1. Introduction
Leptospirosis, a zoonotic disease, remains a significant global public health concern, becoming even more of a threat due to the impacts of climate change and global warming, which have escalated its prevalence. Annually, there are approximately one million reported cases of human leptospirosis, resulting in an estimated 60,000 fatalities [
1]. Despite the availability of a broad spectrum of antibiotics, their effectiveness diminishes when the bacteria infiltrate vital organs and cause substantial damage, often due to delayed diagnosis. Vaccination is a cost-effective and secure preventive measure to combat this disease. However, the current killed vaccine only provides short-term immunity specific to certain serovars and fails to prevent bacterial shedding through urine. Efforts to develop live attenuated vaccines capable of inducing long-term protection have resulted in the creation and testing of various virulence-attenuated mutants [
2,
3]. Immunization with a mutant of the flagellar protein (FcpA) has demonstrated cross-protection against different pathogenic
Leptospira species, and immunization with the LPS mutant strain M1352 led to an 80% reduction in renal colonization upon exposure to the same serovar [
2]. These attenuated vaccines despite inducing robust cross-protection, have fallen short of providing sterilizing immunity, and their capacity to generate a long-lasting protective response has not been tested. Therefore, it is imperative to develop potent vaccines capable of eliciting sterilizing immunity against multiple serovars.
Subunit vaccines represent a cutting-edge generation of candidates prioritizing safety by utilizing components from the outer membranes of microbes, such as lipopolysaccharides, surface proteins, glycoproteins, and toxoids [
4,
5]. Various outer membrane and surface proteins from
Leptospira have been explored as potential subunit vaccine candidates and have demonstrated varying degrees of protection in animal models [
6]. Among these proteins,
Leptospira immunoglobulin-like protein A (LigA), specifically its C-terminal or variable region (LAV), has emerged as an exceptionally promising candidate, supported by the findings of several researchers [
7,
8]. Adjuvants are crucial in enhancing vaccine efficacy by boosting the immune response without directly contributing to antigen-specific protection [
9]. Several surface proteins from
Leptospira have been tested in combination with potent adjuvants like Freund’s adjuvant, liposomes, PLGA-microparticles, xanthan gums, as well as emulsions like AddaVax and Emulsigen-D. These combinations have provided varying degrees of protection, ranging from 50% to 70%. However, none of the formulations have provided sterilizing immunity [
7,
10,
11,
12,
13]. It is important to note that, except alum, all these adjuvants are in the preclinical stage. Fortunately, a new generation of clinical adjuvants, including Montanide, MF59 and Adjuvant Systems (AS03, AS04), have been developed, showing strong potential in enhancing antigen-specific immune responses and protective efficacy [
14]. For instance, AS04 combines monophosphoryl lipid A (MPLA), a TLR4 agonist, with an aluminum salt, and has been tested in vaccines against HPV and HBV [
15]. Our recent study has demonstrated that LAV formulated with AS04 leads to an enhanced immune response and improved protection in a hamster model of leptospirosis [
16]. These findings underscore the potential of AS04 or similar adjuvant formulations in developing a potent vaccine against leptospirosis.
Lipopolysaccharide (LPS) is a major antigen found in Gram-negative bacteria [
17]. It is a stimulant of TLR4, promoting the activation and maturation of antigen-presenting cells (APCs). However, LPS’s toxicity has hindered its use as an adjuvant in human vaccines [
18]. The removal of phosphate from lipid A, derived from the LPS of
Salmonella minnesota, has mitigated its toxicity, creating a less toxic derivative known as monophosphoryl lipid A (MPLA) [
19]. While both LPS and MPLA are recognized by TLR4, MPLA triggers a less inflammatory pathway involving a TRIF adaptor [
20]. When MPLA is formulated with alum (AS04), it can enhance the antigen-specific immune response and modulate it toward a mixed TH1/TH2 or a polarized Th1 cell response. In contrast, alum primarily elicits a skewed Th2 response [
21,
22]. Most of AS04′s adjuvant activity is attributed to the immunostimulatory properties of MPLA, although alum aids in prolonging this stimulation [
22].
Leptospira LPS, a significant antigen, induces protective immunity in animal models [
3]. It is recognized as being naturally less toxic and atypical, signaling through TLR4 and TLR2 [
23]. Considering the crucial roles of TLR2 and TLR4 in protecting against leptospirosis and recognizing the potent adjuvant effects of MPLA, it was of interest to assess the adjuvant potential of the naturally less toxic
Leptospira LPS, specifically lipid A, against leptospirosis [
16,
24].
In our current investigation, we isolated lipid A from Leptospira interrogans serovar Pomona (PLA) and conducted experiments to evaluate its immunostimulatory effects on mouse macrophages. Subsequently, we examined the adjuvant properties of PLA in mice, initially focusing on its ability to enhance the immune response to a model antigen, Ovalbumin (OVA), and then extending our assessment to a Leptospira surface antigen (LAV). We compared the immune responses elicited by these antigens when formulated with either alum alone or in combination with MPLA or PLA. To better understand how PLA functions as an adjuvant, we investigated its capacity to induce a localized inflammatory response and establish lasting immune memory. Finally, we evaluated the immune response and protective effectiveness of the PLA-based formulation in a hamster model of leptospirosis.
2. Material and Methods
2.1. Study Design
In this study, our primary objective was to investigate Leptospira lipid A’s immunostimulatory and adjuvant properties, explicitly focusing on lipid A derived from the pathogenic Leptospira interrogans serovar Pomona (referred to as PLA). We compared PLA activity alongside MPLA, a well-established stimulatory agent and clinical adjuvant. To assess PLA’s stimulatory potential, we utilized mouse macrophages to measure their capacity to induce pro-inflammatory cytokines and maturation markers. Furthermore, we examined PLA’s capability to enhance antigen-specific humoral and cellular immune responses. We initially combined PLA with a model antigen, Ovalbumin (OVA), and a 2% Alhydrogel adjuvant. This allowed us to determine the optimal dosage of PLA. Subsequently, we incorporated a variable proportion of Leptospira immunoglobulin-like protein A (LAV) into the formulation. The immune response against various formulations was analyzed in mice following established methodologies.
To gain insights into the mechanism of action and identify critical innate immune cell mediators, we administered mice with PLA- or MPLA-based formulations containing a fluorescent antigen (LAV). Using advanced techniques such as flow cytometry and RT-PCR, we evaluated immune cells’ recruitment, antigen uptake, and their activation status in draining lymph nodes. Our research also assessed the long-term memory response induced by PLA-based formulations. We monitored LAV-specific antibody levels for up to 24 weeks and subsequently analyzed B and T cell memory responses following in vitro stimulation with LAV as a recall antigen.
Finally, we critically evaluated the efficacy of the PLA-based vaccine formulation against a challenge with virulent Leptospira in a hamster model. This evaluation involved assessing the survival rate of the hamsters, quantifying the bacterial load in their organs, and examining any lesions in infected organs through a histopathological examination.
2.2. Chemicals and Reagents
Unless specifically mentioned otherwise, most of the chemical and cell culture reagents used in this study were sourced from Sigma-Aldrich. St. Louis, MO, USA. We acquired ELISA kits from R&D Biosystems, while BD Biosciences supplied the antibodies for flow cytometry. For the MPLA and 2% Alhydrogel® (alum), we procured them from InvivoGen, San Diego, CA, USA.
2.3. Animals and Housing Conditions
The animal experiments were conducted in accordance with the guidelines set forth by the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Government of India. They were approved by the Institutional Animal Ethics Committee (IAEC) under the reference numbers IAEC/2019/NIAB/18/SF and IAEC/2020/NIAB/11/SF. Female C57BL/6J mice, aged 5 to 6 weeks, were initially procured from the Jackson Laboratory in the USA. These mice were subsequently bred and housed in the Animal Resource and Experimental Facility at the National Institute of Animal Biotechnology (NIAB). Male golden Syrian hamsters were sourced from Charles River and were bred and maintained at Jeeva Life Sciences Pvt. Ltd. in Hyderabad, where the experiments were conducted.
All animals were kept in a controlled environment with a 12 h light and 12 h dark diurnal lighting cycle, maintained at a temperature of 23–24 °C and a humidity level of 45%. Standard pathogen-free conditions were ensured, and the animals had unrestricted access to food and water throughout the study. To monitor their well-being, the mice and hamsters were visually checked twice daily for vital signs and any signs of illness. After the study, the animals were humanely euthanized using 5% isoflurane following the CPCSEA’s Euthanasia Guidelines. Confirmation of death was verified through cervical dislocation, performed two minutes after breathing cessation.
2.4. Antigen Preparation
The OVA was reconstituted in endotoxin-free water (Millipore Sigma, Burlington, MA, USA) at a 1 mg/mL concentration and then stored at −20 °C. The LAV was purified using a previously established method [
16]. Briefly,
E. coli BL21 (DE3) cells carrying the expression plasmid pET28a-LAV were cultured in LB broth supplemented with kanamycin (40 μg/mL). Protein expression was induced by adding 1 mM of isopropyl β-D-1-thiogalactoside (IPTG). After induction, the cells were harvested through centrifugation at 8000 rpm for 5 min. Subsequently, cell lysis was achieved using a lysis solution (composed of 150 mM of NaCl and 100 mM of Tris-HCl at pH 8.0), followed by a sonication step. Debris was removed via centrifugation, and the resulting supernatant was subjected to affinity chromatography using a Ni-NTA bead column from GE Healthcare. The eluted protein was dialyzed against 1 × PBS and passed through Detoxi-Gel
TM from Pierce (Waltham, MA, USA) to eliminate
E. coli LPS contamination. The protein’s endotoxin level was assessed using the Limulus amoebocyte lysate (LAL) assay kit (Thermo Fischer, Waltham, MA, USA) and found to be below 0.011 EU/mL. The protein purity and size were evaluated through SDS-PAGE, and its concentration was determined using the Bradford reagent from Sigma-Aldrich.
2.5. Leptospira Culture
The pathogenic variant of L. interrogans serovar Pomona was grown in a liquid Ellinghausen–McCullough–Johnson–Harris (EMJH) medium at a temperature of 28 °C. The strain underwent a series of passages in the golden Syrian hamsters to enhance its virulence, after which it was isolated from the infected kidneys. The virulent Leptospires were subsequently kept in a semi-solid culture.
2.6. Lipid A Isolation and TLC
Lipid A was extracted from
Leptospira using a previously published method with slight modifications [
25]. In brief, logarithmic growth phase cultures of
Leptospira interrogans serovar Pomona were collected, washed twice with DPBS, and then resuspended in a Bligh–Dyer mixture consisting of chloroform, methanol, and water at a ratio of 1:2:0.8 (
v/
v). The mixture was agitated for 20 min at room temperature. Following this, the cellular fragments were separated through centrifugation at 2000 rpm for 20 min, and the lipid A component was isolated from the lipopolysaccharide (LPS) by subjecting it to an hour of boiling in a hydrolysis buffer containing 50 mM of sodium acetate and 1% sodium dodecyl sulfate, adjusted to a pH of 4.5. After cooling to room temperature, the samples underwent a second extraction step using a two-phase Bligh–Dyer mixture composed of chloroform, methanol, and water at a ratio of 2:2:1.8 (
v/
v/
v). The samples were centrifuged at 2000 rpm for 20 min to separate the phases, and the lower phase containing lipid A was transferred and subsequently dried using a rotary evaporator. The quality of the extracted lipid A was evaluated using silica thin-layer chromatography (TLC) as per the previously described method [
25]. Briefly, 10 µL of lipid A dissolved in a mixture of chloroform and methanol (4:1,
v/
v) was spotted onto a silica plate using a microcapillary glass pipette and allowed to air-dry for 15 min. The plate was then placed in a tank pre-equilibrated with the mobile phase (chloroform/pyridine/88% formic acid/water; 50:50:16:5
v/
v). Once the solvent front reached the top of the plate, it was removed, air-dried, and immersed in a 10% sulfuric acid–ethanol solution. Subsequently, the plate was air-dried in a fume hood and visualized by heating it on a 250 °C ceramic hot plate until the lipid A became charred and visible.
2.7. Cell Stimulation Assay
The isolated lipid A was carefully weighed and dissolved in endotoxin-free water (MilliporeSigma, Burlington, MA, USA) to achieve a 1 mg/mL concentration. This dissolution process was carried out by placing it in a water bath sonicator (Cole-Parmer, Vernon Hills, IL, USA) for 10 min at a temperature of 37 °C. RAW264.7 cells (obtained from ATCC, Manassas, VA, USA) were seeded into 24-well plates and then stimulated with various concentrations of PLA (0.1/0.5/1/2/5/10/20/50 µg/mL) or MPLA (0.5/1/2 µg/mL) for 24 h at 37 °C, 5% CO2. Following this stimulation, the cytokine IL-6 was measured in the culture supernatant using a Sandwich ELISA kit (R&D Systems, Minneapolis, MN, USA) following the manufacturer’s instructions.
2.8. Vaccine Formulation
The test and control formulations were prepared on the dosing day using the following method: The optimal dosage (20 or 40 µg/animal) for inducing in vivo adjuvant activity of PLA was determined with Ovalbumin. Various vaccine formulations were created by combining 100 µg of alum with MPLA (5 µg) or PLA (20 or 40 µg), along with an equal volume of DPBS containing the OVA or LAV antigen (10 µg for the first immunization, and 5 µg for the booster) in a 1:1 ratio. In contrast, formulations for hamster immunization were prepared with LAV (50 µg for the first immunization and 25 µg for the booster) in alum, along with MPLA (20 µg) or PLA (80 µg). To prepare Heat Killed Leptospira (HKL), the log-phase culture of Pomona was counted using the Petroff-Hausser Counter. The cells were washed twice with DPBS, heat-inactivated at 56 °C for 30 min, and then suspended in DPBS at a concentration of 2 × 109 cells/mL, mixed with 100 µg of Alhydrogel in a 1:1 volume ratio.
2.9. Animal Immunizations and Sample Collection
Six male C57BL/6 mice per group were subcutaneously immunized with various vaccine formulations, with a total volume of 100 μL per animal. These mice received a booster dose as described above. Blood samples were taken from the mice before immunization (pre-bleed), one week after the booster, and on the 28th day when the animals were euthanized. Spleens and lymph nodes were collected for further analysis. To assess the long-term immune response and the development of immunological memory, the mice received additional blood draws every other week for up to 24 weeks (approximately 6 months) after the booster. Some mice were euthanized with or without an additional booster, and blood, spleens, and lymph nodes were collected for memory response analysis. Tissues from the injection site were collected 4 and 24 h after immunization and preserved in an RNAprotect solution (Qiagen, Hilden, Germany).
Male golden Syrian hamsters, aged 4 to 5 weeks, were grouped and subcutaneously immunized with different formulations: PBS, heat-killed/inactivated bacterin (HKL; 109), LAV-alum LAV-alum-MPLA, or LAV-alum-PLA. The total administered volume was 200 μL, followed by a booster dose on the 21st day with 25 µg of the antigen. Before immunization, the hamsters were anesthetized using an intraperitoneal injection of ketamine (10 mg/mL) and xylazine (1 mg/mL) at 100 µL per 130 g of body weight. Blood samples were collected on days 0, 21, and 35, and the hamsters were euthanized on day 35. Spleens were collected to assess the immune response specific to the LAV antigen.
2.10. ELISA for Serum Antibody Levels
We determined the antibody levels in serum collected from the mice and hamsters using an ELISA assay, following a well-established standard protocol as previously described [
26]. In brief, 96-well microtiter plates from Nunc, Denmark, were used for the assay. These plates were coated with antigens, specifically OVA or LAV (at a concentration of 100 ng/well for mice and 200 ng/well for hamster experiments) or PLA (500 ng/well) in a 0.1 M bicarbonate buffer. The plates were then incubated overnight at 4 °C. The following day, we washed the plates three times using 1 × PBS with a 0.05% Tween 20 (PBST) solution and subsequently blocked them with 1% BSA for 1 h at room temperature. After another round of washing, we added 100 µL/well of diluted serum (ranging from 1:100 to 1:100,000 in PBST) and allowed them to incubate for 2 h at 37 °C in a humid chamber. Following this incubation, the plates were rewashed, and 100 µL/well of HRP-conjugated secondary antibodies were added (diluted at 1:6000). We used goat anti-mouse secondary antibodies specific to IgG, IgG1, IgG2c, or IgA for mice. For the hamsters, we used mouse anti-hamster antibodies against total IgG, IgG1, or IgG2/3. These were incubated for 1 h at room temperature. After another round of washing (five times), we added 100 µL/well of the TMB substrate and left it at room temperature, shielded from direct light, for 20 min. The enzymatic reaction was halted by adding 50 µL of 2N H2SO4, and we measured the optical density at 450 and 540 nm using an ELISA reader from Perkin Elmer.
2.11. Cell Proliferation and Cytokine Estimation
Lymphocyte proliferation was assessed using splenocytes from different groups of immunized mice and hamsters. To determine proliferation, the cells were stimulated with recall antigens (OVA or LAV) and then counted after 48–72 h, as previously described [
16]. In brief, splenocytes (1 × 10
5 cells/well) were plated in a 24-well plate and exposed to varying concentrations (1, 2.5, and 5 µg/mL) of OVA or LAV for 48 to 72 h. Cell counting was performed using the trypan blue size exclusion method. The levels of IL-4 and IFN-γ released into the culture supernatant after 48 h were quantified using sandwich ELISA kits (R&D Systems), following the manufacturer’s instructions. Lymphocytes from various groups of hamsters were stimulated with varying concentrations (1, 2, and 10 µg/mL) of LAV for 48 to 72 h. Subsequently, the cells were collected, RNA was isolated, and the expression of IL-4 and IFN-γ was analyzed through RT-PCR.
2.12. Flow Cytometry
We initiated the experiment by seeding RAW264.7 cells in 12-well plates at a density of 0.1 × 106 cells per well to investigate the impact of PLA treatment on macrophage activation. These cells were exposed to either MPLA (1 µg/mL) or lipid A (2 or 5 µg/mL) for 24 h. After treatment, the cells were washed twice with prechilled PBS followed by blocking with rat anti-mouse CD16/CD32 (FC antibody; 553141, BD Biosciences, Franklin Lakes, NJ, USA) in a solution of PBS containing 0.5% (w/v) BSA and 2% (v/v) FBS for 30 min. Following this, the cells underwent three additional washes and were subsequently incubated on ice with the following antibodies obtained from BD Biosciences: PerCp Cy5.5 rat anti-mouse I-A/I-E (MHC-II; 562363); APC hamster anti-mouse CD80, 560016; PE rat anti-mouse CD86, 560601, and BV421 rat anti-mouse CD40, 562846. Unbound antibodies were removed through three washes, and the cells were fixed using 1% paraformaldehyde.
To investigate the adjuvant mediated recruitment, antigen uptake, and activation status of APCs, we utilized LAV labeled with Alexa Fluor™ 488 (Invitrogen-A10235, Waltham, MA, USA) formulated with various adjuvants. These formulations were injected, and draining lymph nodes (DLNs) were isolated at 4 and 24 h post-injection. The DLN cells were washed and subjected to Fc receptor blocking at 4 °C for 30 min. Subsequently, these cells were stained with BV-421 hamster anti-mouse CD11c (562782) and PerCp Cy5.5 rat anti-mouse I-A/I-E (MHC-II; 562363) for dendritic cells; BV510 rat anti-mouse CD11b (562950) and PE-CF 594 Rat anti-Mouse F4/80 (565613) for monocytes/macrophages; BV510 rat anti-mouse CD11b (562950) and BV605 rat anti-mouse Ly6C (563011) for monocytes; BV510 rat anti-mouse CD11b (562950) and PE-Cy7 rat anti-mouse Ly6G (560601) for neutrophils; and APC hamster anti-mouse CD80 (560016), PE rat anti-mouse CD86 (560601), and PerCp Cy5.5 rat anti-mouse I-A/I-E (MHC-II; 562363) to identify specific cell types and assess their activation status.
In a separate series of experiments to determine the T cell population, splenocytes from different groups obtained on the 28th day or at 24th and 25th week post-immunization were stained with antibodies from BD Biosciences, including rat anti-mouse FITC-CD3 (555274), BV421-CD4 (562891), PE-CD8a (567630), APC-Cy 7-CD44 (560568) and BV650-CD62L (564108). Similarly, splenocytes obtained from various groups of hamsters (on the 35th day) were stained with eBiosciences APC rat anti-mouse CD4 (17-0041-82)/PE mouse anti-rat CD8b (12-0080-82). Following staining, the cells were washed and fixed with 1% paraformaldehyde. Data acquisition (100,000 events/sample) was performed using the BD LSRFortessa™ Cell Analyzer, and subsequent analysis was carried out using the FlowJo software (BD Biosciences, Franklin Lakes, NJ, USA).
2.13. Generation of Bone Marrow-Derived DCs
The dendritic cells derived from bone marrow were prepared using a previously established protocol [
16]. In brief, bone marrow cells were collected from mice and cultured in 100 mm dishes at a density of 1 × 10
7 cells per dish. These cells were cultured in a complete DMEM medium supplemented with GM-CSF (20 ng/mL; Preprotech, Thermo Fischer Scientific, Cranbury, NJ, USA) and IL-4 (5 ng/mL; Preprotech). To replenish the culture, half of the medium in each dish was replaced with fresh, complete DMEM media containing GM-CSF (40 ng/mL) and IL-4 (10 ng/mL) on the second and seventh days. On the ninth day, non-adherent cells were removed, while the adherent cells were gently scraped off. The cell count and viability were assessed, and these cells were subsequently utilized for specific assays.
2.14. CTL Assay
Antigen-specific cytotoxic T cells (CTLs) generated on the 28th day post-immunization were assessed through an LDH-based cytotoxicity assay, as previously detailed [
16]. To outline the procedure briefly, we prepared a suspension of 3 × 10
7 splenocytes in 10 mL of complete RPMI medium supplemented with 0.2 ng/mL of IL-2, which was then cultured in 25 cm
2 tissue culture flasks. After 5 days of upright maintenance, the non-adherent cells were collected and utilized as effector (E) cells.
In parallel, dendritic cells (DCs; 5 × 10
5) were seeded into 12-well plates and exposed to 2 µg/mL of OVA, LAV, or BSA (Bovine Serum Albumin) for 16 h at 37 °C with 5% CO
2. These treated DCs were employed as target cells (T) and were further treated with mitomycin C (50 μg/mL) for 45 min. The E-cells were counted and then incubated with the target cells (T), creating different E/T ratios (10:1, 25:1, and 50:1), followed by an additional 5 h incubation at 37 °C with 5% CO
2. In this setup, DCs pulsed with OVA or LAV acted as specific target cells, while DCs pulsed with BSA represented non-specific target cells. The cytotoxicity evaluation was conducted using the LDH assay kit (Cytotox 96, Promega), focusing on assessing the specific lysis of target cells by CTLs, as previously outlined [
27]. The maximum release was achieved by lysing DCs with Triton X-100 at a final concentration of 1% (vol/vol), while the LDH released by untreated cells was considered the spontaneous release. Cytotoxicity was calculated using the following formula:
2.15. Lymph Node Sectioning and Immunofluorescent Staining
The inguinal draining lymph nodes from different groups were harvested on the 28th day and processed following previously established protocols. In brief, the lymph nodes were initially fixed in a 4% paraformaldehyde (PFA) solution for one hour, then gradually dehydrated using sucrose solutions of 10%, 20%, and 30% at 4 °C. Afterward, the lymph nodes were cryopreserved in liquid nitrogen within cryomolds and stored at −80 °C until further processing.
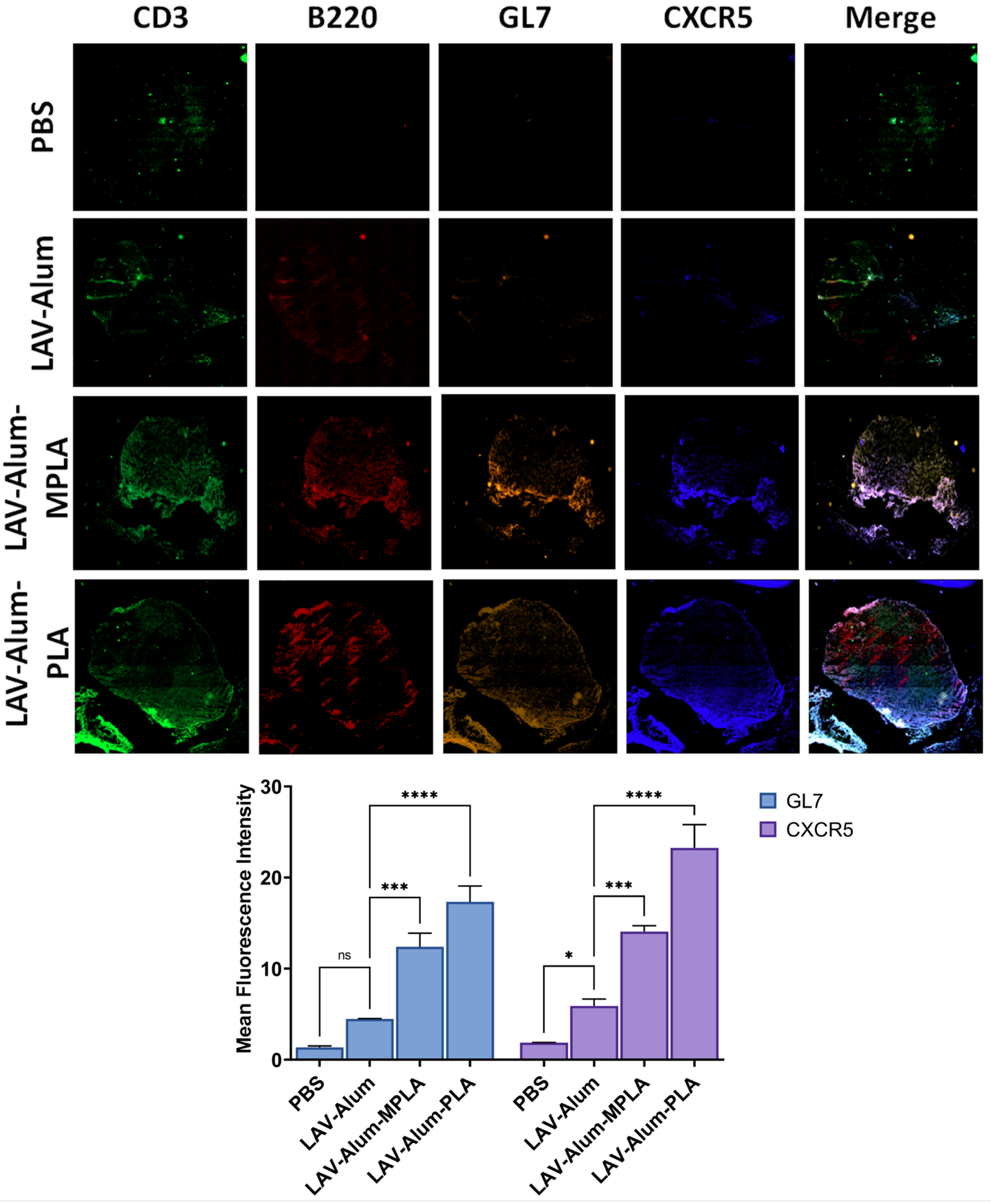
A Leica Cryostat (Leica Geosystems, Aarau, Switzerland) was employed to create 12 μm thick slices for sectioning. These sections were rehydrated in phosphate-buffered saline (PBS; pH 7.4) for three hours and then blocked using a solution of 5% BSA and 0.05% Tween 20 in PBS for one hour. Immunostaining was performed using the following antibodies: Rat anti-mouse FITC-conjugated CD3 (555274, BD Biosciences, 1:300), rat anti-mouse Per-CP-conjugated CD45R (B220; 130-102-815, Miltenyi Biotec, San Jose, CA, USA, 1:100), rat anti-mouse PE-conjugated GC B-Cell marker (GL-7; 561530, BD Biosciences, 1:200), and rabbit anti-mouse Follicular Helper T cell marker CXCR5 (ab254415, Abcam, 1:500). This incubation was carried out overnight in a moist chamber at 4 °C. Following immunostaining, the sections were thoroughly washed five times with PBS and then incubated with the secondary antibody, goat anti-rabbit Alexa Fluor® 405-conjugated IgG H&L (ab175652, Abcam, 1:300), for two hours at room temperature. The antibody-stained sectioned slides were mounted using the VECTASHIELD® Antifade mounting medium from Vector Laboratories. Imaging was performed using a Carl Zeiss Axio Scope VII fluorescent microscope with a 20× Plan Apochromat 0.45 NA objective and an EXFO X-Cite metal halide light source. The images were captured in tile-scanning mode using a Hamamatsu ORCA-ER CCD camera, and subsequently, the Zen Blue software (Version 3.7, Carl Zeiss, Jena, Germany) was used for image processing and stitching.
2.16. qRT-PCR
The previously outlined standard protocol was employed to evaluate the expression of various cytokine and chemokine genes [
16]. In brief, 800 µL of TRIzol (Invitrogen, Carlsbad, CA, USA) and 200 µL of chloroform were added to mouse tissues or a hamster cell pellet. These samples were subsequently lysed and centrifuged at 12,000 rpm for 5 min at 4 °C. RNA was then purified from the resulting aqueous phase using RNA easy mini columns (MNs), following the manufacturer’s instructions. To assess the quality of the RNA, it was subjected to electrophoresis on a formaldehyde gel to identify the 16s and 18s RNA bands, and it was also analyzed using a bioanalyzer. The quantity and purity of the RNA were measured with UV spectroscopy from Invitrogen at a 260/280 ratio. The PrimeScript 1st strand cDNA Synthesis Kit (Takara Bio, San Jose, CA, USA) was utilized as per the manufacturer’s instructions to synthesize the first-strand cDNA. The qRT-PCR procedure was performed using 96-well microtiter plates on a Bio-Rad system. The amplification process involved two steps with a reaction volume of 10 µL. The reaction mixture comprised 50 ng of cDNA, 10 pM of each primer listed in
Supplementary Table S3, and SYBR green dye from Bio-Rad. Three replicates of each sample were analyzed, and the data were evaluated using the fold change (2
−ΔΔCt) method. Changes in gene expression due to the treatment at different time points were compared to the control condition. The mRNA quantities of the analyzed genes were normalized using the corresponding reference housekeeping genes (β-actin or GAPDH). All primers used in the study were synthesized via IDT, and their sequences can be found in
Supplementary Table S3.
2.17. Infection Experiments
The hamsters were subjected to an intraperitoneal challenge on day 35, two weeks after receiving a booster injection. This challenge involved exposing them to a 100-fold effective dose (ED50) of virulent
L. interrogans serovar Pomona. The ED50 was determined using the methodology described in a previous study [
27]. We monitored the hamsters for specific clinical signs throughout four weeks. These signs included hematuria, a loss of appetite, difficulties walking or breathing, disheveled fur, a hunched posture, prostration, or weight loss exceeding 20%. We observed these physiological indicators three times daily. Hamsters with severe clinical symptoms (reaching a moribund state) were euthanized after blood collection and recorded as deceased. For those hamsters that withstood the challenge, blood samples were taken, and they were sacrificed at the end of the monitoring period. We collected the kidneys, liver, and lungs under aseptic conditions to evaluate the bacterial load and histopathological changes.
2.18. Determination of Bacterial Burden
We assessed the bacterial quantity within the organs of all infected animals, including those that met the specified endpoint criteria and those that survived until the end of the experiment before being euthanized. To determine the bacterial load, we utilized the quantitative RT-PCR method, which was previously described [
28]. The kidney, liver, and lung tissues were dissected into smaller fragments and preserved in RNAprotect (Qiagen) for subsequent procedures. We followed a standard protocol to extract the total DNA content from these tissues. Quantification was performed using the Bio-Rad Real-Time PCR System with the 2 × SYBR Green PCR Master Mix (Bio-Rad, Berkeley, CA, USA) and specific primers designed to amplify
Leptospiral 16s rRNA and LipL32 genes. We employed 10-fold serial dilutions of bacterial DNA to establish a
Leptospiral DNA standard curve, resulting in a range from 2 × 10
1 to 2 × 10
9 cells/mL.
2.19. Histopathology
The structural integrity of hamster tissues was preserved through collection, fixation, and immersion in a 10% neutral buffered formalin solution. Subsequently, these tissues were sliced into 5 μm sections using a microtome, stained with hematoxylin and eosin, and examined under light microscopy. An experienced veterinary pathologist, unaware of the sample identities, assessed the lesions induced by
Leptospira in the infected organs. The severity of tubulointerstitial nephritis was determined using a grading system that ranged from 0 (indicating normal) to 3 (indicating severe) based on established criteria [
28]. Likewise, the lung and liver pathology was assessed by counting the average number of inflammatory foci in 10X field of view. The grading scale for lung and liver pathology ranged from 0 (indicating normal) to 3 (indicating severe with more than 7 inflammatory foci).
2.20. Statistical Analysis
In most conducted experiments, the results were assessed using a one-way analysis of variance (ANOVA) along with the Dunnett hypothesis test unless stated otherwise. The data are presented as the average of three replicates, accompanied by the standard error of the mean (SEM). Statistical significance was determined using a significance threshold of p < 0.05.
4. Discussion
Currently, the primary approach for treating and preventing leptospirosis during an outbreak relies on broad-spectrum antibiotics for therapeutic and prophylactic purposes [
30]. However, indiscriminate antibiotic use to protect susceptible humans and animals from infection and subsequent transmission after a leptospirosis outbreak is impractical and can potentially lead to antibiotic resistance [
31]. Although vaccination is the most effective and cost-efficient intervention, the existing killed vaccine offers only short-term and serovar-specific protection, falling short of complete immunity [
3]. This vaccine is primarily administered to animals, and due to its toxicity, it is avoided in humans [
2]. To address the limitations of conventional whole-cell vaccines, efforts have been made to identify protective surface antigens for developing modern subunit vaccines [
6,
32]. Nonetheless, these vaccines require potent adjuvants to enhance their efficacy [
33]. Bacterial LPS, including lipid A, has been extensively studied as an immunostimulatory agent, with numerous investigations exploring the structure–activity relationship of these Toll-like receptor 4 (TLR4) ligands and their potential as adjuvants [
34]. Since the clinical approval of monophosphoryl lipid A (MPLA) as an adjuvant, particularly in the AS0X series by GSK, its use has expanded as a crucial component in vaccines against various infectious diseases [
35,
36,
37]. The development of synthetic Glucopyranosyl lipid A (GLA) further underscores the significance of bacterial lipid A as an adjuvant [
38]. Our recent study demonstrated that MPLA can enhance the antigen-specific immune response and provide protection against
Leptospira infection in a hamster model [
16]. Given that
Leptospira LPS is a major surface antigen, the protective immunity generated by whole-cell vaccines is believed to be primarily mediated by anti-LPS IgMs [
39]. Indeed, several studies have shown the role of LPS in conferring protection against challenges in animal models [
40]. This is further supported by the fact that TLR4 (which LPS binds as a ligand) plays a pivotal role in protecting against the disease [
23].
Leptospira LPS is naturally less endotoxic and atypical as it activates both TLR2 and TLR4 to induce an inflammatory response [
24,
41]. Moreover, there is no need for structural modifications to LPS or lipid A to minimize toxicity while preserving the adjuvant effect. Considering these findings, we were motivated to assess the adjuvant potential of
Leptospira lipid A.
We isolated LPS and lipid A from three significant pathogenic serovars (Icterohaemorrhagiae, Hardjo, Pomona) of
Leptospira interrogans that infect various hosts and examined their innate activity on mouse macrophages. Regardless of the serovar from which it is purified, all lipid A molecules induced a pro-inflammatory response through the TLR4 MyD88-dependent pathway, leading to macrophage activation. However, it is worth noting that the effective dose required for this response was considerably higher than that of MPLA (unpublished data). Interestingly, LPS isolated from serovar Pomona appeared to activate macrophages through a less inflammatory TRIF pathway, resembling the activity of MPLA [
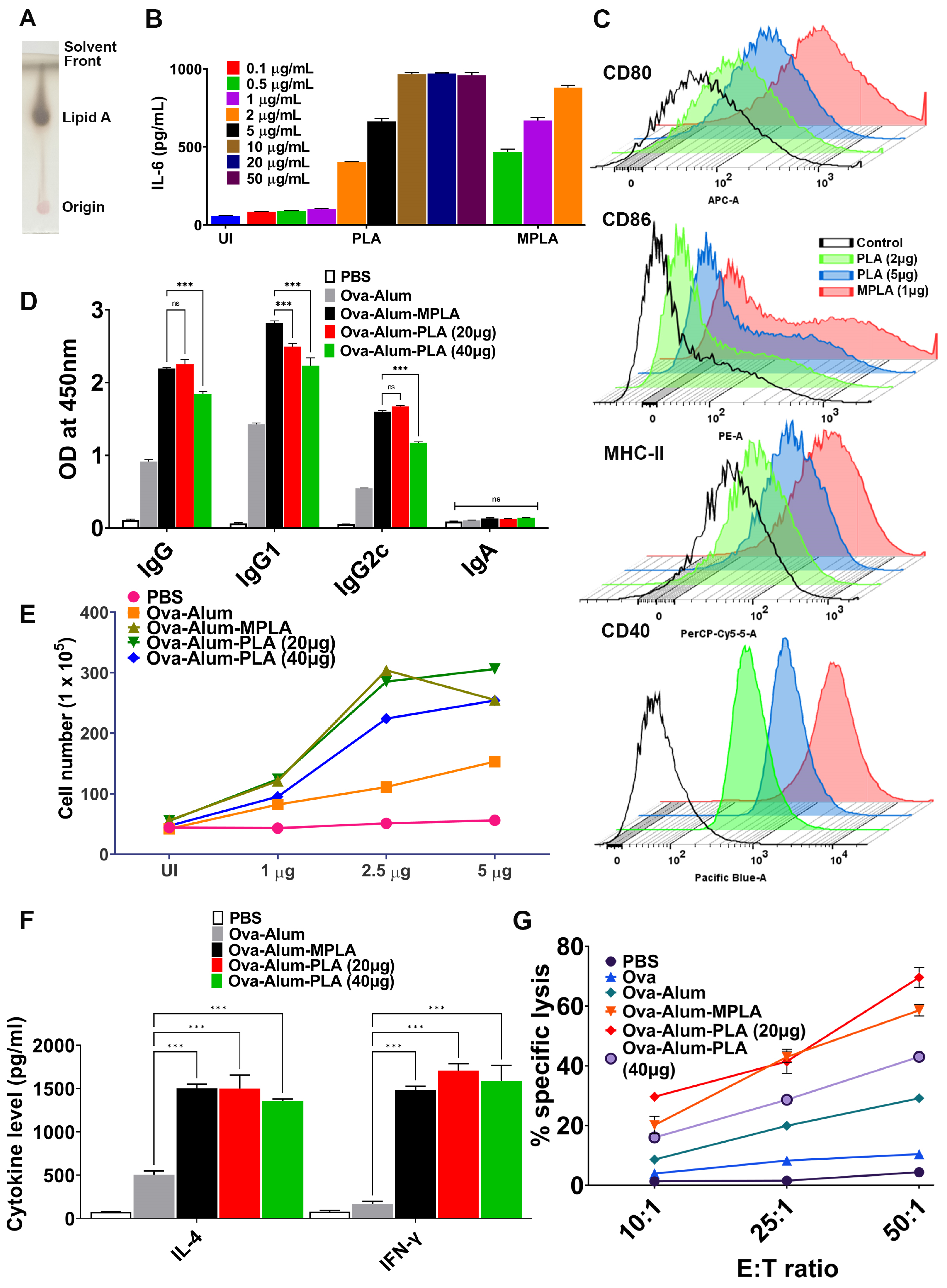
20]. These findings prompted us to explore the potency of PLA in enhancing antigen-specific immune responses. Our in vitro results suggest that PLA may be less stimulatory than MPLA, as it requires a much higher dose to induce pro-inflammatory cytokines and the expression of activation and maturation markers, as shown in
Figure 1B. This difference in activity could be attributed to the source of these molecules, with MPLA being a highly pure commercial product, while PLA is lab-purified. Additionally, the variation in immunostimulatory activity between MPLA and PLA might also arise from structural distinctions in their respective lipid A molecules. Although the lipid A structure appears to be conserved within the three pathogenic serovars we investigated (as indicated by our unpublished data), recent reports suggest structural diversity in lipid A across the broader genus of
Leptospira [
42].
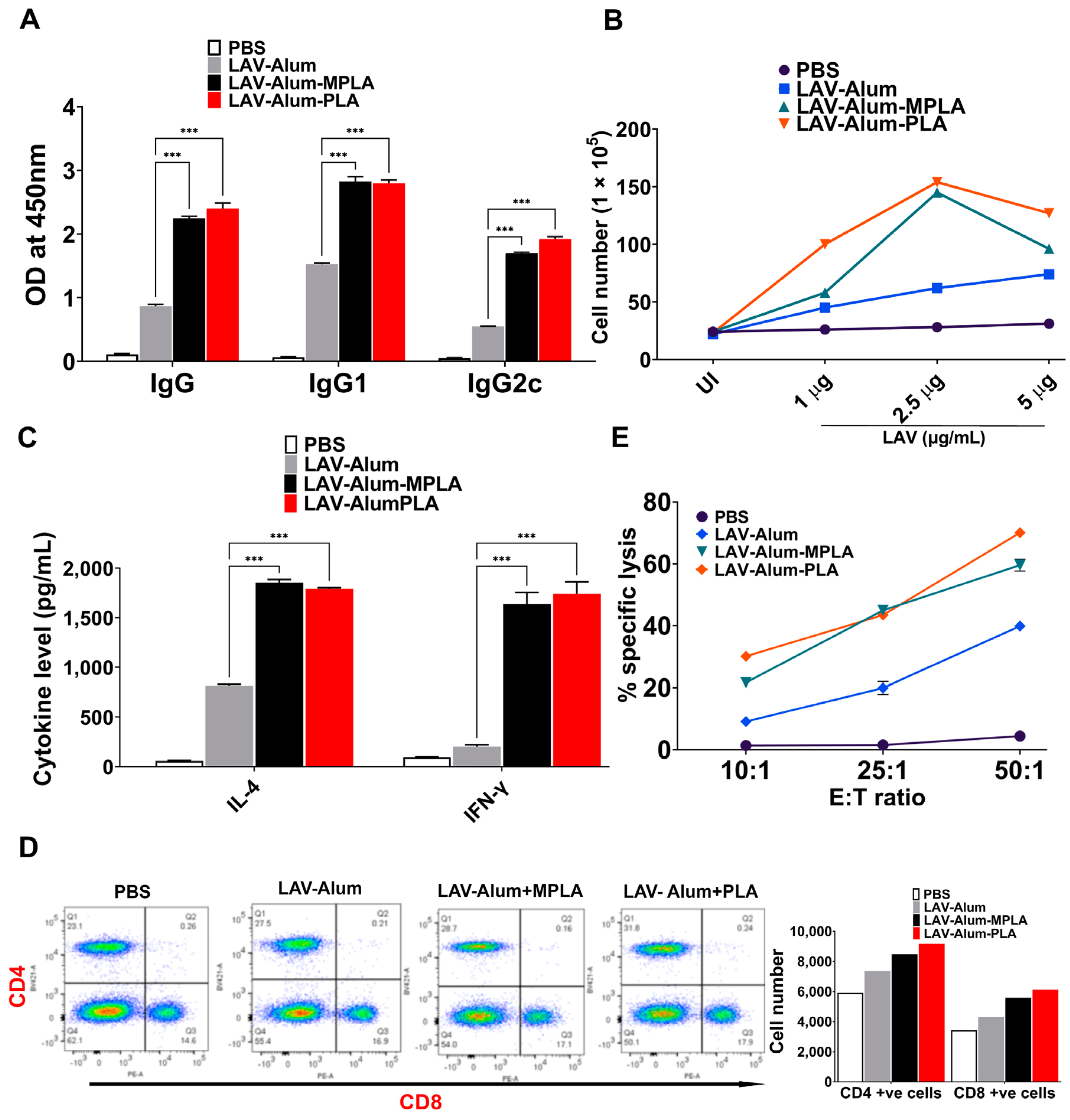
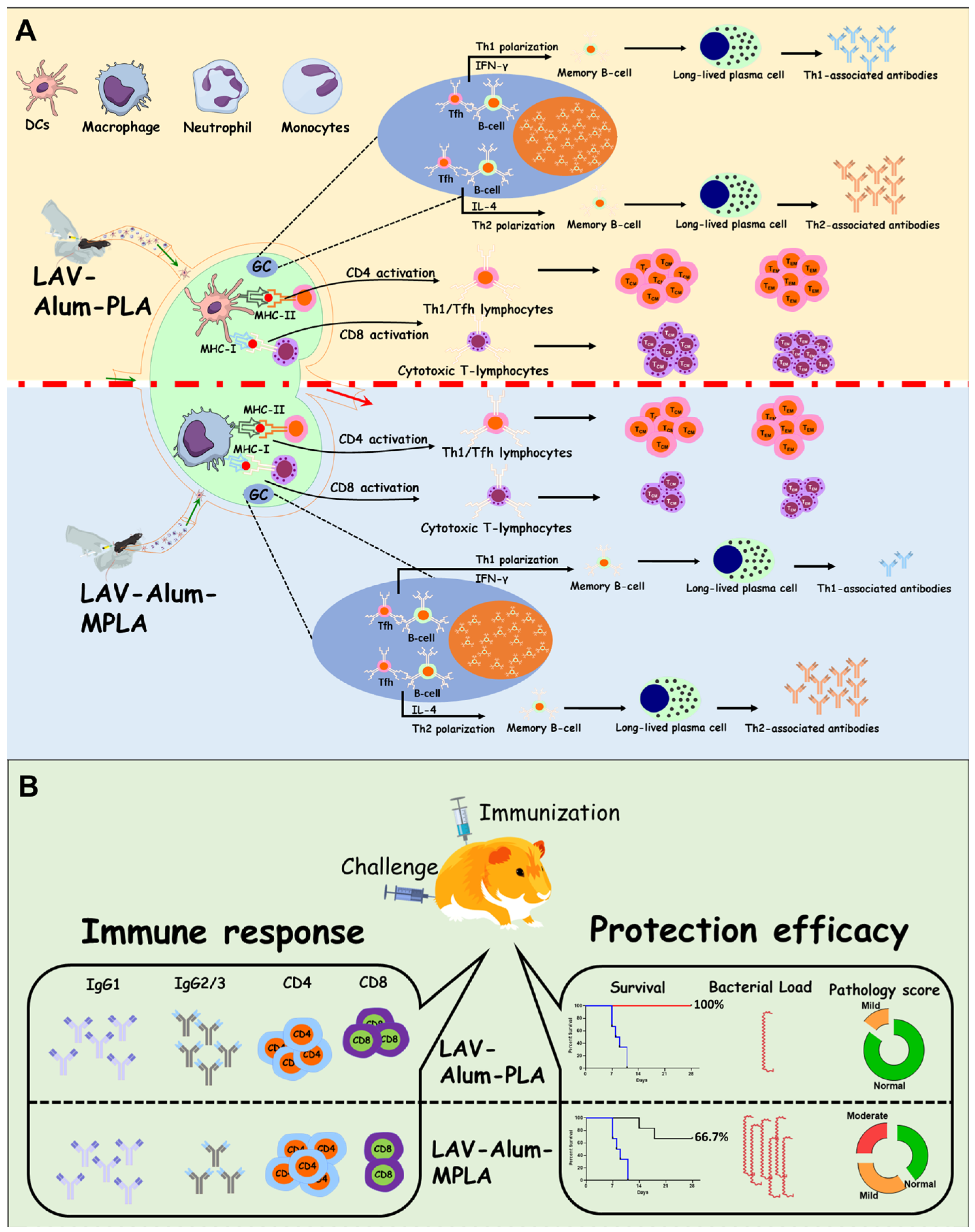
Our in vivo data indicate that alum-PLA elicits significantly higher levels of OVA and LAV-specific humoral and cellular immune responses compared to those induced by alum alone, albeit at a much higher dose (20 µg) than MPLA (5 µg), confirming the generally lower stimulatory activity of PLA compared to MPLA (as shown in
Figure 1 and
Figure 2). The mechanism through which PLA enhances antigen-specific humoral and cellular immune responses appears to be similar to MPLA and is primarily mediated by an enhanced recruitment, activation, maturation, and uptake of antigens by antigen-presenting cells (APCs), ultimately leading to the activation of B and T cell responses [
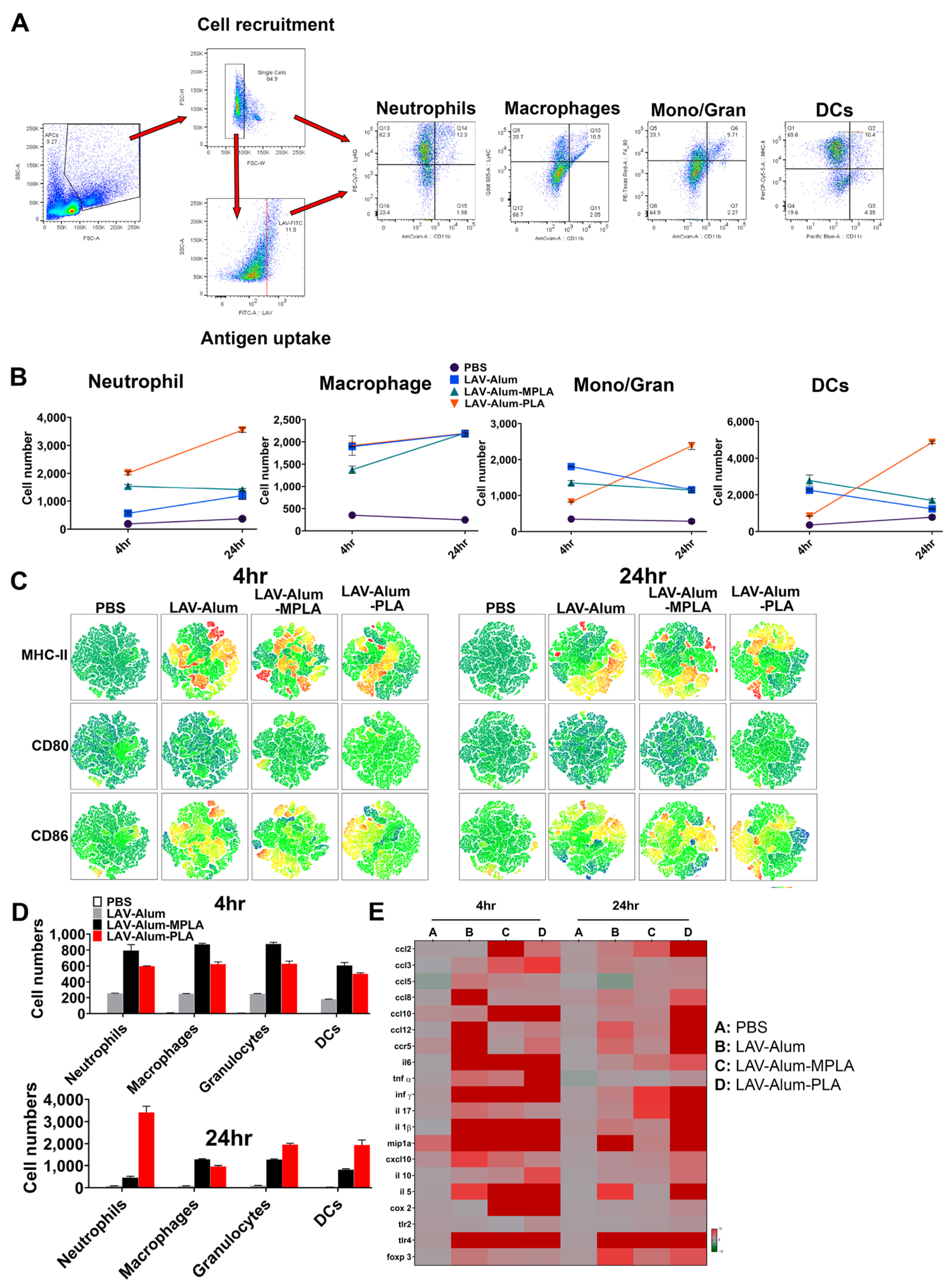
22]. Our results support this, demonstrating the rapid appearance of antigen-loaded and activated APCs in the draining lymph nodes for both PLA- and MPLA-formulated vaccines (as shown in
Figure 3B,D). The cytokines induced at the injection site by the PLA-formulated vaccines correlate with the cellular events detected in the draining lymph nodes (
Figure 3E) [
43]. Chemokines such as CCL2 and CCL3, enhanced by both the PLA and MPLA formulations, are known to promote the recruitment of monocytes and immature DCs [
44]. Differentiated APCs play a critical role in inducing adaptive responses, as they can take up antigens and migrate to the draining lymph nodes [
44]. Our results demonstrate that the formulation of PLA with alum has resulted in optimal APC recruitment and activation in the draining lymph nodes. This may be partly attributed to alum’s ability to prolong the cytokine response to PLA. The direct activation of APCs, such as DCs, by PLA or MPLA may also be crucial for sustaining an antibody response. In fact, it is believed that the direct activation of DCs by TLR agonists, rather than cytokines alone, can enhance their ability to promote antigen-specific immune responses [
45]. The immunostimulatory activity of PLA or MPLA may indirectly play a significant role in enhancing the antibody response, as pro-inflammatory cytokines are known to stimulate T helper cells specialized in assisting B-cells [
46]. Although we have not tested the direct effect of PLA on T cells, it is possible that PLA could activate these cells through TLR4 and enhance the expression of activation markers (CD69) and costimulatory ligands (CD40L) [
47].
The strong and similar levels of innate and antigen-specific adaptive immune responses generated by both MPLA and PLA can be attributed, in part, to the structural similarities and differences between these two immunostimulatory molecules. Both lipid A molecules contain amide linkages, with PLA having a higher number of them. These amide linkages may lead to a more stable and robust interaction with TLR4, resulting in a strong innate immune response followed by a subsequent adaptive immune response [
48,
49]. This phenomenon has been previously observed during
N. meningitidis infection, where the robust immune response is attributed to the strong interaction of its lipid A (containing amide-linked fatty acids) with TLR4 [
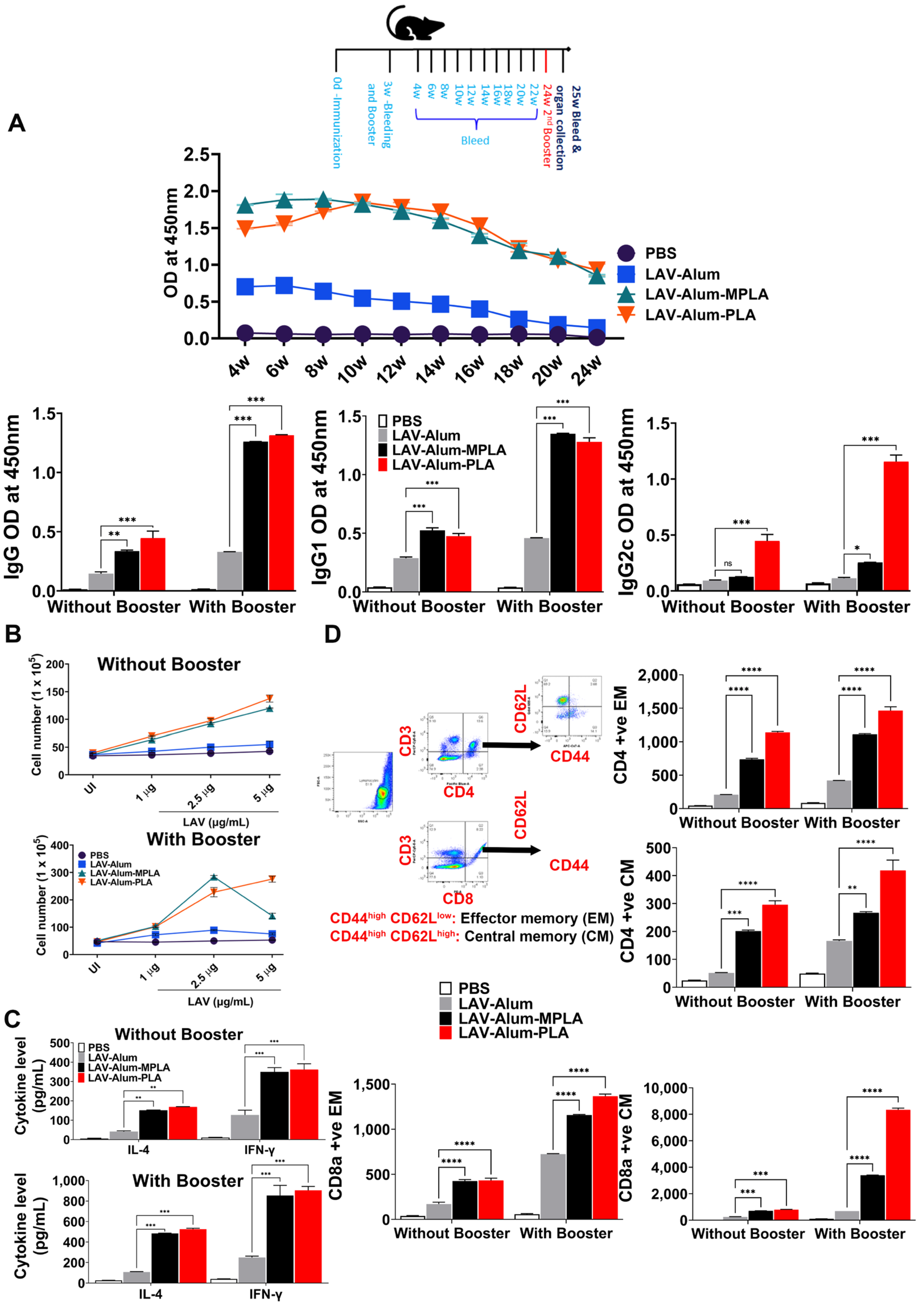
50]. Our results have demonstrated that both MPLA- and PLA-based vaccine formulations induced long-term and persistent immune responses, ultimately leading to the establishment of immunological memory. This persistence can also be attributed to the presence of amide linkages in their lipid A structures (as shown in
Figure 4). Amide linkages contribute to stability and resistance to hydrolysis by acylase enzymes, allowing lipid A to persist for a longer duration and sustain the immune response. Another structural difference between PLA and MPLA is that PLA contains two unsaturated fatty acyl chains. Unsaturated fatty acids within the lipid A structure have been shown to enhance TLR recognition and influence the host’s immune response [
51]. These unsaturated fatty acids can induce structural modifications in lipid A, such as the formation of kinks, which facilitate a better fit into the hydrophobic pocket of MD-2, an accessory protein required for TLR4 signaling. This enhanced binding affinity leads to the increased activation of TLR4-mediated immune responses. In
Salmonella enterica serovar Minnesota, the modification of lipid A with unsaturated fatty acids has been associated with increased cytokine production and inflammation in host cells [
52]. Molecular docking studies have further confirmed that the kinks in lipid A from
E. coli, caused by unsaturated fatty acids, result in more extensive interactions with TLR4 and MD-2 compared to lipid A with saturated fatty acids [
53]. Despite containing more amide linkages and unsaturation, PLA exhibits lower stimulatory activity than MPLA. This difference in activity might be attributed to the presence of methylated phosphate groups in PLA, which could induce distinct recognition patterns with TLR4 and TLR2 [
54]. However, the amide linkages in PLA contribute to its stability, allowing it to induce a sustained but lower level of stimulatory activity.
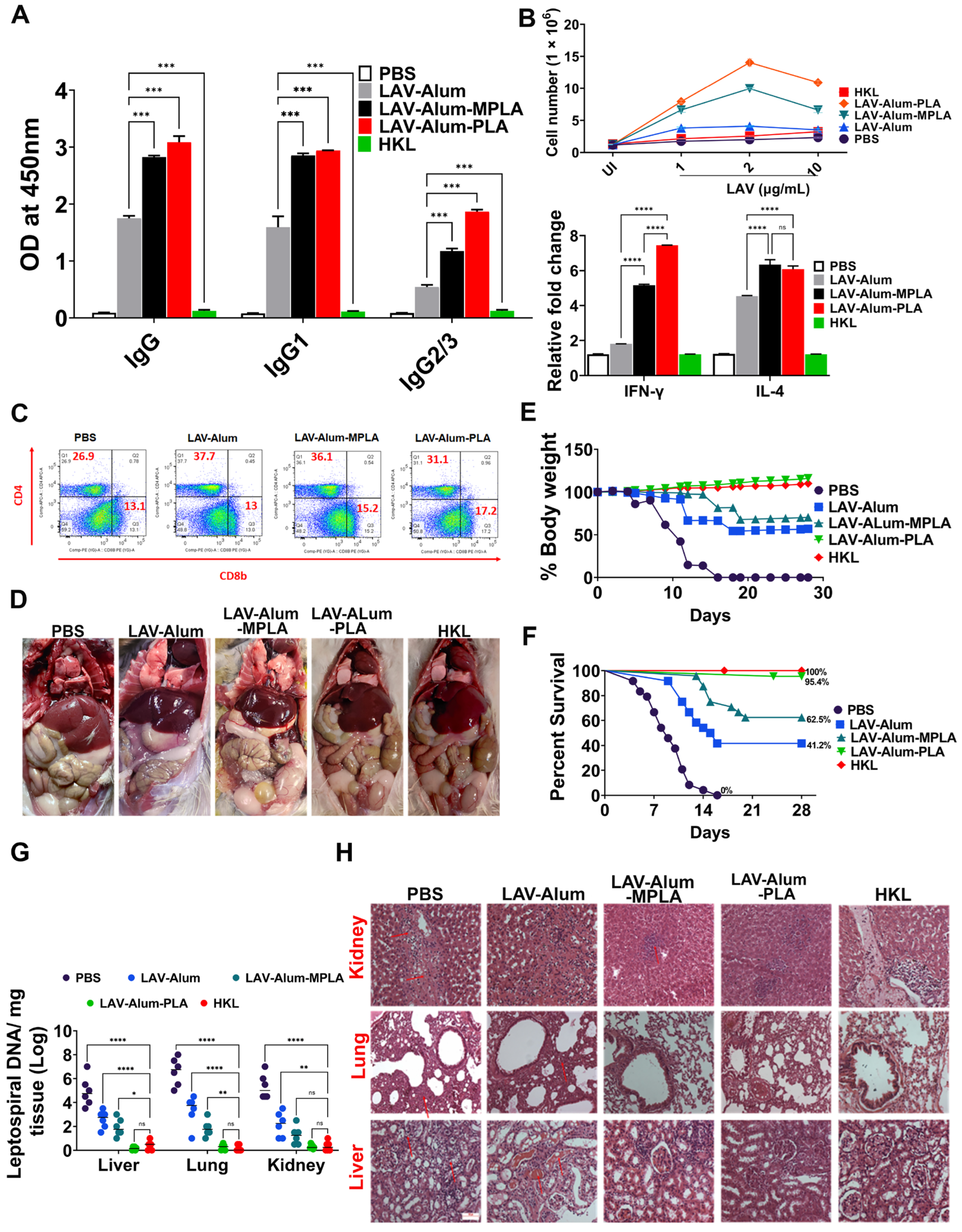
To evaluate the effectiveness of the LAV-alum-PLA formulation in conferring protection against disease, we conducted experiments using a hamster model of leptospirosis. Our findings demonstrate that hamsters immunized with LAV-alum-PLA exhibited significantly elevated levels of antigen-specific antibodies and increased concentrations of both IL-4 and IFN-gamma compared to those immunized with LAV-alum (refer to
Figure 6A,B). This robust immune response was associated with protection, as indicated by many surviving animals with reduced bacterial burden and fewer lesions in their vital organs (see
Figure 6D–F). Interestingly, while LAV-alum-PLA and LAV-alum-MPLA induced similar immune responses, the former provided superior protection, akin to the levels observed with HKL, in terms of the number of survivors and the reduction in bacterial load in organs (see
Figure 6G). This observed difference in the level of immune response and protection between LAV-alum-MPLA and LAV-alum is consistent with prior research, including our studies, where MPLA-based formulations conferred significantly higher levels of protection (ranging from 60 to 67%) compared to alum [
12,
13,
16]. It is worth noting that PLA induced better protection than the MPLA-based formulation and, in some cases, even achieved sterilizing immunity despite generating similar levels of immune response specific to LAV. We hypothesized that, aside from anti-LAV antibodies, anti-PLA antibodies may enhance protection [
55]. Indeed, we detected significant levels of anti-PLA antibodies in animals immunized with LAV-alum-PLA (see
Supplementary Figure S1). The improved efficacy of the formulation may also be attributed to the activation of additional innate pathways, particularly those involving signaling through TLR2; however, further testing and validation of this hypothesis are required [
56]. It is essential to highlight that the protection provided by HKL is achieved by generating an immune response against a range of surface antigens, including LPS. Notably, LAV-specific antibodies and T cells did not contribute to protection, as the bacteria cultured in vitro do not express LigA [
57]. Considering the variation in O-antigens among different serovars and the conservation of lipid A among pathogenic species, it is plausible that the PLA-formulated vaccine may induce cross-protection against challenge with heterologous serovars. However, this remains an active area of investigation, and ongoing experiments are addressing this aspect.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






