
A Vaccine Targeting Ovine Herpesvirus 2 Glycoprotein B Protects against Sheep-Associated Malignant Catarrhal Fever
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine Formulations
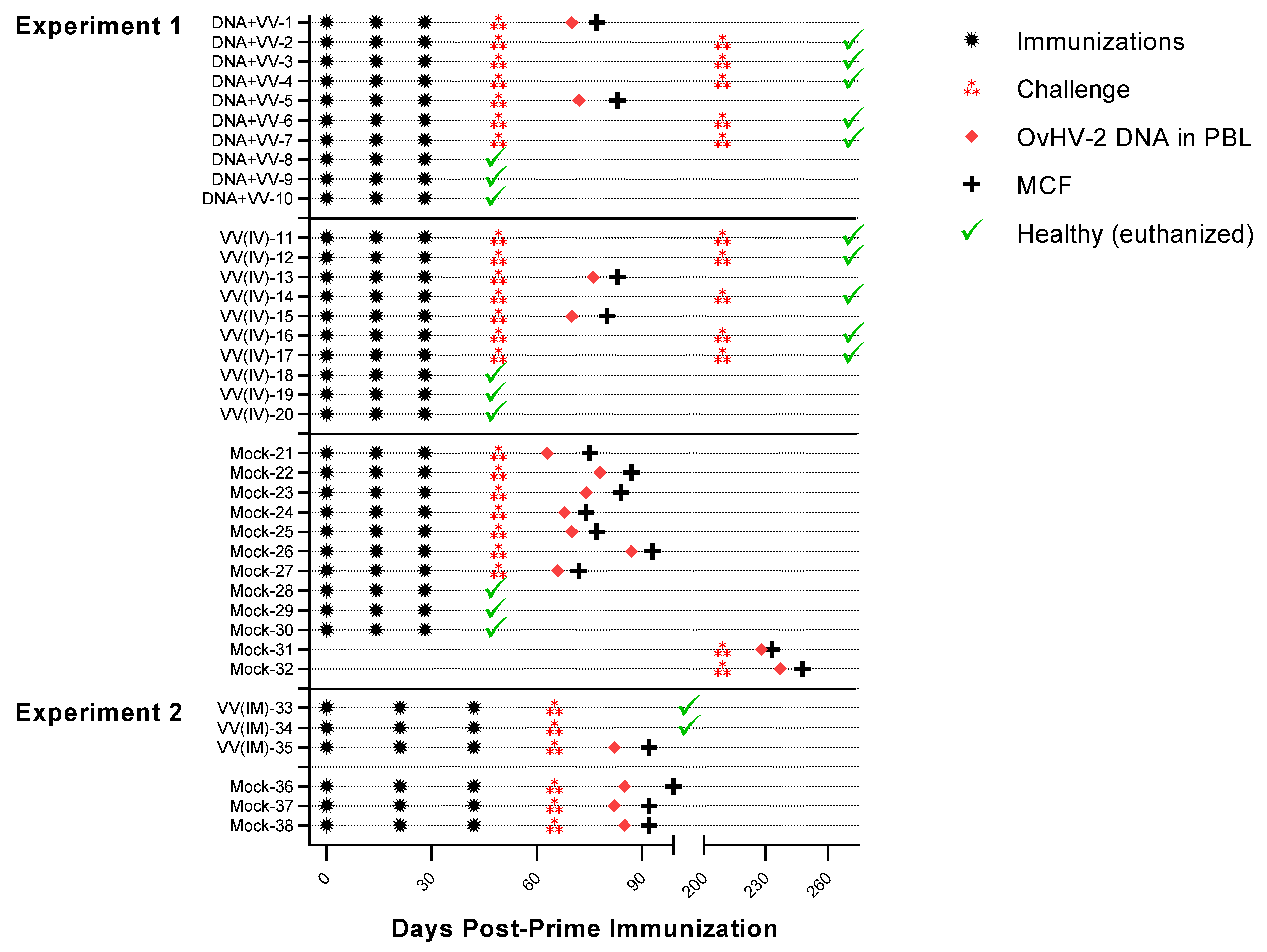
2.3. Immunizations and Challenge
2.4. Animal Monitoring
2.4.1. Pathology
2.4.2. Viral DNA Analysis
2.4.3. Antibody Response Analysis
2.5. Statistical Analysis
3. Results
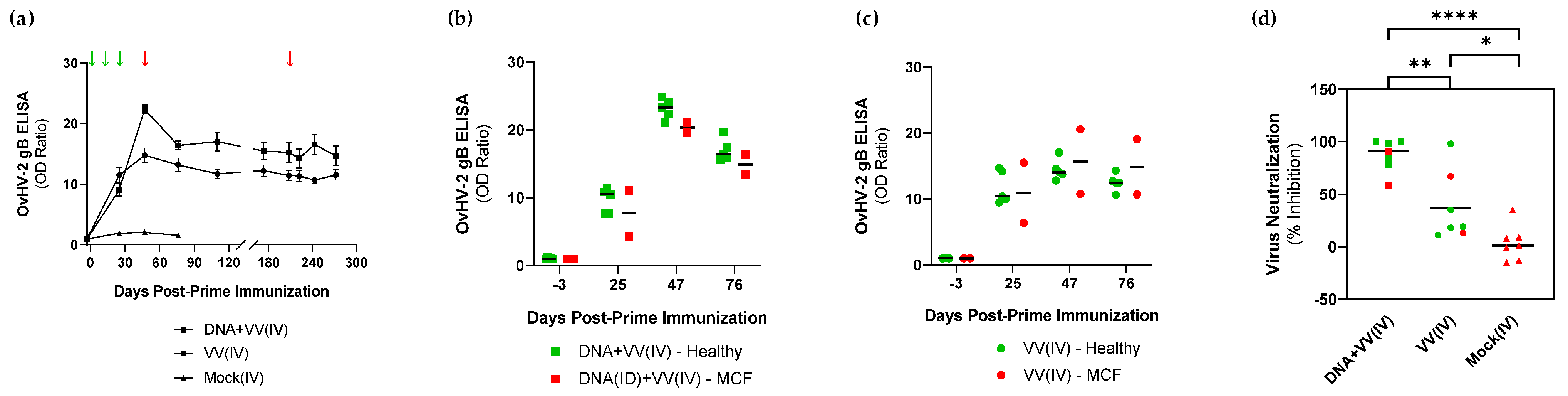
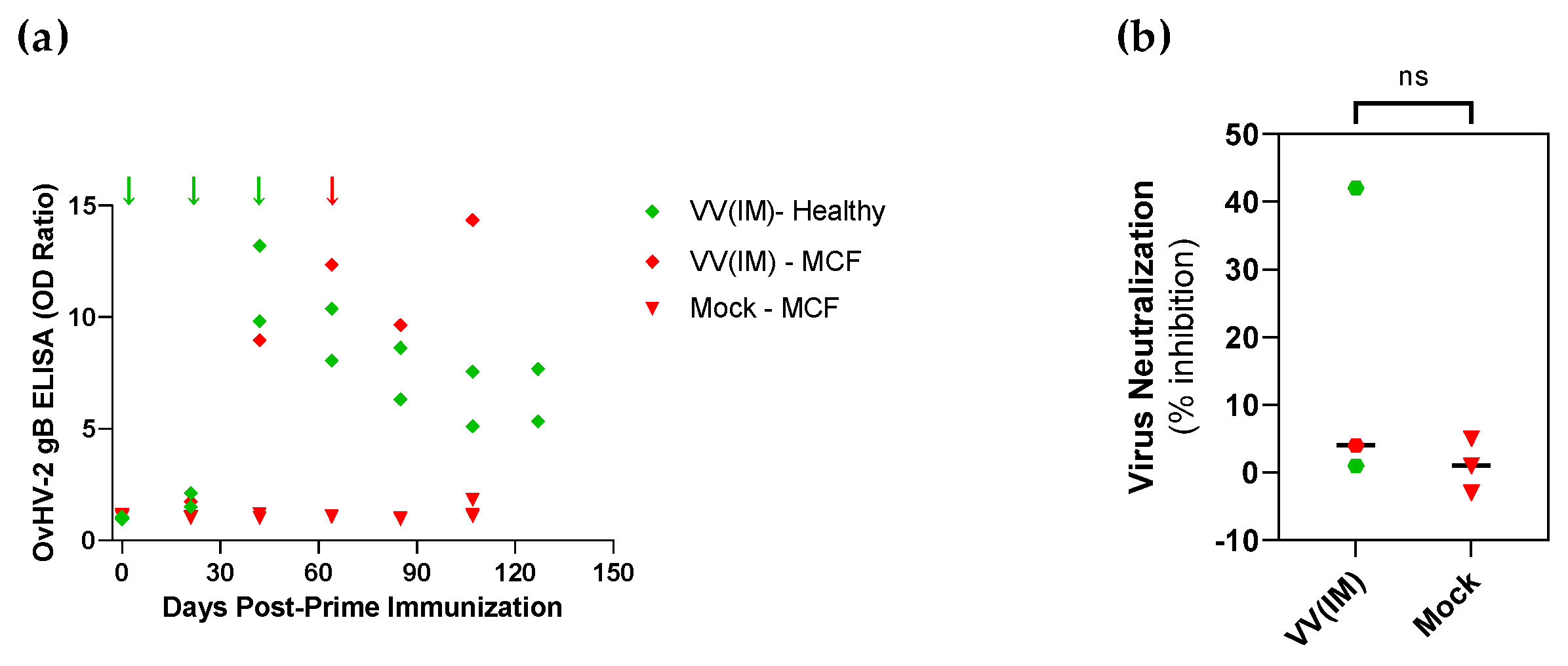
3.1. Safety and Protective Efficacy of OvHV-2-gB Vaccine Candidates
3.2. Immunogenicity Assessment of OvHV-2-gB Vaccine Platforms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davison, A.; Eberle, R.; Ehlers, B.; Hayward, G.; McGeoch, D.; Minson, A.; Pellett, P.; Roizman, B.; Studdert, M.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Plowright, W. Malignant catarrhal fever virus. In Virus Infections of Ruminants, 3rd ed.; Dinter, Z., Morein, B., Eds.; Elsevier Science Publishers B.V.: New York, NY, USA, 1990; pp. 123–150. [Google Scholar]
- Russell, G.C.; Stewart, J.P.; Haig, D.M. Malignant catarrhal fever: A review. Vet. J. 2009, 179, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Hemadri, D.; Bhatia, S. Sheep associated malignant catarrhal fever: An emerging disease of bovids in India. Indian J. Virol. 2013, 24, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, J.; Ackermann, M.; Jayawardane, G.; Russell, G.; Haig, D.M.; Reid, H.; Stewart, J.P. Complete sequence and analysis of the ovine herpesvirus 2 genome. J. Gen. Virol. 2007, 88, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Taus, N.S.; Cunha, C.W.; Jana, M.; O’Toole, D.; Li, H. Cross-reactivity of neutralizing antibodies among malignant catarrhal fever viruses. PLoS ONE 2015, 10, e0145073. [Google Scholar] [CrossRef]
- Taus, N.S.; Herndon, D.R.; Traul, D.L.; Stewart, J.P.; Ackermann, M.; Li, H.; Knowles, D.P.; Lewis, G.S.; Brayton, K.A. Comparison of ovine herpesvirus 2 genomes isolated from domestic sheep (Ovis aries) and a clinically affected cow (Bos bovis). J. Gen. Virol. 2007, 88, 40–45. [Google Scholar] [CrossRef]
- Lankester, F.; Lugelo, A.; Kazwala, R.; Keyyu, J.; Cleaveland, S.; Yoder, J. The Economic Impact of Malignant Catarrhal Fever on Pastoralist Livelihoods. PLoS ONE 2015, 10, e0116059. [Google Scholar] [CrossRef]
- Li, H.; Cunha, C.W.; Taus, N.S.; Knowles, D.P. Malignant catarrhal fever: Inching toward understanding. Annu. Rev. Anim. Biosci. 2014, 2, 209–233. [Google Scholar] [CrossRef]
- Berezowski, J.A.; Appleyard, G.D.; Crawford, T.B.; Haigh, J.; Li, H.; Middleton, D.M.; O’Connor, B.P.; West, K.; Woodbury, M. An outbreak of sheep-associated malignant catarrhal fever in bison (Bison bison) after exposure to sheep at a public auction sale. J. Vet. Diagn. Investig. 2005, 17, 55–58. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Taus, N.S.; Jones, c.; Murphy, B.; Evermann, J.F.; Crawford, T.B. A devastating outbreak of malignant catarrhal fever in a bison feedlot. J. Vet. Diagn. Investig. 2006, 18, 119–123. [Google Scholar] [CrossRef]
- Cunha, C.W.; Taus, N.S.; Dewals, B.G.; Vanderplasschen, A.; Knowles, D.P.; Li, H. Replacement of glycoprotein B in alcelaphine herpesvirus 1 by Its ovine herpesvirus 2 homolog: Implications in vaccine development for sheep-associated malignant catarrhal fever. mSphere 2016, 1, e00108–e00116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirangi, P.K. Attempts to immunize cattle against virulent African malignant catarrhal fever virus (alcelaphine herpesvirus-1) with a herpesvirus isolated from American cattle. Vet. Microbiol. 1991, 28, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Plowright, W.; Herniman, K.A.J.; Jessett, D.M.; Kalunda, M.; Rampton, C.S. Immunisation of cattle against the herpesvirus of malignant catarrhal fever: Failure of inactivated culture vaccines with adjuvant. Res. Vet. Sci. 1975, 19, 159–166. [Google Scholar] [CrossRef]
- Shringi, S.; O’Toole, D.; Cole, E.; Baker, K.N.; White, S.N.; Donofrio, G.; Li, H.; Cunha, C.W. OvHV-2 glycoprotein B delivered by a recombinant BoHV-4 is immunogenic and induces partial protection against sheep-associated malignant catarrhal fever in a rabbit model. Vaccines 2021, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Wambua, L.; Wambua, P.N.; Ramogo, A.M.; Mijele, D.; Otiende, M.Y. Wildebeest-associated malignant catarrhal fever: Perspectives for integrated control of a lymphoproliferative disease of cattle in sub-Saharan Africa. Arch. Virol. 2016, 161, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, H.; Stewart, J.P.; Ireri, R.G.; Campbell, I.; Pow, I.; Reid, H.W.; Haig, D.M. Genome re-arrangements associated with loss of pathogenicity of the gamma-herpesvirus alcelaphine herpesvirus-1. Res. Vet. Sci. 2003, 75, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Haig, D.M.; Grant, D.; Deane, D.; Campbell, I.; Thomson, J.; Jepson, C.; Buxton, D.; Russell, G.C. An immunisation strategy for the protection of cattle against alcelaphine herpesvirus-1-induced malignant catarrhal fever. Vaccine 2008, 26, 4461–4468. [Google Scholar] [CrossRef]
- Russell, G.; Benavides, J.; Grant, D.; Todd, H.; Deane, D.; Percival, A.; Thomson, J.; Connelly, M.; Haig, D. Duration of protective immunity and antibody responses in cattle immunised against alcelaphine herpesvirus-1-induced malignant catarrhal fever. Vet. Res. 2012, 43, 51. [Google Scholar] [CrossRef] [Green Version]
- Lankester, F.; Russell, G.C.; Lugelo, A.; Ndabigaye, A.; Mnyambwa, N.; Keyyu, J.; Kazwala, R.; Grant, D.; Percival, A.; Deane, D.; et al. A field vaccine trial in Tanzania demonstrates partial protection against malignant catarrhal fever in cattle. Vaccine 2016, 34, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Lankester, F.; Lugelo, A.; Werling, D.; Mnyambwa, N.; Keyyu, J.; Kazwala, R.; Grant, D.; Smith, S.; Parameswaran, N.; Cleaveland, S.; et al. The efficacy of alcelaphine herpesvirus-1 (AlHV-1) immunization with the adjuvants Emulsigen® and the monomeric TLR5 ligand FliC in zebu cattle against AlHV-1 malignant catarrhal fever induced by experimental virus challenge. Vet. Microbiol. 2016, 195, 144–153. [Google Scholar] [CrossRef]
- Parameswaran, N.; Russell, G.C.; Bartley, K.; Grant, D.M.; Deane, D.; Todd, H.; Dagleish, M.P.; Haig, D.M. The effect of the TLR9 ligand CpG-oligodeoxynucleotide on the protective immune response to alcelaphine herpesvirus-1-mediated malignant catarrhal fever in cattle. Veterianry Res. 2014, 45, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, E.; Russell, G.; Grant, D.; Mutisya, C.; Omoto, L.; Dobson, E.; Lankester, F.; Nene, V. A randomised vaccine field trial in Kenya demonstrates protection against wildebeest-associated malignant catarrhal fever in cattle. Vaccine 2019, 37, 5946–5953. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.W.; Knowles, D.P.; Taus, N.S.; O’Toole, D.; Nicola, A.V.; Aguilar, H.C.; Li, H. Antibodies to ovine herpesvirus 2 glycoproteins decrease virus infectivity and prevent malignant catarrhal fever in rabbits. Vet. Microbiol. 2015, 175, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, L.; Sorel, O.; Van Campe, W.; Boudry, C.; Roels, S.; Myster, F.o.; Reschner, A.; Coulie, P.G.; Kerkhofs, P.; Vanderplasschen, A.; et al. An essential role for γ-herpesvirus latency-associated nuclear antigen homolog in an acute lymphoproliferative disease of cattle. Proc. Natl. Acad. Sci. USA 2013, 110, E1933–E1942. [Google Scholar] [CrossRef] [Green Version]
- Gailbreath, K.L.; Taus, N.S.; Cunha, C.W.; Knowles, D.P.; Li, H. Experimental infection of rabbits with ovine herpesvirus 2 from sheep nasal secretions. Vet. Microbiol. 2008, 132, 65–73. [Google Scholar] [CrossRef]
- Cunha, C.W.; O’Toole, D.; Taus, N.S.; Knowles, D.P.; Li, H. Are rabbits a suitable model to study sheep-associated malignant catarrhal fever in susceptible hosts? Vet. Microbiol. 2013, 163, 358–363. [Google Scholar] [CrossRef]
- Hussy, D.; Stauber, N.; Leutenegger, C.M.; Rieder, S.; Ackermann, M. Quantitative fluorogenic PCR assay for measuring ovine herpesvirus 2 replication in sheep. Clin. Diagn. Lab. Immunol. 2001, 8, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Traul, D.L.; Taus, N.S.; Oaks, J.L.; O’Toole, D.; Rurangirwa, F.R.; Baszler, T.V.; Li, H. Validation of non-nested and real-time PCR for diagnosis of sheep-associated malignant catarrahal fever in clinical samples. J. Vet. Diagn. Investig. 2007, 19, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Traul, D.L.; Elias, S.; Taus, N.S.; Herrmann, L.M.; Oaks, J.L.; Li, H. A real-time PCR assay for measuring alcelaphine herpesvirus-1 DNA. J. Virol. Methods 2005, 129, 186–190. [Google Scholar] [CrossRef]
- Gailbreath, K.L.; O’Toole, D.; Taus, N.S.; Knowles, D.P.; Oaks, J.L.; Li, H. Experimental nebulization of American bison (Bison bison) with low doses of ovine herpesvirus 2 from sheep nasal secretions. Vet. Microbiol. 2010, 143, 389–393. [Google Scholar] [CrossRef]
- Li, H.; Cunha, C.W.; O’Toole, D.; Nicola, A.V.; Knowles, D.P.; Taus, N.S. Development of an in vivo system to measure antibody-blocking of ovine herpesvirus 2 entry. J. Virol. Methods 2013, 188, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Jorge, S.; Dellagostin, O.A. The development of veterinary vaccines: A review of traditional methods and modern biotechnology approaches. Biotechnol. Res. Innov. 2017, 1, 6–13. [Google Scholar] [CrossRef]
- Meeusen, E.N.T.; Walker, J.; Peters, A.; Pastoret, P.-P.; Jungersen, G. Current Status of Veterinary Vaccines. Clin. Microbiol. Rev. 2007, 20, 489–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, G.C.; Haig, D.M.; Dagleish, M.P.; Todd, H.; Percival, A.; Grant, D.M.; Thomson, J.; Karagianni, A.E.; Benavides, J. Analysis of immune responses to attenuated alcelaphine herpesvirus 1 formulated with and without adjuvant. Vaccine X 2021, 8, 100090. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Number of Rabbits | Vaccination | ||
|---|---|---|---|---|
| Immunogen 2 | Regimen | |||
| Experiment 1 | ||||
| DNA+VV(IV) | 10 | pOvHV-2-ORF8 + AlHV-1∆ORF73/OvHV-2-ORF8 | DNA prime and 1st booster, intradermal VV 2nd booster, intravenous | |
| VV(IV) | 10 | AlHV-1∆ORF73/OvHV-2-ORF8 | VV prime and 2 boosters, intravenous | |
| Mock(IV) | 10 | Mock | 3 inoculations, intravenous | |
| Experiment 2 | ||||
| VV(IM) | 3 | AlHV-1∆ORF73/OVHV-2-ORF8 | VV prime and 2 boosters, intramuscular | |
| Mock(IM) | 3 | Mock | 3 inoculations, intramuscular | |
| Experiment 1 | Experiment 2 | |||||
|---|---|---|---|---|---|---|
| DNA+VV(IV) | VV(IV) | Mock | VV(IM) | Mock | ||
| Protected (Healthy) | 5 (71.4%) | 5 (71.4%) | 0 (0%) | 2 (66.7%) | 0 (0%) | |
| Unprotected (MCF) | 2 (28.6%) | 2 (28.6%) | 7 (100%) | 1 (33.3%) | 3 (100%) | |
| Total | 7 | 7 | 7 | 3 | 3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunha, C.W.; Baker, K.N.; O’Toole, D.; Cole, E.; Shringi, S.; Dewals, B.G.; Vanderplasschen, A.; Li, H. A Vaccine Targeting Ovine Herpesvirus 2 Glycoprotein B Protects against Sheep-Associated Malignant Catarrhal Fever. Vaccines 2022, 10, 2156. https://doi.org/10.3390/vaccines10122156
Cunha CW, Baker KN, O’Toole D, Cole E, Shringi S, Dewals BG, Vanderplasschen A, Li H. A Vaccine Targeting Ovine Herpesvirus 2 Glycoprotein B Protects against Sheep-Associated Malignant Catarrhal Fever. Vaccines. 2022; 10(12):2156. https://doi.org/10.3390/vaccines10122156
Chicago/Turabian StyleCunha, Cristina W., Katherine N. Baker, Donal O’Toole, Emily Cole, Smriti Shringi, Benjamin G. Dewals, Alain Vanderplasschen, and Hong Li. 2022. "A Vaccine Targeting Ovine Herpesvirus 2 Glycoprotein B Protects against Sheep-Associated Malignant Catarrhal Fever" Vaccines 10, no. 12: 2156. https://doi.org/10.3390/vaccines10122156