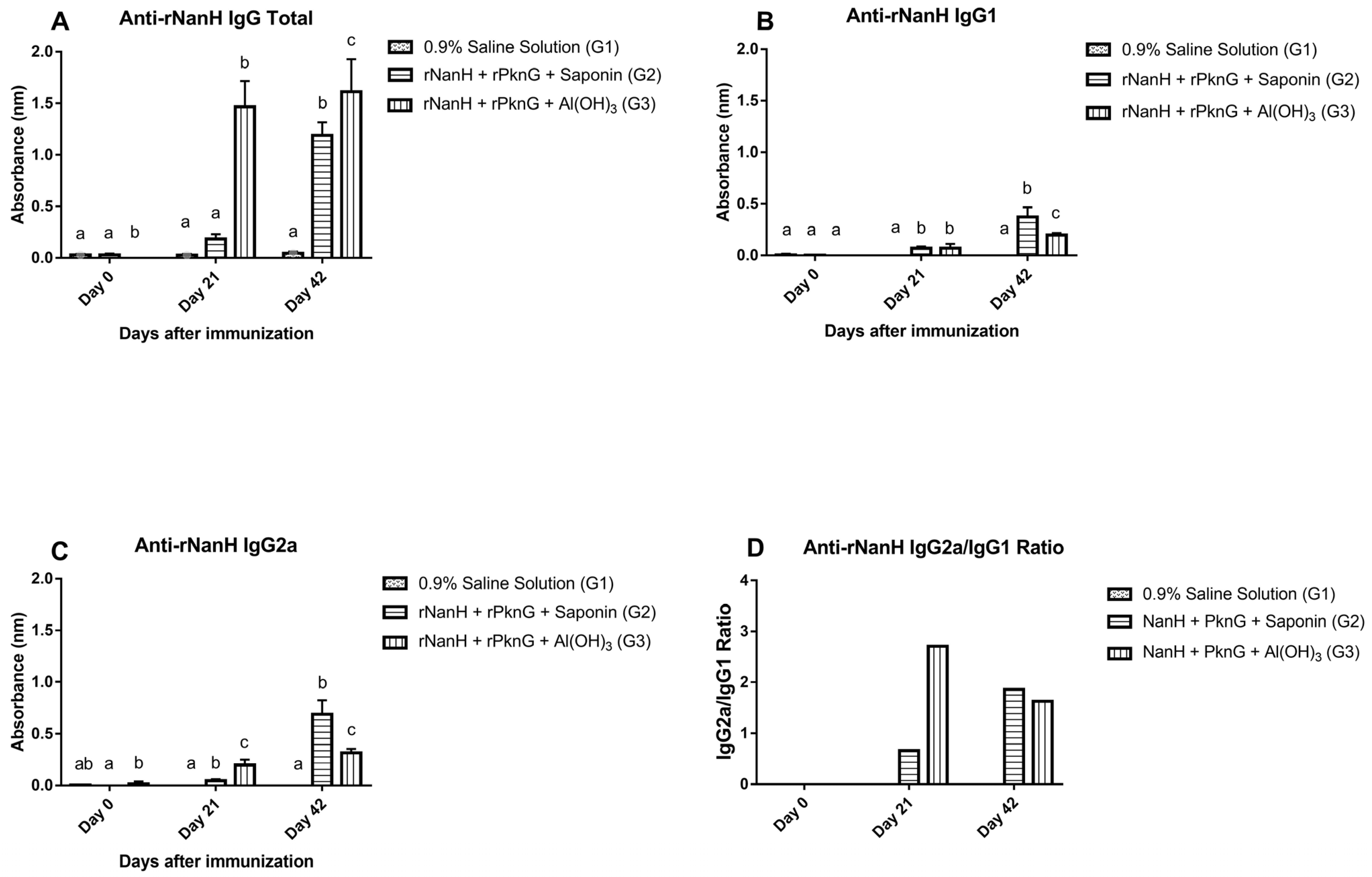
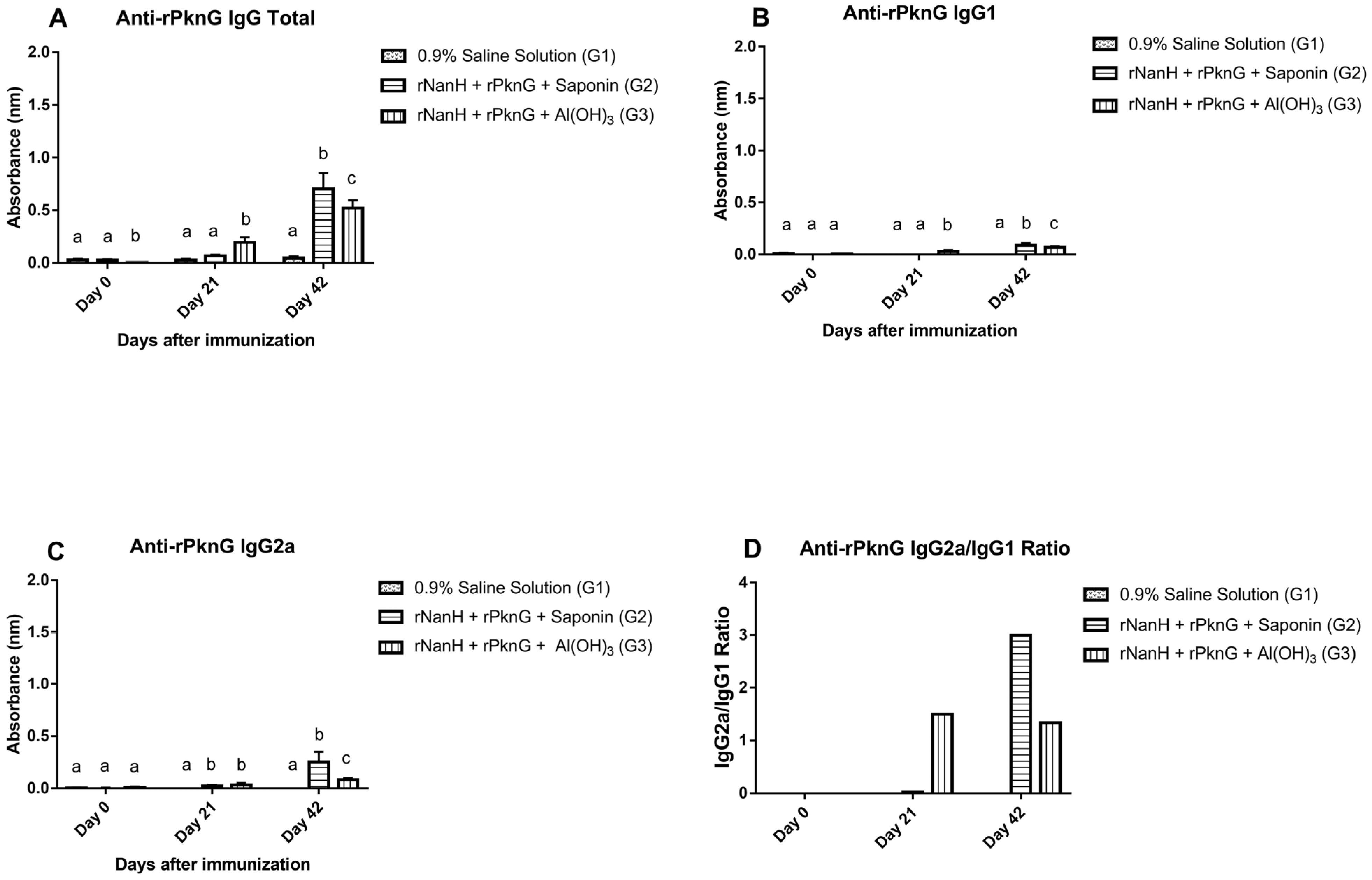
3.3. Assessment of the Humoral Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dorella, F.A.; Pacheco, L.G.C.; Oliveira, S.C.; Miyoshi, A.; Azevedo, V. Corynebacterium Pseudotuberculosis: Microbiology, Biochemical Properties, Pathogenesis and Molecular Studies of Virulence. Vet. Res. 2006, 37, 201–218. [Google Scholar] [CrossRef] [Green Version]
- Windsor, P.A. Control of Caseous Lymphadenitis. Vet. Clin. N. Am.—Food Anim. Pract. 2011, 27, 193–202. [Google Scholar] [CrossRef]
- Baird, G.J.; Fontaine, M.C. Corynebacterium Pseudotuberculosis and Its Role in Ovine Caseous Lymphadenitis. J. Comp. Pathol. 2007, 137, 179–210. [Google Scholar] [CrossRef] [PubMed]
- de Sá Guimarães, A.; do Carmo, F.B.; Pauletti, R.B.; Seyffert, N.; Ribeiro, D.; Lage, A.P.; Heinemann, M.B.; Miyoshi, A.; Azevedo, V.; Guimarães Gouveia, A.M. Caseous Lymphadenitis: Epidemiology, Diagnosis, and Control. IIOAB J. 2011, 2, 33–43. [Google Scholar]
- Costa, M.P.; McCulloch, J.A.; Almeida, S.S.; Dorella, F.A.; Fonseca, C.T.; Oliveira, D.M.; Teixeira, M.F.; Laskowska, E.; Lipinska, B.; Meyer, R.; et al. Molecular Characterization of the Corynebacterium Pseudotuberculosis Hsp60-Hsp10 Operon, and Evaluation of the Immune Response and Protective Efficacy Induced by Hsp60 DNA Vaccination in Mice. BMC Res. Notes 2011, 4, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Pinho, R.B.; de Oliveira Silva, M.T.; Bezerra, F.S.B.; Borsuk, S. Vaccines for Caseous Lymphadenitis: Up-to-Date and Forward-Looking Strategies. Appl. Microbiol. Biotechnol. 2021, 105, 2287–2296. [Google Scholar] [CrossRef]
- Leal, K.S.; de Oliveira Silva, M.T.; de Fátima Silva Rezende, A.; Bezerra, F.S.B.; Begnini, K.; Seixas, F.; Collares, T.; Dellagostin, O.; Portela, R.W.; de Carvalho Azevedo, V.A.; et al. Recombinant M. Bovis BCG Expressing the PLD Protein Promotes Survival in Mice Challenged with a C. Pseudotuberculosis Virulent Strain. Vaccine 2018, 36, 3578–3583. [Google Scholar] [CrossRef]
- Brum, A.A.; Rezende, A.D.F.S.; Brilhante, F.S.; Collares, T.; Begnine, K.; Seixas, F.K.; Collares, T.V.; Dellagostin, O.A.; Azevedo, V.; Santos, A.; et al. Recombinant Esterase from Corynebacterium Pseudotuberculosis in DNA and Subunit Recombinant Vaccines Partially Protects Mice against Challenge. J. Med. Microbiol. 2017, 66, 635–642. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Silva, M.T.; Bezerra, F.S.B.; de Pinho, R.B.; Begnini, K.R.; Seixas, F.K.; Collares, T.; Portela, R.D.; Azevedo, V.; Dellagostin, O.; Borsuk, S. Association of Corynebacterium Pseudotuberculosis Recombinant Proteins RCP09720 or RCP01850 with RPLD as Immunogens in Caseous Lymphadenitis Immunoprophylaxis. Vaccine 2018, 36, 74–83. [Google Scholar] [CrossRef]
- Barral, T.D.; Kalil, M.A.; Mariutti, R.B.; Arni, R.K.; Gismene, C.; Sousa, F.S.; Collares, T.; Seixas, F.K.; Borsuk, S.; Estrela-Lima, A.; et al. Immunoprophylactic Properties of the Corynebacterium Pseudotuberculosis-Derived MBP:PLD:CP40 Fusion Protein. Appl. Microbiol. Biotechnol. 2022, 106, 8035–8051. [Google Scholar] [CrossRef]
- Fu, M.; Su, H.; Su, Z.; Yin, Z.; Jin, J.; Wang, L.; Zhang, Q.; Xu, X. Transcriptome Analysis of Corynebacterium Pseudotuberculosis-Infected Spleen of Dairy Goats. Microb. Pathog. 2020, 147, 104370. [Google Scholar] [CrossRef] [PubMed]
- Trost, E.; Ott, L.; Schneider, J.; Schröder, J.; Jaenicke, S.; Goesmann, A.; Husemann, P.; Stoye, J.; Dorella, F.A.; Rocha, F.S.; et al. The Complete Genome Sequence of Corynebacterium Pseudotuberculosis FRC41 Isolated from a 12-Year-Old Girl with Necrotizing Lymphadenitis Reveals Insights into Gene-Regulatory Networks Contributing to Virulence. BMC Genom. 2010, 11, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana-Jorge, K.T.O.; Santos, T.M.; Tartaglia, N.R.; Aguiar, E.L.; Souza, R.F.S.; Mariutti, R.B.; Eberle, R.J.; Arni, R.K.; Portela, R.W.; Meyer, R.; et al. Putative Virulence Factors of Corynebacterium Pseudotuberculosis FRC41: Vaccine Potential and Protein Expression. Microb. Cell Fact. 2016, 15, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, L.S.; Lopes, N.D.R.; Pereira, V.C.; Andrade, C.L.B.; Torres, A.J.L.; Ribeiro, M.B.; Freire, S.M.; dos Santos, R.M.; D’ávila, M.; Nascimento, R.M.; et al. The Association of Bacterin and Recombinant Proteins Induces a Humoral Response in Sheep against Caseous Lymphadenitis. Vaccines 2022, 10, 1406. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Y.; Maqbool, B.; Yuan, L.; He, S.; Zhang, C.; Xu, W.; Hu, S. Early IgG Response to Foot and Mouth Disease Vaccine Formulated with a Vegetable Oil Adjuvant. Vaccines 2019, 7, 143. [Google Scholar] [CrossRef] [Green Version]
- Guy, B. The Perfect Mix: Recent Progress in Adjuvant Research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef]
- Lopes Bastos, B. Corynebacterium Pseudotuberculosis: Immunological Responses in Animal Models and Zoonotic Potential. J. Clin. Cell Immunol. 2012, 1, 10–4172. [Google Scholar] [CrossRef]
- Ramos, C.R.R.; Abreu, P.A.E.; Nascimento, A.L.T.O.; Ho, P.L. A High-Copy T7 Escherichia Coli Expression Vector for the Production of Recombinant Proteins with a Minimal N-Terminal His-Tagged Fusion Peptide. Brazilian J. Med. Biol. Res. 2004, 37, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.W.; Droppa-Almeida, D.; Borsuk, S.; Azevedo, V.; Portela, R.W.; Miyoshi, A.; Rocha, F.S.; Dorella, F.A.; Vivas, W.L.; Padilha, F.F.; et al. Corynebacterium Pseudotuberculosis Cp09 Mutant and Cp40 Recombinant Protein Partially Protect Mice against Caseous Lymphadenitis. BMC Vet. Res. 2014, 10, 965. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Silva, M.T.; de Pinho, R.B.; Bezerra, F.S.B.; Campos, V.F.; Azevedo, V.; Borsuk, S. Establishment of an Objective Endpoint in Mice Model for Caseous Lymphadenitis Vaccine Trials. Vet. Microbiol. 2019, 230, 86–89. [Google Scholar] [CrossRef]
- Droppa-Almeida, D.; Vivas, W.L.P.; Silva, K.K.O.; Rezende, A.F.S.; Simionatto, S.; Meyer, R.; Lima-Verde, I.B.; Delagostin, O.; Borsuk, S.; Padilha, F.F. Recombinant CP40 from Corynebacterium Pseudotuberculosis Confers Protection in Mice after Challenge with a Virulent Strain. Vaccine 2016, 34, 1091–1096. [Google Scholar] [CrossRef]
- Thais de Oliveira Silva, M.; Barros de Pinho, R.; da Rocha Fonseca, B.; Silvestre Brilhante Bezerra, F.; Severo Sabedra Sousa, F.; Kommling Seixas, F.; Collares, T.; José Meyer Nascimento, R.; Wagner Portela, R.; Ariston Carvalho Azevedo, V.; et al. NanH and PknG Putative Virulence Factors as a Recombinant Subunit Immunogen against Corynebacterium Pseudotuberculosis Infection in Mice. Vaccine 2020, 38, 8099–8106. [Google Scholar] [CrossRef]
- Rezende, A.F.S.; Brum, A.A.; Bezerra, F.S.B.; Braite, D.C.; Sá, G.L.; Thurow, H.S.; Seixas, F.K.; Azevedo, V.A.C.; Portela, R.W.; Borsuk, S. Assessment of the Acid Phosphatase CP01850 from Corynebacterium Pseudotuberculosis in DNA and Subunit Vaccine Formulations against Caseous Lymphadenitis. Arq. Bras. Med. Vet. e Zootec. 2020, 72, 199–207. [Google Scholar] [CrossRef]
- Droppa-Almeida, D.; Da Silva, G.A.; Do Amorim Costa Gaspar, L.M.; Pereyra, B.B.S.; Nascimento, R.J.M.; Borsuk, S.; Franceschi, E.; Padilha, F.F. Peptide Vaccines Designed with the Aid of Immunoinformatic against Caseous Lymphadenitis Promotes Humoral and Cellular Response Induction in Mice. PLoS ONE 2021, 16, e0256864. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Advances in Saponin-Based Adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Jacob, E.; Hod-Dvorai, R.; Ben-Mordechai, O.L.; Boyko, Y.; Avni, O. Dual Function of Polycomb Group Proteins in Differentiated Murine T Helper (CD4+) Cells. J. Mol. Signal. 2011, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, B. Modulation of Immune Response in Experimental Chagas Disease. World J. Exp. Med. 2013, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumsden, J.M.; Nurmukhambetova, S.; Klein, J.H.; Sattabongkot, J.; Bennett, J.W.; Bertholet, S.; Fox, C.B.; Reed, S.G.; Ockenhouse, C.F.; Howard, R.F.; et al. Evaluation of Immune Responses to a Plasmodium Vivax CSP-Based Recombinant Protein Vaccine Candidate in Combination with Second-Generation Adjuvants in Mice. Vaccine 2012, 30, 3311–3319. [Google Scholar] [CrossRef]
- Bezerra, F.S.B.; de Oliveira Silva, M.T.; Rezende, A.D.F.S.; Lopes, A.S.; de Pinho, R.B.; Seixas, F.K.; Collares, T.V.; Portela, R.W.D.; Azevedo, V.A. de C.; Borsuk, S. Saponin-Adjuvanted Recombinant Vaccines Containing RCP00660, RCP09720 or RCP01850 Proteins against Corynebacterium Pseudotuberculosis Infection in Mice. Vaccine 2021, 39, 2568–2574. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scholl, N.R.; Silva, M.T.d.O.; Barbosa, T.N.; de Pinho, R.B.; Alves, M.S.D.; Portela, R.W.; Azevedo, V.A.d.C.; Borsuk, S. Evaluation of the Association of Recombinant Proteins NanH and PknG from Corynebacterium pseudotuberculosis Using Different Adjuvants as a Recombinant Vaccine in Mice. Vaccines 2023, 11, 519. https://doi.org/10.3390/vaccines11030519
Scholl NR, Silva MTdO, Barbosa TN, de Pinho RB, Alves MSD, Portela RW, Azevedo VAdC, Borsuk S. Evaluation of the Association of Recombinant Proteins NanH and PknG from Corynebacterium pseudotuberculosis Using Different Adjuvants as a Recombinant Vaccine in Mice. Vaccines. 2023; 11(3):519. https://doi.org/10.3390/vaccines11030519
Chicago/Turabian StyleScholl, Nicole Ramos, Mara Thais de Oliveira Silva, Tallyson Nogueira Barbosa, Rodrigo Barros de Pinho, Mirna Samara Dié Alves, Ricardo Wagner Portela, Vasco Ariston de Carvalho Azevedo, and Sibele Borsuk. 2023. "Evaluation of the Association of Recombinant Proteins NanH and PknG from Corynebacterium pseudotuberculosis Using Different Adjuvants as a Recombinant Vaccine in Mice" Vaccines 11, no. 3: 519. https://doi.org/10.3390/vaccines11030519