
Infection, Transmission, Pathogenesis and Vaccine Development against Mycoplasma gallisepticum
 , , and
, , and
Abstract
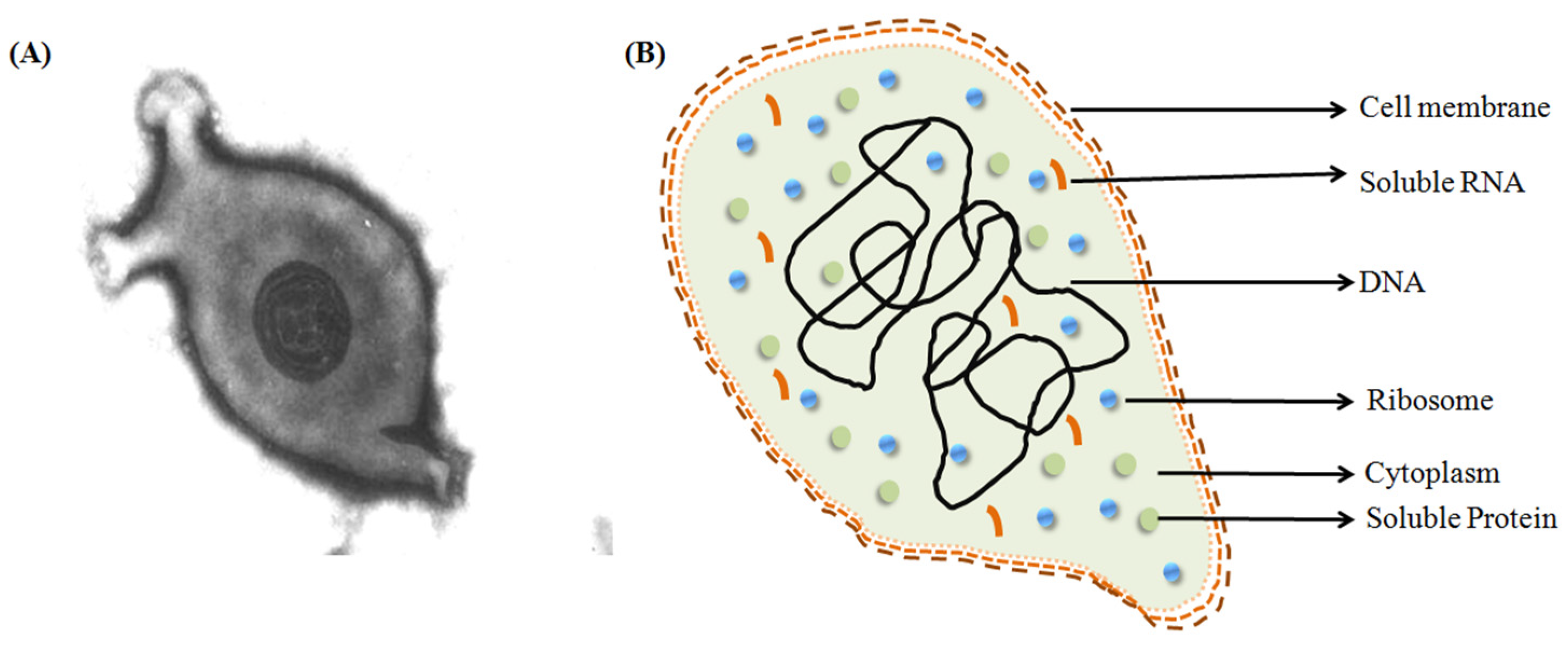
:1. Introduction
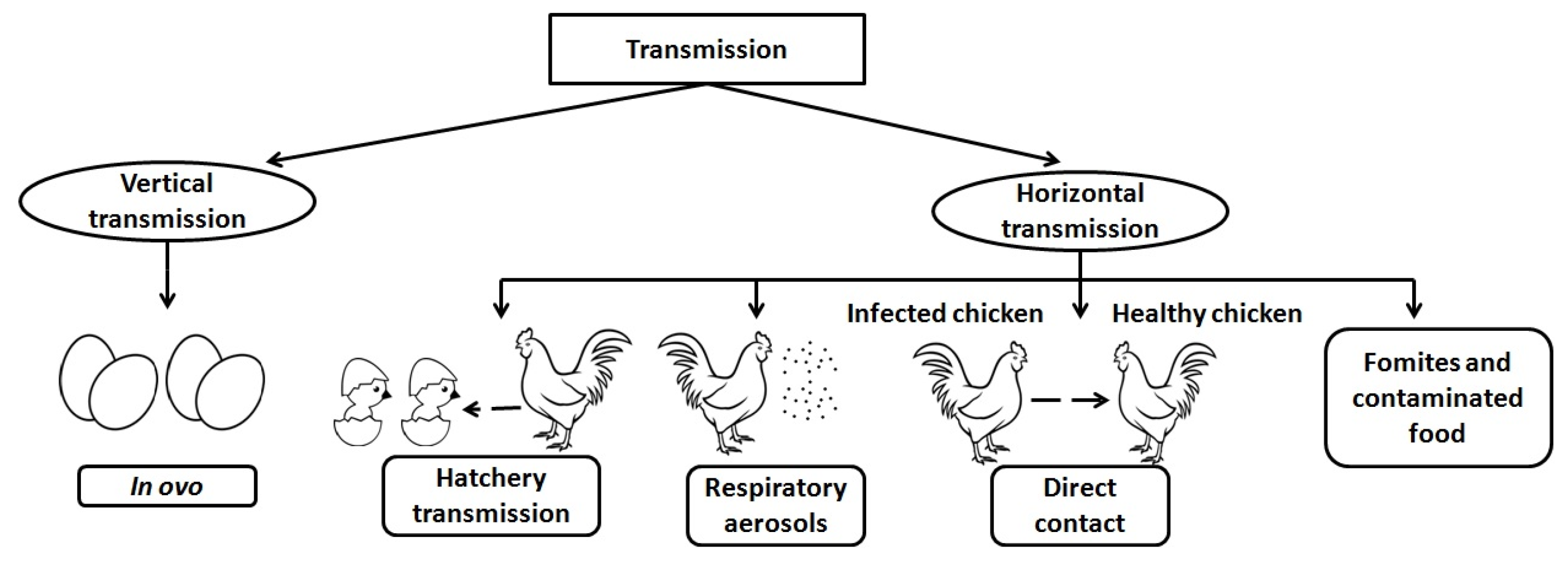
2. Transmission
2.1. Vertical
2.2. Horizontal
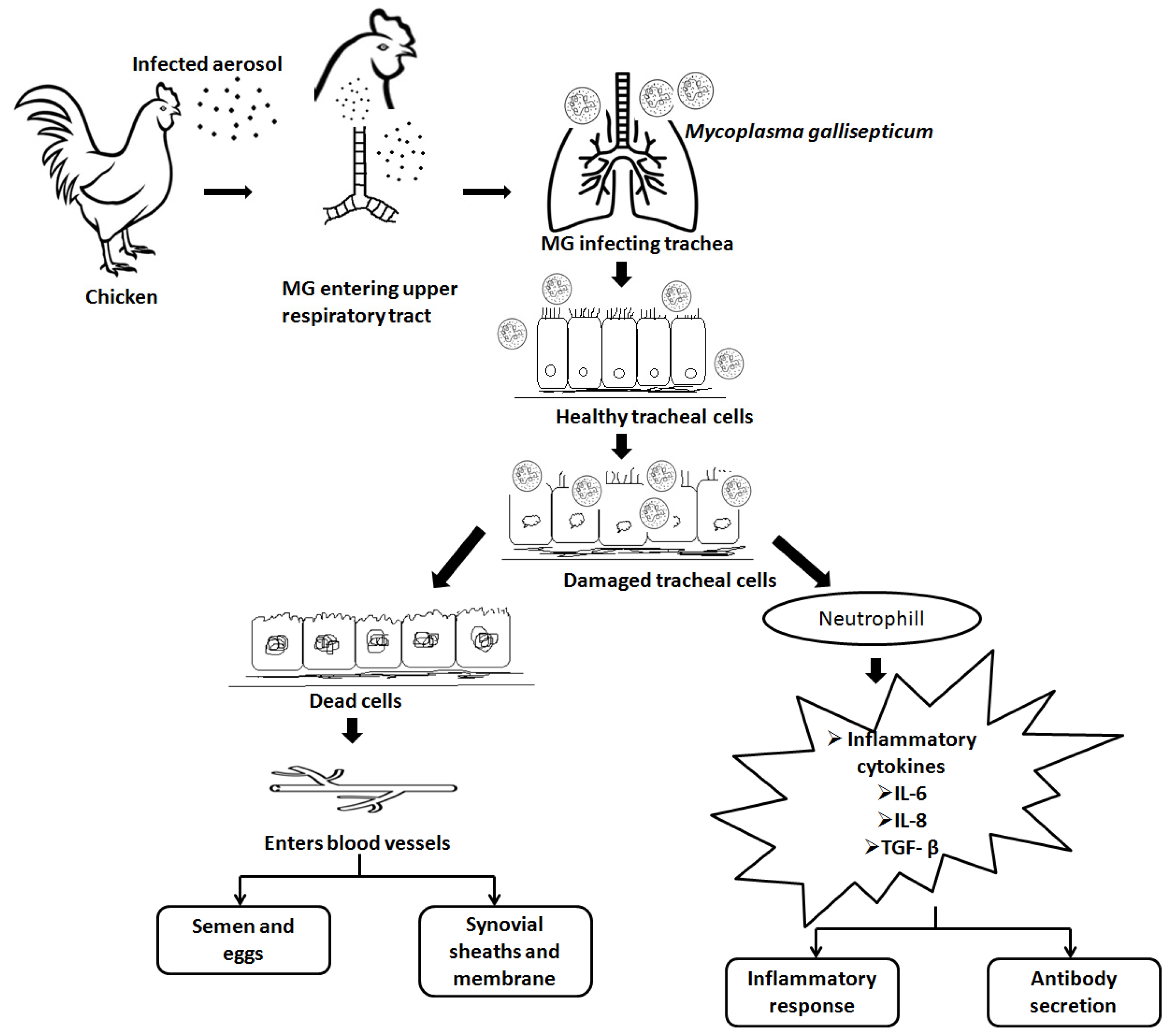
3. Pathogenesis and Host Immune Response
4. Diagnosis of Mycoplasma gallisepticum
5. Virulence and Immune Evasion Proteins
5.1. Adhesions
5.2. Immune Evasion
5.3. Phase Variation by Mycoplasmas
5.4. Heat Shock Proteins
6. Prevention from M. gallisepticum
7. Currently Available Vaccines against M. gallisepticum
7.1. Bacterins/Inactivated Vaccine
7.2. Live-Attenuated Vaccines
7.2.1. First Generation (1975–2000)
7.2.2. Second Generation (2000-Present)
7.3. Genetic Engineered Vaccines
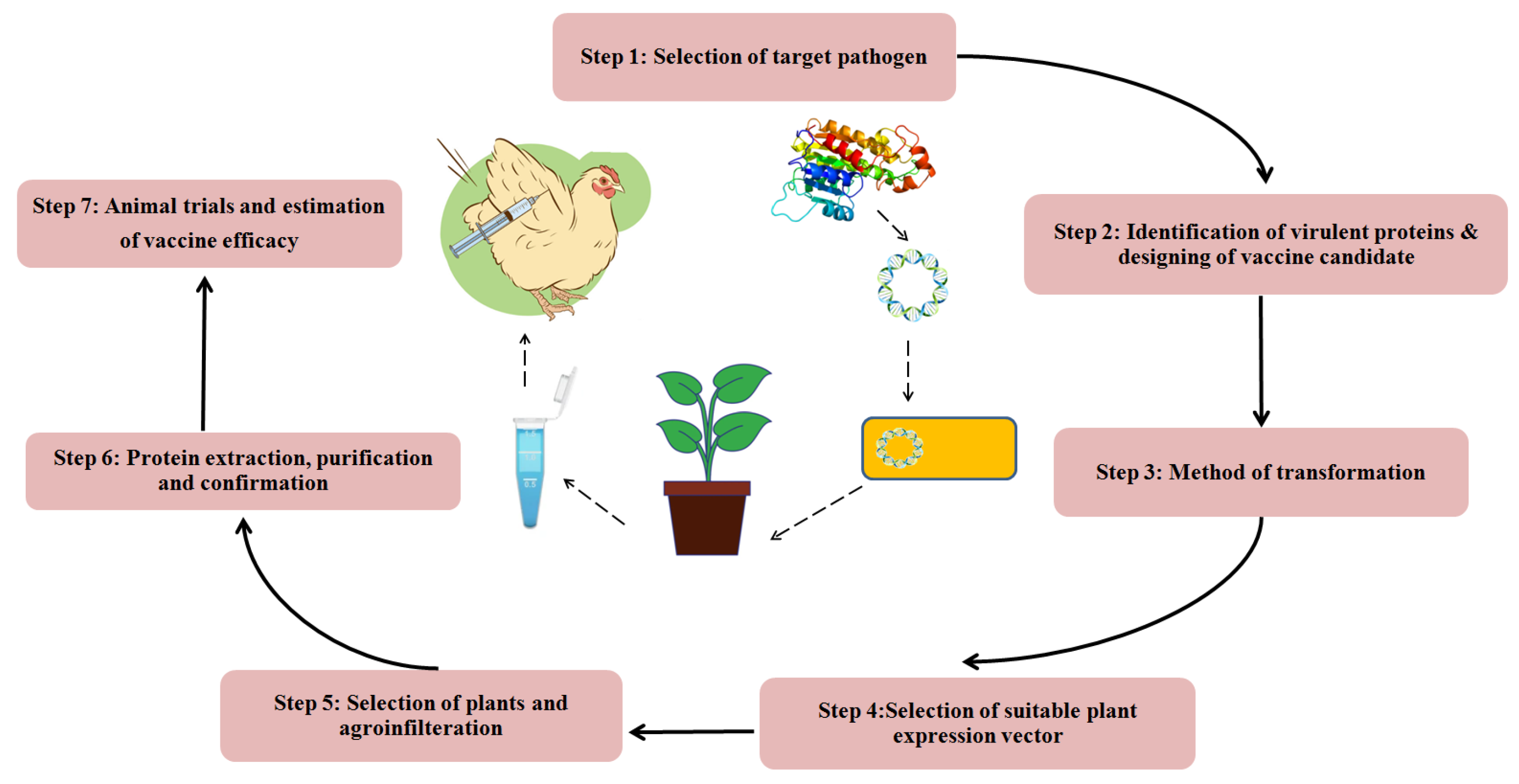
8. Future Vaccines
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Razin, S.; Yogev, D.; Naot, Y. Molecular Biology and Pathogenicity of Mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himmelreich, R.; Hilbert, H.; Plagens, H.; Pirkl, E.; Li, B.-C.; Herrmann, R. Complete Sequence Analysis of the Genome of the Bacterium Mycoplasma Pneumoniae. Nucleic Acids Res. 1996, 24, 4420–4449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayflick, L.; Chanock, R.M. Mycoplasma Species of Man. Bacteriol. Rev. 1965, 29, 185–221. [Google Scholar] [CrossRef] [PubMed]
- Dybvig, K.; Voelker, L.L. Molecular biology of Mycoplasmas. Annu. Rev. Microbiol. 1996, 50, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Dhondt, A.A.; Tessaglia, D.L.; Slothower, R.L. Epidemic Mycoplasmal conjunctivitis in house finches from eastern north America. J. Wildl. Dis. 1998, 34, 265–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stipkovits, L.; Kempf, I. Mycoplasmoses in Poultry. Rev. Sci. Tech. OIE 1996, 15, 1495–1525. [Google Scholar] [CrossRef]
- Levisohn, S.; Dykstra, M.J.; Lin, M.Y.; Kleven, S.H. Comparison of in Vivo and in Vitro Methods for Pathogenicity Evaluation for Mycoplasma gallisepticum in Respiratory Infection. Avian Pathol. 1986, 15, 233–246. [Google Scholar] [CrossRef]
- OIE Reference Laboratory Reports Activities. 2018. Available online: https://www.woah.org/fileadmin/Home/eng/Our_scientific_expertise/reflabreports/2018/report_689_2018_Glanders_UNITED_ARAB_EMIRATES.pdf (accessed on 12 February 2023).
- OIE Reference Laboratory Reports Activities. 2021. Available online: https://www.woah.org/fileadmin/Home/eng/Our_scientific_expertise/reflabreports/2021/report_1349_2021_Glanders_GERMANY.pdf (accessed on 12 February 2023).
- Brion, A. Aetiology and Control of Chronic Respiratory Disease. Br. Vet. J. 1961, 117, 296–305. [Google Scholar] [CrossRef]
- Hochachka, W.M.; Dhondt, A.A. Density-dependent decline of host abundance resulting from a new infectious disease. Proc. Natl. Acad. Sci. USA 2000, 97, 5303–5306. [Google Scholar] [CrossRef] [Green Version]
- Ley, D. Molecular Epidemiologic Investigations of Mycoplasma gallisepticum Conjunctivitis in Songbirds by Random Amplified Polymorphic DNA Analyses. Emerg. Infect. Dis. 1997, 3, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Hartup, B.K.; Kollias, G.V.; Ley, D.H. Mycoplasmal conjunctivitis in songbirds from New York. J. Wildl. Dis. 2000, 36, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Hartup, B.K.; Dhondt, A.A.; Sydenstricker, K.V.; Hochachka, W.M.; Kollias, G.V. Host range and dynamics of Mycoplasmal conjunctivitis among birds in north America. J. Wildl. Dis. 2001, 37, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, J.M.; Yavari, C.A.; Dare, C.M. Mycoplasmas and Respiratory Disease in Pheasants and Partridges. Avian Pathol. 2001, 30, 391–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benčina, D.; Mrzel, I.; ZormanRojs, O.; Bidovec, A.; Dovč, A. Characterisation of Mycoplasma gallisepticum Strains Involved in Respiratory Disease in Pheasants and Peafowl. Vet. Rec. 2003, 152, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Tiong, S. Isolation of Mycoplasma gallisepticum from Sinuses of Three Quails (Coturnix Coturnix Japonica). Vet. Rec. 1978, 103, 539. [Google Scholar] [CrossRef] [PubMed]
- Benčina, D.; Dorrer, D.; Tadina, T. Mycoplasma Species Isolated from Six Avian Species. Avian Pathol. 1987, 16, 653–664. [Google Scholar] [CrossRef]
- Bozeman, L.H.; Kleven, S.H.; Davis, R.B. Mycoplasma Challenge Studies in Budgerigars (Melopsittacus Undulatus) and Chickens. Avian Dis. 1984, 28, 426. [Google Scholar] [CrossRef]
- Poveda, J.B.; Carranza, J.; Miranda, A.; Garrido, A.; Hermoso, M.; Fernandez, A.; Domenech, J. An Epizootiological Study of Avian Mycoplasmas in Southern Spain. Avian Pathol. 1990, 19, 627–633. [Google Scholar] [CrossRef]
- Abdelrahman, A.A.; Shany, S.A.S.; Dardeer, M.A.A.; Hassan, K.E.; Ali, A.; El-Kady, M.F. Avian Mycoplasma gallisepticum and Mycoplasma synoviae: Advances in diagnosis and control. Ger.J. Vet. Res. 2021, 1, 46–55. [Google Scholar] [CrossRef]
- Roberts, D.H.; McDaniel, J.W. Mechanism of Egg Transmission of Mycoplasma gallisepticum. J. Comp. Pathol. 1967, 77, 439–442. [Google Scholar] [CrossRef]
- Armour, N.K.; Ferguson-Noel, N. Evaluation of the Egg Transmission and Pathogenicity of Mycoplasma gallisepticum Isolates Genotyped as Ts-11. Avian Pathol. 2015, 44, 296–304. [Google Scholar] [CrossRef]
- Jiang, Z.; Li, S.; Zhu, C.; Zhou, R.; Leung, P.H.M. Mycoplasma pneumoniae Infections: Pathogenesis and Vaccine Development. Pathogens 2021, 10, 119. [Google Scholar] [CrossRef]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Peebles, E.D. Occurrence of Horizontal Transmission in Layer Chickens after Administration of an in Ovo Strain F Mycoplasma gallisepticum Vaccine. Poult. Sci. 2019, 98, 4492–4497. [Google Scholar] [CrossRef]
- Dhondt, A.A.; Dhondt, K.V.; Hawley, D.M.; Jennelle, C.S. Experimental Evidence for Transmission of Mycoplasma gallisepticum in House Finches by Fomites. Avian Pathol. 2007, 36, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yu, S.; Hu, M.; Han, X.; Chen, D.; Qiu, X.; Ding, C. Identification of Biofilm Formation by Mycoplasma gallisepticum. Vet. Microbiol. 2012, 161, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Masukagami, Y.; Nijagal, B.; Tseng, C.-W.; Dayalan, S.; Tivendale, K.A.; Markham, P.F.; Browning, G.F.; Sansom, F.M. Metabolite Profiling of Mycoplasma gallisepticum Mutants, Combined with Bioinformatic Analysis, Can Reveal the Likely Functions of Virulence-Associated Genes. Vet. Microbiol. 2018, 223, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Papazisi, L.; Frasca, S.; Gladd, M.; Liao, X.; Yogev, D.; Geary, S.J. GapA and CrmACoexpression Is Essential for Mycoplasma gallisepticum Cytadherence and Virulence. Infect. Immun. 2002, 70, 6839–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunoya, T.; Kanai, K.; Yagihashi, T.; Hoshi, S.; Shibuya, K.; Tajima, M. Natural Case of Salpingitis Apparently Caused by Mycoplasma gallisepticum in Chickens. Avian Pathol. 1997, 26, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avakian, A.P.; Kleven, S.H. The Humoral Immune Response of Chickens to Mycoplasma gallisepticum and Mycoplasma Synoviae Studied by Immunoblotting. Vet. Microbiol. 1990, 24, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Gaunson, J.E.; Philip, C.J.; Whithear, K.G.; Browning, G.F. The Cellular Immune Response in the Tracheal Mucosa to Mycoplasma gallisepticum in Vaccinated and Unvaccinated Chickens in the Acute and Chronic Stages of Disease. Vaccine 2006, 24, 2627–2633. [Google Scholar] [CrossRef]
- Javed, M.A.; Frasca, S.; Rood, D.; Cecchini, K.; Gladd, M.; Geary, S.J.; Silbart, L.K. Correlates of Immune Protection in Chickens Vaccinated with Mycoplasma gallisepticum Strain GT5 Following Challenge with Pathogenic M. Gallisepticum Strain R low. Infect. Immun. 2005, 73, 5410–5419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaunson, J.E.; Philip, C.J.; Whithear, K.G.; Browning, G.F. Lymphocytic Infiltration in the Chicken Trachea in Response to Mycoplasma gallisepticum Infection. Microbiology 2000, 146, 1223–1229. [Google Scholar] [CrossRef] [Green Version]
- Whithear, K.G. Control of Avian Mycoplasmoses by Vaccination: -EN- -FR- -ES-. Rev. Sci. Tech. OIE 1996, 15, 1527–1553. [Google Scholar] [CrossRef] [PubMed]
- Horizon Bioscience. Mycoplasmas: Molecular Biology, Pathogenicity and Strategies for Control; Blanchard, A., Browning, G., Eds.; Horizon Bioscience: Wymondham, UK; Norfolk, VA, USA, 2005. [Google Scholar]
- Majumder, S.; Zappulla, F.; Silbart, L.K. Mycoplasma gallisepticum Lipid Associated Membrane Proteins Up-Regulate Inflammatory Genes in Chicken Tracheal Epithelial Cells via TLR-2 Ligation through an NF-ΚB Dependent Pathway. PLoS ONE 2014, 9, e112796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matucci, A.; Stefani, E.; Gastaldelli, M.; Rossi, I.; De Grandi, G.; Gyuranecz, M.; Catania, S. Molecular differentiation of Mycoplasma gallisepticum outbreaks: A last decade study on Italian farms using GTS and MLST. Vaccines 2020, 8, 665. [Google Scholar] [CrossRef] [PubMed]
- Limsatanun, A.; Pakpinyo, S.; Limpavithayakul, K.; Prasertsee, T. Targeted sequencing analysis of Mycoplasma gallisepticum isolates in chicken layer and breeder flocks in Thailand. Sci. Rep. 2022, 12, 9900. [Google Scholar] [CrossRef] [PubMed]
- Indiková, I.; Much, P.; Stipkovits, L.; Siebert-Gulle, K.; Szostak, M.P.; Rosengarten, R.; Citti, C. Role of the GapA and CrmACytadhesins of Mycoplasma gallisepticum in Promoting Virulence and Host Colonization. Infect. Immun. 2013, 81, 1618–1624. [Google Scholar] [CrossRef] [Green Version]
- May, M.; Papazisi, L.; Gorton, T.S.; Geary, S.J. Identification of Fibronectin-Binding Proteins in Mycoplasma gallisepticum Strain R. Infect. Immun. 2006, 74, 1777–1785. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.; Geary, S.J.; Gladd, M.; Djordjevic, S.P. The Mycoplasma gallisepticum OsmC-like Protein MG1142 Resides on the Cell Surface and Binds Heparin. Microbiology 2007, 153, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Pflaum, K.; Tulman, E.R.; Beaudet, J.; Canter, J.; Geary, S.J. Variable Lipoprotein Hemagglutinin A Gene ( VlhA ) Expression in Variant Mycoplasma gallisepticum Strains In Vivo. Infect. Immun. 2018, 86, e00524-18. [Google Scholar] [CrossRef] [Green Version]
- Boguslavsky, S.; Menaker, D.; Lysnyansky, I.; Liu, T.; Levisohn, S.; Rosengarten, R.; García, M.; Yogev, D. Molecular Characterization of the Mycoplasma gallisepticum PvpA Gene Which Encodes a Putative Variable Cytadhesin Protein. Infect. Immun. 2000, 68, 3956–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Zhang, L.; Chen, Y.; Li, Y.; Wang, Z.; Li, G.; Wang, G.; Xin, J. GroEL Protein (Heat Shock Protein 60) of Mycoplasma gallisepticum Induces Apoptosis in Host Cells by Interacting with Annexin A2. Infect. Immun. 2019, 87, e00248-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Zhang, F.; Wang, Y.; Liu, T.; Tan, L.; Wang, S.; Tian, M.; Li, T.; Wang, X.; Ding, C.; et al. Characterization of Mycoplasma gallisepticum Pyruvate Dehydrogenase Alpha and Beta Subunits and Their Roles in Cytoadherence. PLoS ONE 2018, 13, e0208745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudahi-Orenstein, S.; Levisohn, S.; Geary, S.J.; Yogev, D. Cytadherence-Deficient Mutants of Mycoplasma gallisepticum Generated by Transposon Mutagenesis. Infect. Immun. 2003, 71, 3812–3820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, G.F.; Marenda, M.S.; Noormohammadi, A.H.; Markham, P.F. The Central Role of Lipoproteins in the Pathogenesis of Mycoplasmoses. Vet. Microbiol. 2011, 153, 44–50. [Google Scholar] [CrossRef]
- Seto, S.; Kenri, T.; Tomiyama, T.; Miyata, M. Involvement of P1 Adhesin in Gliding Motility of Mycoplasma Pneumoniae as Revealed by the Inhibitory Effects of Antibody under Optimized Gliding Conditions. J. Bacteriol. 2005, 187, 1875–1877. [Google Scholar] [CrossRef] [Green Version]
- Noormohammadi, A.H. Role of Phenotypic Diversity in Pathogenesis of Avian Mycoplasmosis. Avian Pathol. 2007, 36, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Glew, M.D.; Browning, G.F.; Markham, P.F.; Walker, I.D. PMGA Phenotypic Variation in Mycoplasma gallisepticum Occurs In Vivo and Is Mediated by Trinucleotide Repeat Length Variation. Infect. Immun. 2000, 68, 6027–6033. [Google Scholar] [CrossRef] [Green Version]
- Chopra-Dewasthaly, R.; Spergser, J.; Zimmermann, M.; Citti, C.; Jechlinger, W.; Rosengarten, R. Vpma Phase Variation Is Important for Survival and Persistence of Mycoplasma Agalactiae in the Immunocompetent Host. PLoS Pathog. 2017, 13, e1006656. [Google Scholar] [CrossRef] [Green Version]
- Czurda, S.; Hegde, S.M.; Rosengarten, R.; Chopra-Dewasthaly, R. Xer1-Independent Mechanisms of Vpma Phase Variation in Mycoplasma Agalactiae Are Triggered by Vpma-Specific Antibodies. Int. J. Med. Microbiol. 2017, 307, 443–451. [Google Scholar] [CrossRef]
- Ma, L.; Jensen, J.S.; Mancuso, M.; Myers, L.; Martin, D.H. Kinetics of Genetic Variation of the Mycoplasma Genitalium MG192 Gene in Experimentally Infected Chimpanzees. Infect. Immun. 2016, 84, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citti, C.; Nouvel, L.-X.; Baranowski, E. Phase and Antigenic Variation in Mycoplasmas. Future Microbiol. 2010, 5, 1073–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, F.; Zhao, C.; Bi, D.; Tian, W.; Chen, J.; Sun, J.; Peng, X. Mycoplasma gallisepticum (HS Strain) Surface Lipoprotein PMGA Interacts with Host Apolipoprotein A-I during Infection in Chicken. Appl. MicrobiolBiotechnol. 2016, 100, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Vogl, G.; Plaickner, A.; Szathmary, S.; Stipkovits, L.; Rosengarten, R.; Szostak, M.P. Mycoplasma gallisepticum Invades Chicken Erythrocytes during Infection. Infect. Immun. 2008, 76, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosengarten, R.; Behrens, A.; Stetefeld, A.; Heller, M.; Ahrens, M.; Sachse, K.; Yogev, D.; Kirchhoff, H. Antigen Heterogeneity among Isolates of Mycoplasma Bovis Is Generated by High-Frequency Variation of Diverse Membrane Surface Proteins. Infect. Immun. 1994, 62, 5066–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosengarten, R.; Yogev, D. Variant Colony Surface Antigenic Phenotypes within Mycoplasma Strain Populations: Implications for Species Identification and Strain Standardization. J. Clin. Microbiol. 1996, 34, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Yogev, D.; Menaker, D.; Strutzberg, K.; Levisohn, S.; Kirchhoff, H.; Hinz, K.H.; Rosengarten, R. A Surface Epitope Undergoing High-Frequency Phase Variation Is Shared by Mycoplasma gallisepticum and Mycoplasma Bovis. Infect. Immun. 1994, 62, 4962–4968. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, S.H.E. Heat Shock Proteins and the Immune Response. Immunol. Today 1990, 11, 129–136. [Google Scholar] [CrossRef]
- Purswell, J.L.; Evans, J.D.; Branton, S.L. Serologic Response of Roosters to Gradient Dosage Levels of a Commercially Available Live F Strain-Derived Mycoplasma gallisepticum Vaccine Over Time. Avian Dis. 2011, 55, 490–494. [Google Scholar] [CrossRef]
- Michiels, T.; Welby, S.; Vanrobaeys, M.; Quinet, C.; Rouffaer, L.; Lens, L.; Martel, A.; Butaye, P. Prevalence of Mycoplasma gallisepticum and Mycoplasma Synoviae in Commercial Poultry, Racing Pigeons and Wild Birds in Belgium. Avian Pathol. 2016, 45, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Gazzar, E.I.; Laibinis, M.; Ferguson, N. Characterization of a Ts-11–like Mycoplasma gallisepticum Isolate from Commercial Broiler Chickens. Avian Dis. 2011, 55, 569–574. [Google Scholar] [CrossRef] [PubMed]
- de Jong, A.; Youala, M.; Klein, U.; El Garch, F.; Simjee, S.; Moyaert, H.; Rose, M.; Gautier-Bouchardon, A.V.; Catania, S.; Ganapathy, K.; et al. Minimal Inhibitory Concentration of Seven Antimicrobials to Mycoplasma gallisepticum and Mycoplasma Synoviae Isolates from Six European Countries. Avian Pathol. 2021, 50, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Silley, P.; Simjee, S.; Schwarz, S. Surveillance and Monitoring of Antimicrobial Resistance and Antibiotic Consumption in Humans and Animals: -EN- -FR- Surveillance et Contrôle de La Résistance Aux Antimicrobiens et de La Consommationd’antibiotiquesEnMédecineHumaine et Vétérinaire -ES- Vigilancia y Seguimiento de La Resistencia a Los Antimicrobianos y El Consumo de AntibióticosEn Personas y Animales. Rev. Sci. Tech. OIE 2012, 31, 105–120. [Google Scholar] [CrossRef]
- Ferguson-Noel, N.; Cookson, K.; Laibinis, V.A.; Kleven, S.H. The Efficacy of Three Commercial Mycoplasma gallisepticum Vaccines in Laying Hens. Avian Dis. 2012, 56, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Ishfaq, M.; Hu, W.; Khan, M.Z.; Ahmad, I.; Guo, W.; Li, J. Current Status of Vaccine Research, Development, and Challenges of Vaccines for Mycoplasma gallisepticum. Poult. Sci. 2020, 99, 4195–4202. [Google Scholar] [CrossRef]
- Redweik, G.A.J.; Jochum, J.; Mellata, M. Live Bacterial Prophylactics in Modern Poultry. Front. Vet. Sci. 2020, 7, 592312. [Google Scholar] [CrossRef]
- Vaxxinova. “Poultry Vaccines”. Available online: https://vaxxinova.com/poultry-vaccines/ (accessed on 23 March 2022).
- Jacob, R.; Branton, S.L.; Evans, J.D.; Leigh, S.A.; Peebles, E.D. Effects of Live and Killed Vaccines against Mycoplasma gallisepticum on the Performance Characteristics of Commercial Layer Chickens. Poult. Sci. 2014, 93, 1403–1409. [Google Scholar] [CrossRef]
- Kleven, S.H. Control of Avian Mycoplasma Infections in Commercial Poultry. Avian Dis. 2008, 52, 367–374. [Google Scholar] [CrossRef]
- Cummings, T.S.; Kleven, S.H. Evaluation of Protection against Mycoplasma gallisepticum Infection in Chickens Vaccinated with the F Strain of M. Gallisepticum. Avian Dis. 1986, 30, 169. [Google Scholar] [CrossRef]
- Machado, L.d.S.; Santos, F.F.d.; Togashi, C.K.; Abreu, D.L.d.C.; Pimentel, J.C.; Sesti, L.; Pereira, V.L.d.A.; Nascimento, E.R.d. Egg Quality in Laying Hens Exposed to Mycoplasma gallisepticum F-Strain Attenuated Vaccine. Poult. Sci. 2017, 96, 976–979. [Google Scholar] [CrossRef]
- Rodriguez, R.; Kleven, S.H. Pathogenicity of Two Strains of Mycoplasma gallisepticum in Broilers. Avian Dis. 1980, 24, 800. [Google Scholar] [CrossRef] [PubMed]
- Gazzar, E.I. Mycoplasma gallisepticum Infection in Poultry. 2022. Available online: https://www.msdvetmanual.com/poultry/mycoplasmosis/mycoplasma-gallisepticum-infection-in-poultry (accessed on 10 February 2023).
- Abd-El-Motelib, T.Y.; Kleven, S.H. A Comparative Study of Mycoplasma gallisepticum Vaccines in Young Chickens. Avian Dis. 1993, 37, 981. [Google Scholar] [CrossRef]
- Whithear, K.; Soeripto; Harrigan, K.; Ghiocas, E. Immunogenicity of a Temperature Sensitive Mutant Mycoplasma Gaffisepticum Vaccine. Aust. Vet. J. 1990, 67, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, A.H.; Jones, J.F.; Underwood, G.; Whithear, K.G. Poor Systemic Antibody Response After Vaccination of Commercial Broiler Breeders with Mycoplasma gallisepticum Vaccine Ts-11 Not Associated with Susceptibility to Challenge. Avian Dis. 2002, 46, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, A.H.; Browning, G.F.; Cowling, P.J.; O’Rourke, D.; Whithear, K.G.; Markham, P.F. Detection of Antibodies to Mycoplasma gallisepticum Vaccine Ts-11 by an Autologous PMGA Enzyme-Linked Immunosorbent Assay. Avian Dis. 2002, 46, 405–411. [Google Scholar] [CrossRef]
- Gates, A.E.; Frasca, S.; Nyaoke, A.; Gorton, T.S.; Silbart, L.K.; Geary, S.J. Comparative Assessment of a Metabolically Attenuated Mycoplasma gallisepticum Mutant as a Live Vaccine for the Prevention of Avian Respiratory Mycoplasmosis. Vaccine 2008, 26, 2010–2019. [Google Scholar] [CrossRef]
- Kanci, A.; Wijesurendra, D.S.; Wawegama, N.K.; Underwood, G.J.; Noormohammadi, A.H.; Markham, P.F.; Browning, G.F. Evaluation of Mycoplasma gallisepticum (MG) Ts-304 Vaccine as a Live Attenuated Vaccine in Turkeys. Vaccine 2018, 36, 2487–2493. [Google Scholar] [CrossRef]
- Hudson, P.; Gorton, T.S.; Papazisi, L.; Cecchini, K.; Frasca, S.; Geary, S.J. Identification of a Virulence-Associated Determinant, Dihydrolipoamide Dehydrogenase (Lpd), in Mycoplasma gallisepticum through In Vivo Screening of Transposon Mutants. Infect. Immun. 2006, 74, 931–939. [Google Scholar] [CrossRef] [Green Version]
- Ferguson-Noel, N.M.; Laibinis, V.A.; Kleven, S.H. Evaluation of Mycoplasma gallisepticum K-Strain as a Live Vaccine in Chickens. Avian Dis. 2012, 56, 44–50. [Google Scholar] [CrossRef]
- Ferguson-Noel, N.M.; Williams, S.M. The Efficacy of Mycoplasma gallisepticum K-Strain Live Vaccine in Broiler and Layer Chickens. Avian Pathol. 2015, 44, 75–80. [Google Scholar] [CrossRef]
- Kanci Condello, A.; KulappuArachchige, S.N.; Shil, P.K.; Underwood, G.J.; Noormohammadi, A.H.; Markham, P.F.; Wawegama, N.K.; Browning, G.F. Duration of Protective Immunity Induced by Mycoplasma gallisepticum Strain Ts-304 Vaccine in Chickens. Vet. Microbiol. 2020, 251, 108883. [Google Scholar] [CrossRef]
- Zhang, G.Z.; Zhang, R.; Zhao, H.L.; Wang, X.T.; Zhang, S.P.; Li, X.J.; Qin, C.Z.; Lv, C.M.; Zhao, J.X.; Zhou, J.F. A Safety Assessment of a Fowlpox-Vectored Mycoplasma gallisepticum Vaccine in Chickens. Poult. Sci. 2010, 89, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. Multi-Epitope Vaccines: A Promising Strategy against Tumors and Viral Infections. Cell Mol. Immunol. 2018, 15, 182–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudtson, K.L.; Minion, F.C. Construction of Tn4001 Lac Derivatives to Be Used as Promoter Probe Vectors in Mycoplasmas. Gene 1993, 137, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Dybvig, K.; French, C.T.; Voelker, L.L. Construction and Use of Derivatives of Transposon Tn 4001 That Function in Mycoplasma Pulmonis and Mycoplasma Arthritidis. J. Bacteriol. 2000, 182, 4343–4347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.; Ghosh, P.P.; Azim, K.F.; Mukta, S.; Abir, R.A.; Nahar, J.; Hasan Khan, M.M. Reverse Vaccinology Approach to Design a Novel Multi-Epitope Subunit Vaccine against Avian Influenza A (H7N9) Virus. Microb. Pathog. 2019, 130, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Madlala, T.; Adeleke, V.T.; Fatoba, A.J.; Okpeku, M.; Adeniyi, A.A.; Adeleke, M.A. Designing Multiepitope-Based Vaccine against Eimeria from Immune Mapped Protein 1 (IMP-1) Antigen Using Immunoinformatic Approach. Sci. Rep. 2021, 11, 18295. [Google Scholar] [CrossRef]
- Mugunthan, S.P.; Harish, M.C. Multi-Epitope-Based Vaccine Designed by Targeting Cytoadherence Proteins of Mycoplasma gallisepticum. ACS Omega 2021, 6, 13742–13755. [Google Scholar] [CrossRef]
- Mugunthan, S.P.; Mani Chandra, H. A Computational Reverse Vaccinology Approach for the Design and Development of Multi-Epitopic Vaccine Against Avian Pathogen Mycoplasma gallisepticum. Front. Vet. Sci. 2021, 8, 721061. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant Protein Expression in Escherichia Coli: Advances and Challenges. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Shim, B.-S.; Hong, K.-J.; Maharjan, P.M.; Choe, S. Plant Factory: New Resource for the Productivity and Diversity of Human and Veterinary Vaccines. Clin Exp. Vaccine Res. 2019, 8, 136. [Google Scholar] [CrossRef]
- Elyazeed, H.A.; Al-Atfeehy, N.M.; Abotaleb, R.; Sayed, R.; Marouf, S. Preparation of ELISA and Lateral Flow Kits for Rapid Diagnosis of Mycoplasma gallisepticum in Poultry. Sci. Rep. 2020, 10, 9056. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Pingali, M.S.; Paital, B.; Panda, F.; Pati, S.G.; Singh, A.; Varadwaj, P.K.; Samanta, S.K. A detailed review of the outbreak of COVID-19. Front. Biosci. (Landmark Ed) 2021, 26, 149–170. [Google Scholar] [CrossRef] [PubMed]
- Shivalkar, S.; Pingali, M.S.; Verma, A.; Singh, A.; Singh, V.; Paital, B.; Das, D.; Varadwaj, P.K.; Samanta, S.K. Outbreak of COVID-19: A Detailed Overview and Its Consequences. Adv. Exp. Med. Biol. 2021, 1353, 23–45. [Google Scholar] [CrossRef] [PubMed]
- WHO. Vaccines and immunization. Available online: https://www.who.int/health-topics/vaccines-and-immunization#tab=tab_1 (accessed on 12 February 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Protein | Gene Annotation | Reference |
|---|---|---|---|
| Adhesion | GapA | M. gallisepticum A_0934 | [29,40] |
| CrmA | M. gallisepticum A_0939 | [29,40] | |
| Hlp3 | M. gallisepticum A_0928 | [41] | |
| PlpA | M. gallisepticum A_1199 | [41] | |
| OsmC-like protein | M. gallisepticum A_1142 | [42] | |
| Immune evasion | vlhA 1.04 | M. gallisepticum A_0070 | [43] |
| vlhA 2.02 | M. gallisepticum A_0117 | [43] | |
| vlhA 3.03 | M. gallisepticum A_0380 | [43] | |
| vlhA 4.07 | M. gallisepticum A_0977 | [43] | |
| vlhA 5.13 | M. gallisepticum A_1261 | [43] | |
| Phase variation | PvpA | M. gallisepticum A_0258 | [44] |
| Heat shock proteins | GroEl | M. gallisepticum A_0152 | [45] |
| Strain | Name | Manufacturer |
|---|---|---|
| Strain F | Mycoplasma gallisepticum vaccine | Shandong Lvdu Biosciences [68] |
| CEVAC MG F | CevaSanteAnimale [68] | |
| PoulvacMyco F | Zoetis United States [68] | |
| AviPro® MG-F | Elanco [69] | |
| Strain K | VAXXON® MG Live | Vaxxinova® International BV [70] |
| Strain ts-11 | VAXSAFE MG VACCINE (MG TS-11) | Bioproperties Pty LTD [68] |
| Strain 6/85 | Nobilis MG 6/85 | MSD Animal Health [68] |
| MYCOVAC-L® | Merck [69] | |
| Strain S6 | VAXXON® MG Inac | Vaxxinova® International BV [70] |
| Strain R | MG-Bac Vaccine | Zoetis United States [68] |
| AviPro104 MG BACTERIN | Lohmann Animal Health International [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugunthan, S.P.; Kannan, G.; Chandra, H.M.; Paital, B. Infection, Transmission, Pathogenesis and Vaccine Development against Mycoplasma gallisepticum. Vaccines 2023, 11, 469. https://doi.org/10.3390/vaccines11020469
Mugunthan SP, Kannan G, Chandra HM, Paital B. Infection, Transmission, Pathogenesis and Vaccine Development against Mycoplasma gallisepticum. Vaccines. 2023; 11(2):469. https://doi.org/10.3390/vaccines11020469
Chicago/Turabian StyleMugunthan, Susithra Priyadarshni, Ganapathy Kannan, Harish Mani Chandra, and Biswaranjan Paital. 2023. "Infection, Transmission, Pathogenesis and Vaccine Development against Mycoplasma gallisepticum" Vaccines 11, no. 2: 469. https://doi.org/10.3390/vaccines11020469