
Identification and Evaluation of Novel Antigen Candidates against Salmonella Pullorum Infection Using Reverse Vaccinology

Abstract
:1. Introduction
2. Materials and Methods
2.1. S. Pullorum Strain R51 Isolation and Identification
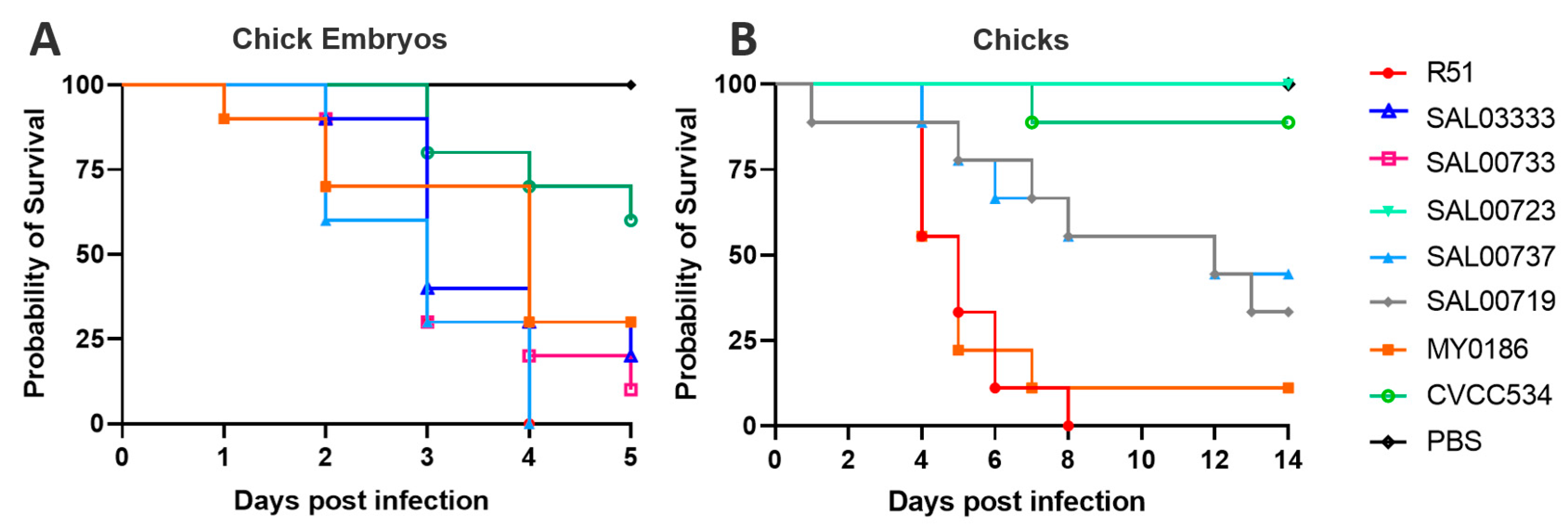
2.2. Virulence Assays in Selecting the Candidate Strain
2.3. Whole Genomic Sequencing and Additional Clinical Collections
2.4. Selection of Essential Proteins
2.5. Identification of the Signal Peptide
2.6. Antigenicity Prediction
2.7. Evaluation of Adhesin Probability
2.8. Prediction of Transmembrane Domains and Solubility
2.9. Protein Sequence Conservation Analysis
2.10. Bacterial Strains and Growth Conditions
2.11. Gene Cloning, Protein Expression, and Purification
2.12. In Ovo Vaccination with Candidate Proteins
2.13. Immunogenicity Assay
2.14. Bacterial Loads after Challenge
2.15. Immune Protection Assessment
2.16. Statistical Analyses
3. Results
3.1. Virulence Assay for the Selection of the Candidate Strain
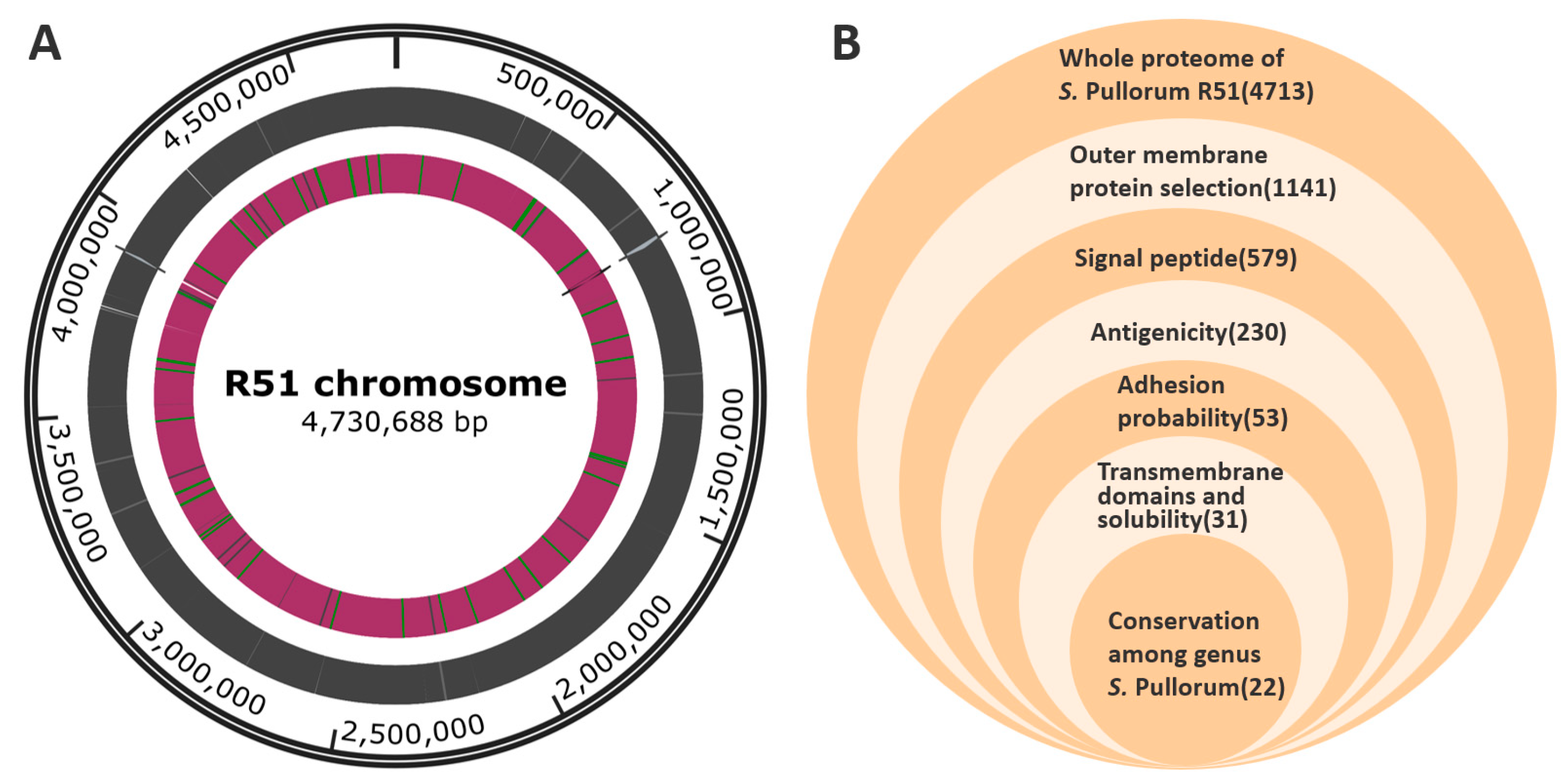
3.2. Complete Genome and Proteome of R51
3.3. OMP and Extracellular Protein Preselection
3.4. Identification of the Signal Peptide
3.5. Antigenicity Prediction
3.6. Evaluation of Adhesive Probability
3.7. Prediction of Transmembrane Domains and Solubility
3.8. Comparative Analysis
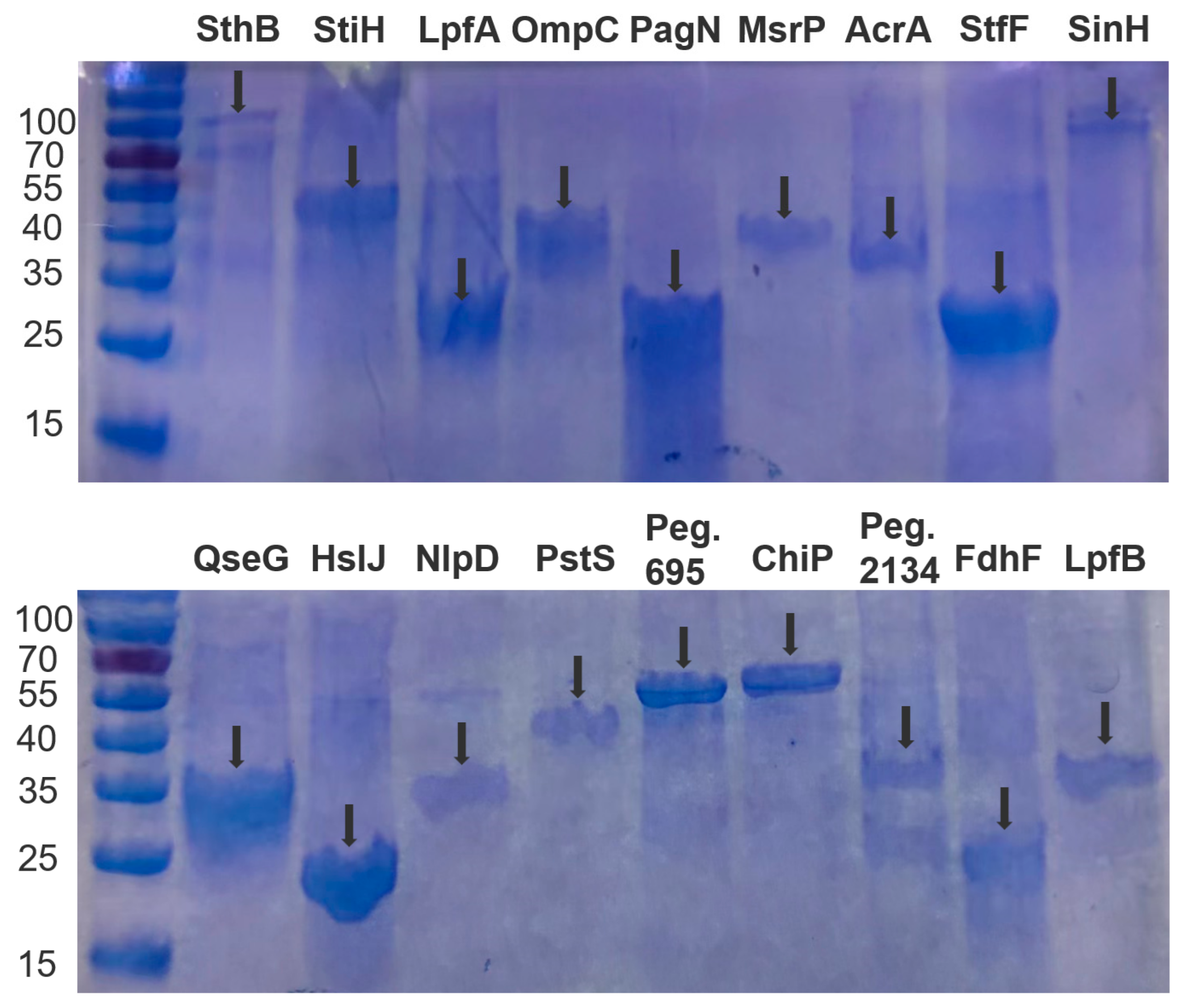
3.9. Protein Expression and Purification
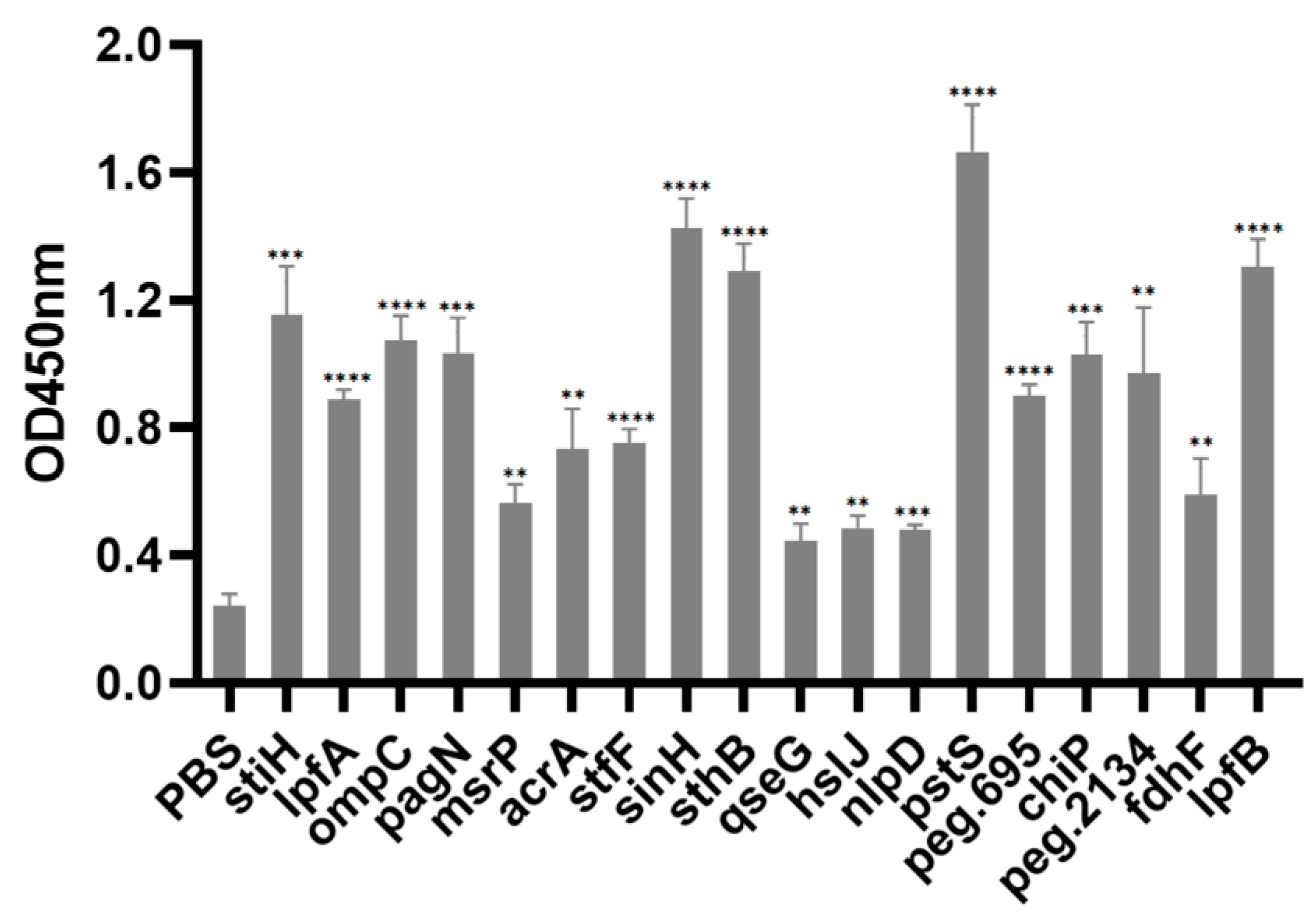
3.10. Immunogenicity Assay
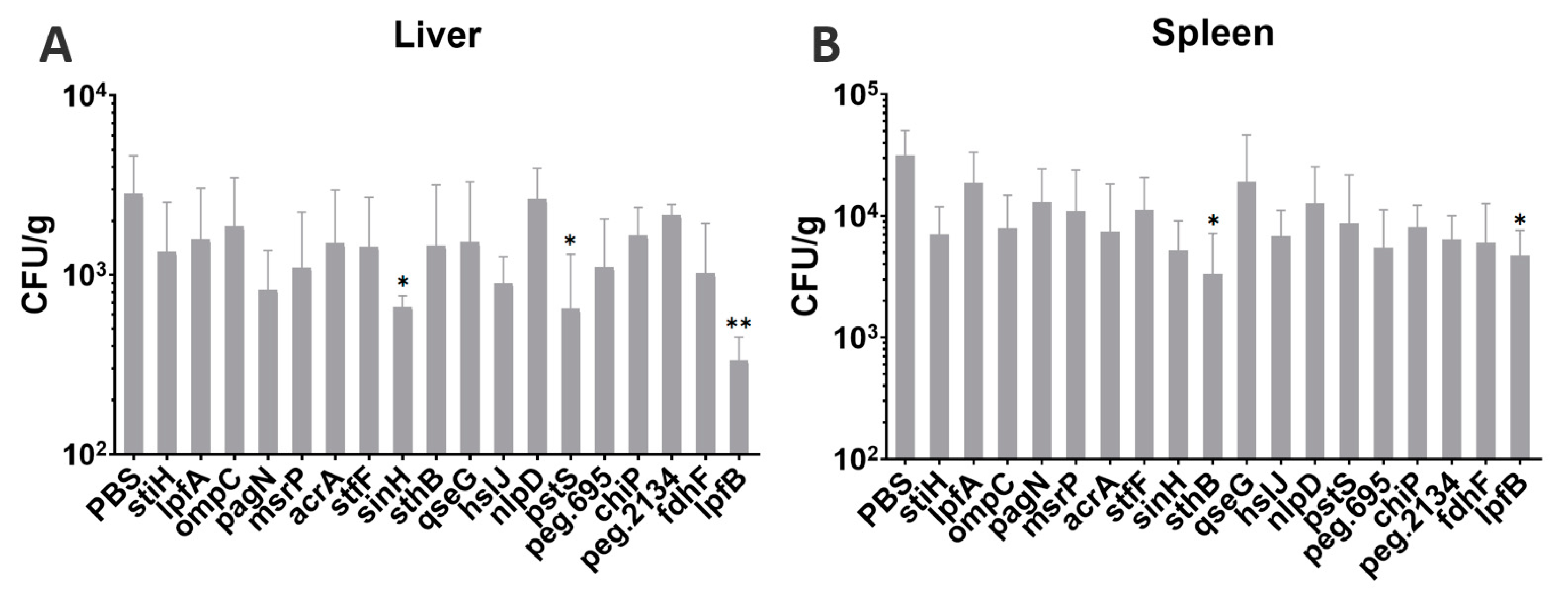
3.11. Bacterial Loads after Challenge
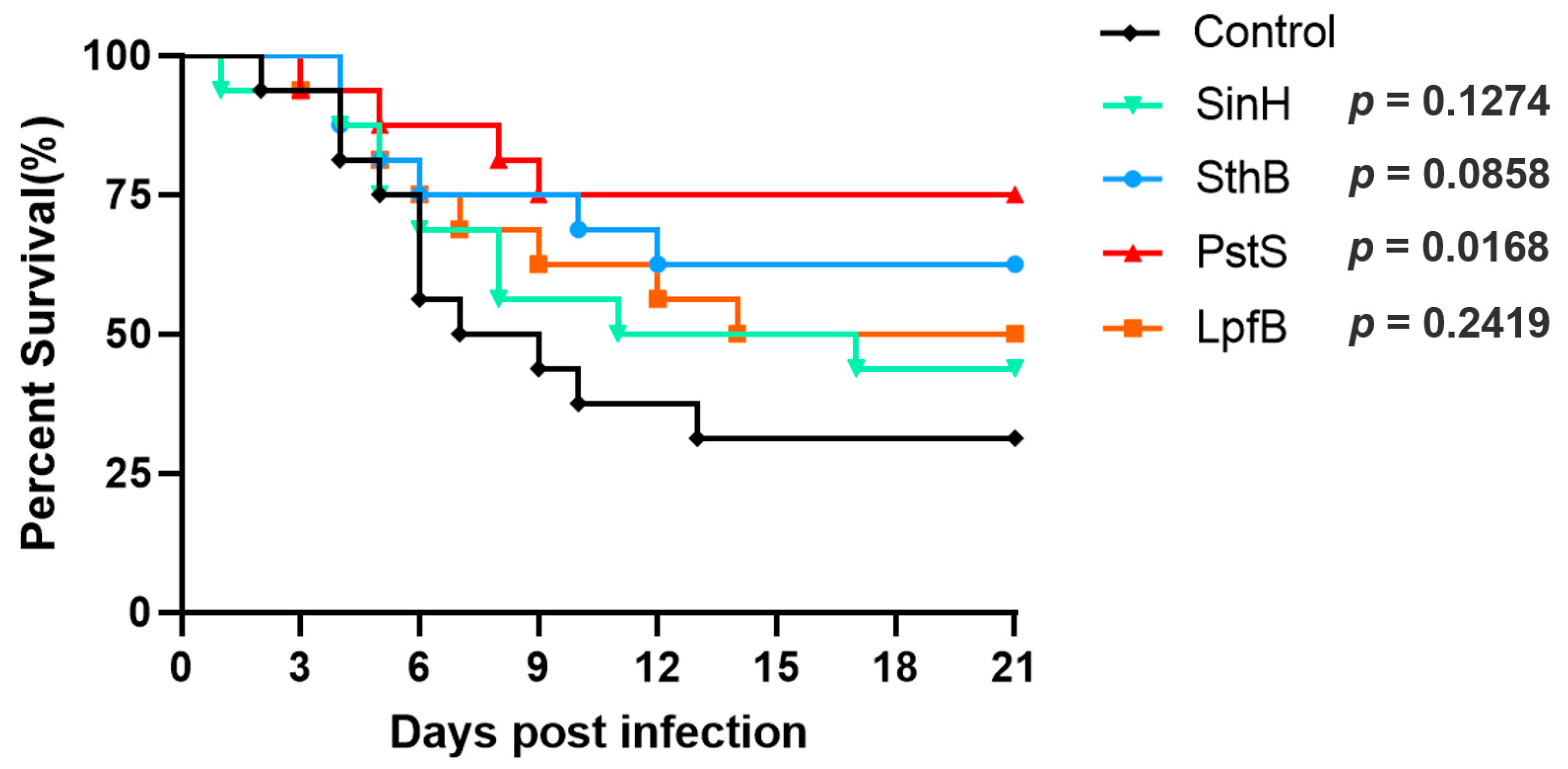
3.12. Evaluation of Protective Immunity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, X.; Kang, X.; Zhou, K.; Yue, M. A global dataset for prevalence of Salmonella Gallinarum between 1945 and 2021. Sci. Data 2022, 9, 495. [Google Scholar] [CrossRef]
- Li, Y.; Kang, X.; Ed-Dra, A.; Zhou, X.; Jia, C.; Müller, A.; Liu, Y.; Kehrenberg, C.; Yue, M. Genome-Based Assessment of Antimicrobial Resistance and Virulence Potential of Isolates of Non-Pullorum/Gallinarum Salmonella Serovars Recovered from Dead Poultry in China. Microbiol. Spectr. 2022, 10, e0096522. [Google Scholar] [CrossRef]
- Barrow, P.A.; Neto, O.C.F. Pullorum disease and fowl typhoid—New thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H. Fowl typhoid and pullorum disease. Rev. Sci. Tech. 2000, 19, 405–424. [Google Scholar] [CrossRef]
- Snoeyenbos, G.H.; Smyser, C.F.; Van Roekel, H. Research Note: Salmonella Infections of the Ovary and Peritoneum of Chickens. Avian Dis. 1969, 13, 668. [Google Scholar] [CrossRef]
- Wigley, P.; Berchieri, A., Jr.; Page, K.L.; Smith, A.L.; Barrow, P.A. Salmonella enterica serovar Pullorum persists in splenic macrophages and in the reproductive tract during persistent, disease-free carriage in chickens. Infect. Immun. 2001, 69, 7873–7879. [Google Scholar] [CrossRef]
- Kang, X.; Zhou, X.; Tang, Y.; Jiang, Z.; Chen, J.; Mohsin, M.; Yue, M. Characterization of Two-Component System CitB Family in Salmonella Pullorum. Int. J. Mol. Sci. 2022, 23, 10201. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, X.; Tang, Y.; Jiang, Z.; Kang, X.; Wang, J.; Yue, M. Characterization of two-component system CitB family in Salmonella enterica serovar Gallinarum biovar Gallinarum. Vet. Microbiol. 2023, 278, 109659. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, A.J.; Hargis, B.M.; Tsolis, R.M. Tracing the Origins of Salmonella Outbreaks. Science 2000, 287, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Jia, C.; Paudyal, N.; Li, F.; Mao, J.; Liu, X.; Dong, C.; Zhou, K.; Liao, X.; Gong, J.; et al. Comprehensive Assessment of Subtyping Methods for Improved Surveillance of Foodborne Salmonella. Microbiol. Spectr. 2022, 10, e0247922. [Google Scholar] [CrossRef]
- Tang, B.; Siddique, A.; Jia, C.; Ed-Dra, A.; Wu, J.; Lin, H.; Yue, M. Genome-based risk assessment for foodborne Salmonella enterica from food animals in China: A One Health perspective. Int. J. Food Microbiol. 2023, 390, 110120. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ed-Dra, A.; Zhou, H.; Wu, B.; Zhang, Y.; Yue, M. Antimicrobial resistance and genomic investigation of non-typhoidal Salmonella isolated from outpatients in Shaoxing city, China. Front. Public Health 2022, 10, 988317. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Teng, L.; Xu, X.; Li, X.; Peng, X.; Zhou, X.; Du, J.; Tang, Y.; Jiang, Z.; Wang, Z.; et al. A nontyphoidal Salmonella serovar domestication accompanying enhanced niche adaptation. EMBO Mol. Med. 2022, 14, e16366. [Google Scholar] [CrossRef]
- Tang, B.; Elbediwi, M.; Nambiar, R.B.; Yang, H.; Lin, J.; Yue, M. Genomic Characterization of Antimicrobial-Resistant Salmonella enterica in Duck, Chicken, and Pig Farms and Retail Markets in Eastern China. Microbiol. Spectr. 2022, 10, e0125722. [Google Scholar] [CrossRef] [PubMed]
- Elbediwi, M.; Tang, Y.; Yue, M. Genomic characterization of ESBL-producing Salmonella Thompson isolates harboring mcr-9 from dead chick embryos in China. Vet. Microbiol. 2023, 278, 109634. [Google Scholar] [CrossRef]
- Teng, L.; Liao, S.; Zhou, X.; Jia, C.; Feng, M.; Pan, H.; Ma, Z.; Yue, M. Prevalence and Genomic Investigation of Multidrug-Resistant Salmonella Isolates from Companion Animals in Hangzhou, China. Antibiotics 2022, 11, 625. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, X.; Jiang, Z.; Qi, Y.; Ed-Dra, A.; Yue, M. Antimicrobial Resistance Profiles and Genetic Typing of Salmonella Serovars from Chicken Embryos in China. Antibiotics 2021, 10, 1156. [Google Scholar] [CrossRef]
- Elbediwi, M.; Tang, Y.; Shi, D.; Ramadan, H.; Xu, Y.; Xu, S.; Li, Y.; Yue, M. Genomic Investigation of Antimicrobial-Resistant Salmonella enterica Isolates from Dead Chick Embryos in China. Front. Microbiol. 2021, 12, 684400. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, J.; Ed-Dra, A.; Li, X.; Peng, X.; Xia, L.; Guo, Q.; Yao, G.; Yue, M. Prevalence and genomic investigation of Salmonella isolates recovered from animal food-chain in Xinjiang, China. Food Res. Int. 2021, 142, 110198. [Google Scholar] [CrossRef]
- Design, T.S.; Koster, W.; Potter, A. Salmonella vaccines in poultry: Past, present and future. Expert. Rev. Vaccines 2013, 12, 87–96. [Google Scholar] [CrossRef]
- Silva, E.N.; Snoeyenbos, G.H.; Weinack, O.M.; Smyser, C.F. Studies on the Use of 9R Strain of Salmonella gallinarum as a Vaccine in Chickens. Avian Dis. 1981, 25, 38. [Google Scholar] [CrossRef]
- Woodward, M.J.; Gettinby, G.; Breslin, M.F.; Corkish, J.D.; Houghton, S. The efficacy of Salenvac, a Salmonella enterica subsp. Enterica serotype Enteritidis iron-restricted bacterin vaccine, in laying chickens. Avian Pathol. 2002, 31, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K. Serotype-Specific and Serotype-Independent Strategies for Preharvest Control of Food-Borne Salmonella in Poultry. Avian Dis. 2007, 51, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A. Salmonella infections: Immune and non-immune protection with vaccines. Avian Pathol. 2007, 36, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G. Bacterial surface proteins and vaccines. F1000 Biol. Rep. 2010, 2, 777–780. [Google Scholar] [CrossRef]
- Yue, M.; Rankin, S.C.; Blanchet, R.T.; Nulton, J.D.; Edwards, R.; Schifferli, D.M. Diversification of the Salmonella Fimbriae: A Model of Macro- and Microevolution. PLoS ONE 2012, 7, e38596. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Han, X.; De Masi, L.; Zhu, C.; Ma, X.; Zhang, J.; Wu, R.; Schmieder, R.; Kaushik, R.S.; Fraser, G.P.; et al. Allelic variation contributes to bacterial host specificity. Nat. Commun. 2015, 6, 8754. [Google Scholar] [CrossRef]
- De Masi, L.; Yue, M.; Hu, C.; Rakov, A.V.; Rankin, S.C.; Schifferli, D.M. Cooperation of Adhesin Alleles in Salmonella -Host Tropism. mSphere 2017, 2, e00066-17. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.J.; Jones, C.; Blohmke, C.J.; Darton, T.C.; Goudet, A.; Sergeant, R.; Maillere, B.; Pollard, A.J.; Altmann, D.M.; Boyton, R.J. The serodominant secreted effector protein of Salmonella, SseB, is a strong CD4 antigen containing an immunodominant epitope presented by diverse HLA class II alleles. Immunology 2014, 143, 438–446. [Google Scholar] [CrossRef]
- Toyota-Hanatani, Y.; Kyoumoto, Y.; Baba, E.; Ekawa, T.; Ohta, H.; Tani, H.; Sasai, K. Importance of subunit vaccine antigen of major FliC antigenic site of Salmonella Enteritidis II: A challenge trial. Vaccine 2009, 27, 1680–1684. [Google Scholar] [CrossRef] [PubMed]
- Gil-Cruz, C.; Bobat, S.; Marshall, J.L.; Kingsley, R.A.; Ross, E.A.; Henderson, I.R.; Leyton, D.L.; Coughlan, R.E.; Khan, M.; Jensen, K.T.; et al. The porin OmpD from nontyphoidal Salmonella is a key target for a protective B1b cell antibody response. Proc. Natl. Acad. Sci. USA 2009, 106, 9803–9808. [Google Scholar] [CrossRef] [PubMed]
- Prejit, N.; Pratheesh, P.T.; Nimisha, S. Expression and purification of an immunogenic SUMO-OmpC fusion protein of Salmonella Typhimurium in Escherichia coli. Biologicals 2019, 62, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chakraborty, K.; Nagaraja, T.; Basak, S.; Koley, H.; Dutta, S.; Mitra, U.; Das, S. An adhesion protein of Salmonella enterica serovar Typhi is required for pathogenesis and potential target for vaccine development. Proc. Natl. Acad. Sci. USA 2011, 108, 3348–3353. [Google Scholar] [CrossRef]
- Schussek, S.; Trieu, A.; Doolan, D. Genome- and proteome-wide screening strategies for antigen discovery and immunogen design. Biotechnol. Adv. 2014, 32, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Donati, C.; Rappuoli, R. Reverse vaccinology in the 21st century: Improvements over the original design. Ann. N. Y. Acad. Sci. 2013, 1285, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Seib, K.L.; Zhao, X.; Rappuoli, R.; Seib, K.L.; Zhao, X.; Rappuoli, R. Developing vaccines in the era of genomics: A decade of reverse vaccinology. Clin. Microbiol. Infect. 2012, 18 (Suppl. S5), 109–116. [Google Scholar] [CrossRef]
- Pizza, M.; Scarlato, V.; Masignani, V.; Giuliani, M.M.; Aricò, B.; Comanducci, M.; Jennings, G.T.; Baldi, L.; Bartolini, E.; Capecchi, B.; et al. Identification of Vaccine Candidates Against Serogroup B Meningococcus by Whole-Genome Sequencing. Science 2000, 287, 1816–1820. [Google Scholar] [CrossRef]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30 (Suppl. S2), B87–B97. [Google Scholar] [CrossRef]
- Wizemann, T.M.; Adamou, J.E.; Langermann, S. Adhesins as Targets for Vaccine Development. Emerg. Infect. Dis. 1999, 5, 395–403. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, W.; Elbediwi, M.; Pan, H.; Wang, L.; Zhou, C.; Zhao, B.; Xu, X.; Li, D.; Yan, X.; et al. Characterization of Salmonella Resistome and Plasmidome in Pork Production System in Jiangsu, China. Front. Vet. Sci. 2020, 7, 617. [Google Scholar] [CrossRef]
- Vingataramin, L.; Frost, E.H. A single protocol for extraction of gDNA from bacteria and yeast. Biotechniques 2015, 58, 120–125. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An integrative web server to predict subcellular localization of proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Asakawa, N.; Tsuji, T.; Akazawa, F.; Ino, A.; Sonoyama, M.; Mitaku, S. SOSUI-GramN: High performance prediction for sub-cellular localization of proteins in gram-negative bacteria. Bioinformation 2008, 2, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-S.; Cheng, C.-W.; Su, W.-C.; Chang, K.-C.; Huang, S.-W.; Hwang, J.-K.; Lu, C.-H. CELLO2GO: A Web Server for Protein subCELlular LOcalization Prediction with Functional Gene Ontology Annotation. PLoS ONE 2014, 9, e99368. [Google Scholar] [CrossRef]
- Blobel, G.; Dobberstein, B. Transfer of Proteins across Membranes.1. Presence of Proteolytically Processed and Un-processed Nascent Immunoglobulin Light-Chains on Membrane-Bound Ribosomes of Murine Myeloma. J. Cell Biol. 1975, 67, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef]
- Sachdeva, G.; Kumar, K.; Jain, P.; Ramachandran, S. SPAAN: A software program for prediction of adhesins and adhesin-like proteins using neural networks. Bioinformatics 2005, 21, 483–491. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, B.K.; Gardner, P.P.; Lim, C.S. Solubility-Weighted Index: Fast and accurate prediction of protein solubility. Bioinformatics 2020, 36, 4691–4698. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Protein production by auto-induction in high-density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef]
- Xu, W.; Fang, S.; Wang, Y.; Zhang, T.; Hu, S. Molecular mechanisms associated with macrophage activation by Rhizoma Atractylodis Macrocephalae polysaccharides. Int. J. Biol. Macromol. 2020, 147, 616–628. [Google Scholar] [CrossRef]
- Rasmussen, D.M.; Soens, R.W.; Davie, T.J.; Vaneerd, C.K.; Bhattacharyya, B.; May, J.F. The structure of DcrB, a lipoprotein from Salmonella enterica, reveals flexibility in the N-terminal segment of the Mog1p/PsbP-like fold. J. Struct. Biol. 2018, 204, 513–518. [Google Scholar] [CrossRef]
- Cameron, E.A.; Gruber, C.C.; Ritchie, J.M.; Waldor, M.K.; Sperandio, V. The QseG Lipoprotein Impacts the Virulence of Enterohemorrhagic Escherichia coli and Citrobacter rodentium and Regulates Flagellar Phase Variation in Salmonella enterica Serovar Typhimurium. Infect. Immun. 2018, 86, e00936-17. [Google Scholar] [CrossRef]
- Reading, N.C.; Rasko, D.A.; Torres, A.G.; Sperandio, V. The two-component system QseEF and the membrane protein QseG link adrenergic and stress sensing to bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 5889–5894. [Google Scholar] [CrossRef]
- Secundino, I.; López-Macías, C.; Cervantes-Barragán, L.; Gil-Cruz, C.; Ríos-Sarabia, N.; Pastelin-Palacios, R.; Villasis-Keever, M.A.; Becker, I.; Puente, J.L.; Calva, E.; et al. Salmonella porins induce a sustained, lifelong specific bactericidal antibody memory response. Immunology 2006, 117, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.K.; Porteen, K.; Dubal, Z.B.; Asha, K.; Shweta, S.; Ripan, B. Evaluation of recombinant outer membrane protein based vaccine against Salmonella Typhimurium in birds. Biologicals 2013, 41, 162–168. [Google Scholar]
- Lambert, M.A.; Smith, S.G. The PagN protein mediates invasion via interaction with proteoglycan. FEMS Microbiol. Lett. 2009, 297, 209–216. [Google Scholar] [CrossRef]
- Juillan-Binard, C.; Picciocchi, A.; Andrieu, J.-P.; Dupuy, J.; Petit-Hartlein, I.; Caux-Thang, C.; Vivès, C.; Nivière, V.; Fieschi, F. A Two-component NADPH Oxidase (NOX)-like System in Bacteria Is Involved in the Electron Transfer Chain to the Methionine Sulfoxide Reductase MsrP. J. Biol. Chem. 2017, 292, 2485–2494. [Google Scholar] [CrossRef] [PubMed]
- Andrieu, C.; Vergnes, A.; Loiseau, L.; Aussel, L.; Ezraty, B. Characterisation of the periplasmic methionine sulfoxide reductase (MsrP) from Salmonella Typhimurium. Free Radic. Biol. Med. 2020, 160, 506–512. [Google Scholar] [CrossRef]
- Li, M.; Wang, K.; Tang, A.; Tang, A.; Chen, A.; Huang, Z. Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States. Antibiotics 2021, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Humphries, A.D.; Weening, E.H.; de Zoete, M.R.; Winter, S.; Papaconstantinopoulou, A.; Dougan, G.; Baumler, A.J. Molecular and Phenotypic Analysis of the CS54 Island of Salmonella enterica Serotype Typhimurium: Identification of Intestinal Colonization and Persistence Determinants. Infect. Immun. 2003, 71, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Zgurskaya, H.I.; Nikaido, H. AcrA is a highly asymmetric protein capable of spanning the periplasm. J. Mol. Biol. 1999, 285, 409–420. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nagasawa, S.; Hayashi-Nishino, M.; Yamaguchi, A.; Nishino, K. AcrA dependency of the AcrD efflux pump in Salmonella enterica serovar Typhimurium. J. Antibiot. 2011, 64, 433–437. [Google Scholar] [CrossRef]
- Norris, T.L.; Baumler, A.J. Phase variation of the lpf operon is a mechanism to evade cross-immunity between Salmonella serotypes. Proc. Natl. Acad. Sci. USA 1999, 96, 13393–13398. [Google Scholar] [CrossRef]
- Doughty, S.; Sloan, J.; Bennett-Wood, V.; Robertson, M.; Robins-Browne, R.M.; Hartland, E.L. Identification of a novel fimbrial gene cluster related to long polar fimbriae in locus of enterocyte effacement-negative strains of enterohemorrhagic Escherichia coli. Infect. Immun. 2002, 70, 6761–6769. [Google Scholar] [CrossRef]
- Lilic, M.; Jovanovic, M.; Jovanovic, G.; Savic, D.J. Identification of the CysB-regulated gene, hslJ, related to the Escherichia coli novobiocin resistance phenotype. FEMS Microbiol. Lett. 2003, 224, 239–246. [Google Scholar] [CrossRef]
- Keith, K.E.; Valvano, M. Characterization of SodC, a periplasmic superoxide dismutase from Burkholderia cenocepacia. Infect. Immun. 2007, 75, 2451–2460. [Google Scholar] [CrossRef]
- Tidhar, A.; Rushing, M.D.; Kim, B.; Slauch, J.M. Periplasmic superoxide dismutase SodCI of Salmonella binds peptidoglycan to remain tethered within the periplasm. Mol. Microbiol. 2015, 97, 832–843. [Google Scholar] [CrossRef]
- Lee, J.; Hiibel, S.R.; Reardon, K.F.; Wood, T.K. Identification of stress-related proteins in Escherichia coli using the pollutant cis-dichloroethylene. J. Appl. Microbiol. 2010, 108, 2088–2102. [Google Scholar]
- Tidhar, A.; Flashner, Y.; Cohen, S.; Levi, Y.; Zauberman, A.; Gur, D.; Aftalion, M.; Elhanany, E.; Zvi, A.; Shafferman, A.; et al. The NlpD Lipoprotein Is a Novel Yersinia pestis Virulence Factor Essential for the Development of Plague. PLoS ONE 2009, 4, e7023. [Google Scholar] [CrossRef]
- Ji, X.; Lu, P.; Xue, J.; Zhao, N.; Zhang, Y.; Dong, L.; Zhang, X.; Li, P.; Hu, Y.; Wang, J.; et al. The lipoprotein NlpD in Cronobacter sakazakii responds to acid stress and regulates macrophage resistance and virulence by maintaining membrane integrity. Virulence 2021, 12, 415–429. [Google Scholar] [CrossRef]
- Peng, X.; Ed-Dra, A.; Song, Y.; Elbediwi, M.; Nambiar, R.B.; Zhou, X.; Yue, M. Lacticaseibacillus rhamnosus alleviates intestinal inflammation and promotes microbiota-mediated protection against Salmonella fatal infections. Front. Immunol. 2022, 13, 973224. [Google Scholar] [CrossRef]
- Tanghe, A.; Lefèvre, P.; Denis, O.; D’Souza, S.; Braibant, M.; Lozes, E.; Singh, M.; Montgomery, D.; Content, J.; Huygen, K. Immunogenicity and protective efficacy of tuberculosis DNA vaccines encoding putative phosphate transport receptors. J. Immunol. 1999, 162, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Biswas, S.; Gu, G.; Elbediwi, M.; Li, Y.; Yue, M. Characterization of Multidrug Resistance Patterns of Emerging Salmonella enterica Serovar Rissen along the Food Chain in China. Antibiotics 2020, 9, 660. [Google Scholar] [CrossRef]
- Kariuki, S.; Gordon, M.A.; Feasey, N.; Parry, C.M. Antimicrobial resistance and management of invasive Salmonella disease. Vaccine 2015, 33, C21–C29. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R. Reverse vaccinology. Curr. Opin. Microbiol. 2000, 3, 445–450. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Rappuoli, R.; De Groot, A.S.; Chen, R.T. Emerging Vaccine Informatics. J. Biomed. Biotechnol. 2010, 2010, 218590. [Google Scholar] [CrossRef]
- Dalsass, M.; Brozzi, A.; Medini, D.; Rappuoli, R. Comparison of Open-Source Reverse Vaccinology Programs for Bacterial Vaccine Antigen Discovery. Front. Immunol. 2019, 10, 113. [Google Scholar] [CrossRef]
- Gonzalez, D.; Richez, M.; Bergonzi, C.; Chabriere, E.; Elias, M. Crystal structure of the phosphate-binding protein (PBP-1) of an ABC-type phosphate transporter from Clostridium perfringens. Sci. Rep. 2014, 4, 6636. [Google Scholar] [CrossRef]
- Xu, X.; Chen, Y.; Pan, H.; Pang, Z.; Li, F.; Peng, X.; Ed-Dra, A.; Li, Y.; Yue, M. Genomic characterization of Salmonella Uzaramo for human invasive infection. Microb. Genom. 2020, 6, mgen000401. [Google Scholar] [CrossRef]
- Lamarche, M.G.; Wanner, B.L.; Crépin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate ho-meostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 32, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.; Batista, M.; Cavalcante, R.; Pegos, V.; Passos, H.; Silva, D.; Balan, A.; Ferreira, L.; Ferreira, R. Sublingual immunization with the phosphate-binding-protein (PstS) reduces oral colonization by Streptococcus mutans. Mol. Oral Microbiol. 2015, 31, 410–422. [Google Scholar] [CrossRef]
- Humphries, A.D.; Townsend, S.M.; Kingsley, R.A.; Nicholson, T.L.; Tsolis, R.M.; Bäumler, A.J. Role of fimbriae as antigens and intestinal colonization factors of Salmonella serovars. FEMS Microbiol. Lett. 2001, 201, 121–125. [Google Scholar] [CrossRef]
- Humphries, A.; DeRidder, S.; Baumler, A. Salmonella enterica serotype typhimurium fimbrial proteins serve as antigens during infection of mice. Infect. Immun. 2005, 73, 5329–5338. [Google Scholar] [CrossRef]
- Heinz, E.; Stubenrauch, C.J.; Grinter, R.; Croft, N.P.; Purcell, A.W.; Strugnell, R.A.; Dougan, G.; Lithgow, T. Conserved Features in the Structure, Mechanism, and Biogenesis of the Inverse Autotransporter Protein Family. Genome Biol. Evol. 2016, 8, 1690–1705. [Google Scholar] [CrossRef]
- Xing, Y.; Clark, J.R.; Chang, J.D.; Chirman, D.M.; Green, S.; Zulk, J.J.; Jelinski, J.; Patras, K.A.; Maresso, A.W. Broad protective vaccination against systemic Escherichia coli with autotransporter antigens. PLoS Pathog. 2023, 19, e1011082. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Program | Web Address |
|---|---|---|
| Proteome acquisition | NCBI | https://www.ncbi.nlm.nih.gov/nuccore/nz_cp068386.1 (accessed on 10 February 2022) |
| Subcellular location | PSORTb | https://www.psort.org/psortb/ (accessed on 15 February 2022) |
| Subcellular location | CELLO2GO | https://cello.life.nctu.edu.tw/cello2go/ (accessed on 11 February 2022) |
| Subcellular location | BUSCA | https://busca.biocomp.unibo.it/ (accessed on 11 February 2022) |
| Subcellular location | SOSUI | https://harrier.nagahama-i-bio.ac.jp/sosui/ (accessed on 10 February 2022) |
| Signal peptide | SignalP-5.0 | https://services.healthtech.dtu.dk/service.php?SignalP-5.0 (accessed on 16 February 2022) |
| Antigenicity | VaxiJen-3.0 | https://www.ddg-pharmfac.net/vaxijen3/ (accessed on 18 February 2022) |
| Adhesin probability | SPAAN | https://github.com/3itamura-felipe/adhesin_finder (accessed on 22 February 2022) |
| Transmembrane domains | TMHMM-2.0 | https://services.healthtech.dtu.dk/service.php?TMHMM-2.0 (accessed on 23 February 2022) |
| Solubility | SoDoPE | https://tisigner.com/sodope (accessed on 23 February 2022) |
| Conservations in Pullorum | BLASTp | https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 28 February 2022) |
| SOSUI | BUSCA | CELLO2GO | PSORTb | |
|---|---|---|---|---|
| Extracellular | 157 | 438 | 144 | 72 |
| 3% | 9% | 3% | 2% | |
| Outer Membrane | 184 | 79 | 171 | 110 |
| 4% | 2% | 4% | 2% | |
| Periplasm | 361 | NA | 595 | 162 |
| 8% | NA | 12% | 3% | |
| Cytoplasmic | 2565 | 3052 | 2875 | 2002 |
| 54% | 65% | 61% | 42% | |
| Inner membrane | 969 | 1107 | 928 | 1158 |
| 21% | 23% | 20% | 25% | |
| Unknown | 477 | 37 | NA | 1209 |
| 10% | 1% | NA | 26% |
| Candidates | Annotation |
|---|---|
| dcrB | DcrB is up-regulated by conditions that promote the production of known virulence factors [55] |
| qseG | QseG is an OM protein necessary for translocation of TTSS effectors [56,57] |
| ompC | OmpC is the main protein responsible for the antibody-mediated memory bactericidal response induced by porins [58,59] |
| pagN | Haemagglutinin that facilitates the adhesion to and invasion of epithelial mammalian cells [33,60] |
| msrP | MSR system is involved in the repair of periplasmic proteins oxidized by hypochlorous acid HOCl, which is generated in particular within phagocytic cells [61,62] |
| sinH | sinH encodes an autotransporter protein that facilitates adhesion and invasion into host cells [63,64] |
| acrA | AcrA protein is a component of multidrug efflux pumps, which can increase drug resistance [65,66] |
| lpfA | Long polar fimbria protein A precursor [67,68] |
| peg.1230 | Putative fimbriae stiH |
| peg.1041 | Fimbrial operons sthB |
| peg.3954 | Minor fimbrial subunit StfF |
| hslJ | Heat shock protein, correlated to E. coli resistance to the antibiotic Nov [69] |
| sodC | Periplasmic superoxide dismutase plays a critical role in this survival by combating phagocytic superoxide [70,71] |
| yhcN | YhcN is involved in the response to hydrogen peroxide stress [72] |
| nlpD | A virulence factor that permits S. Typhimurium to survive under acid stress conditions [73,74] |
| pstS | Phosphate ABC transporter, periplasmic phosphate-binding protein PstS [75,76] |
| peg.695 | Involved in the transport of maltose and maltodextrins |
| chiP | Involved in the uptake of chitosugars [77] |
| peg.2134 | N-acetylneuraminic acid outer membrane channel protein NanC |
| Fdh-2 | Formate dehydrogenase |
| Peg.1210 | Probable secreted protein |
| lpfB | Chaperone protein lpfB precursor [68] |
| Candidates | Inter-Assay | Intra-Assay 1 | Intra-Assay 2 | Intra-Assay 3 | ||||
|---|---|---|---|---|---|---|---|---|
| OD492 | CV% | OD492 | CV% | OD492 | CV% | OD492 | CV% | |
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |||||
| PBS | 0.237 ± 0.015 | 6.31 | 0.231 ± 0.012 | 5.19 | 0.258 ± 0.018 | 6.98 | 0.223 ± 0.008 | 3.59 |
| StiH | 1.120 ± 0.084 | 7.46 | 1.217 ± 0.035 | 2.88 | 1.129 ± 0.032 | 2.83 | 1.013 ± 0.025 | 2.47 |
| LpfA | 0.889 ± 0.025 | 2.76 | 0.923 ± 0.019 | 2.06 | 0.880 ± 0.039 | 4.43 | 0.865 ± 0.024 | 2.77 |
| OmpC | 1.074 ± 0.063 | 5.89 | 0.986 ± 0.022 | 2.23 | 1.102 ± 0.042 | 3.81 | 1.133 ± 0.036 | 3.18 |
| PagN | 1.033 ± 0.092 | 8.92 | 0.905 ± 0.036 | 3.98 | 1.076 ± 0.033 | 3.07 | 1.118 ± 0.043 | 3.85 |
| MsrP | 0.564 ± 0.049 | 8.62 | 0.564 ± 0.028 | 4.96 | 0.504 ± 0.028 | 5.56 | 0.623 ± 0.039 | 6.26 |
| AcrA | 0.732 ± 0.104 | 14.21 | 0.876 ± 0.015 | 1.71 | 0.635 ± 0.035 | 5.51 | 0.684 ± 0.033 | 4.82 |
| StfF | 0.753 ± 0.035 | 4.60 | 0.801 ± 0.016 | 2.00 | 0.722 ± 0.017 | 2.35 | 0.735 ± 0.037 | 5.03 |
| SinH | 1.426 ± 0.076 | 5.33 | 1.376 ± 0.032 | 2.33 | 1.368 ± 0.034 | 2.49 | 1.533 ± 0.047 | 3.07 |
| SthB | 1.289 ± 0.073 | 5.66 | 1.324 ± 0.044 | 3.32 | 1.187 ± 0.022 | 1.85 | 1.355 ± 0.035 | 2.58 |
| QseG | 0.445 ± 0.044 | 9.82 | 0.483 ± 0.032 | 6.63 | 0.469 ± 0.019 | 4.05 | 0.384 ± 0.024 | 6.25 |
| HslJ | 0.485 ± 0.031 | 6.40 | 0.455 ± 0.031 | 6.81 | 0.528 ± 0.017 | 3.22 | 0.473 ± 0.019 | 4.02 |
| NlpD | 0.480 ± 0.014 | 2.87 | 0.472 ± 0.028 | 5.93 | 0.468 ± 0.024 | 5.13 | 0.499 ± 0.026 | 5.21 |
| PstS | 1.665 ± 0.120 | 7.20 | 1.597 ± 0.033 | 2.07 | 1.565 ± 0.039 | 2.49 | 1.834 ± 0.053 | 2.89 |
| Peg.695 | 0.899 ± 0.030 | 3.27 | 0.897 ± 0.042 | 4.68 | 0.936 ± 0.033 | 3.53 | 0.864 ± 0.023 | 2.66 |
| ChiP | 1.029 ± 0.083 | 8.04 | 0.968 ± 0.013 | 1.34 | 1.146 ± 0.026 | 2.27 | 0.973 ± 0.035 | 3.60 |
| Peg.2134 | 0.973 ± 0.167 | 17.19 | 0.856 ± 0.037 | 4.32 | 1.210 ± 0.021 | 1.74 | 0.854 ± 0.034 | 3.98 |
| FdhF | 0.589 ± 0.094 | 15.90 | 0.721 ± 0.043 | 5.96 | 0.514 ± 0.017 | 3.31 | 0.532 ± 0.033 | 6.20 |
| LpfB | 1.304 ± 0.071 | 5.41 | 1.372 ± 0.026 | 1.90 | 1.334 ± 0.032 | 2.40 | 1.207 ± 0.042 | 3.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Kang, X.; Song, Y.; Zhou, X.; Yue, M. Identification and Evaluation of Novel Antigen Candidates against Salmonella Pullorum Infection Using Reverse Vaccinology. Vaccines 2023, 11, 865. https://doi.org/10.3390/vaccines11040865
Jiang Z, Kang X, Song Y, Zhou X, Yue M. Identification and Evaluation of Novel Antigen Candidates against Salmonella Pullorum Infection Using Reverse Vaccinology. Vaccines. 2023; 11(4):865. https://doi.org/10.3390/vaccines11040865
Chicago/Turabian StyleJiang, Zhijie, Xiamei Kang, Yan Song, Xiao Zhou, and Min Yue. 2023. "Identification and Evaluation of Novel Antigen Candidates against Salmonella Pullorum Infection Using Reverse Vaccinology" Vaccines 11, no. 4: 865. https://doi.org/10.3390/vaccines11040865