
A Next-Generation Adenoviral Vaccine Elicits Mucosal and Systemic Immunogenicity and Reduces Viral Shedding after SARS-CoV-2 Challenge in Nonhuman Primates

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
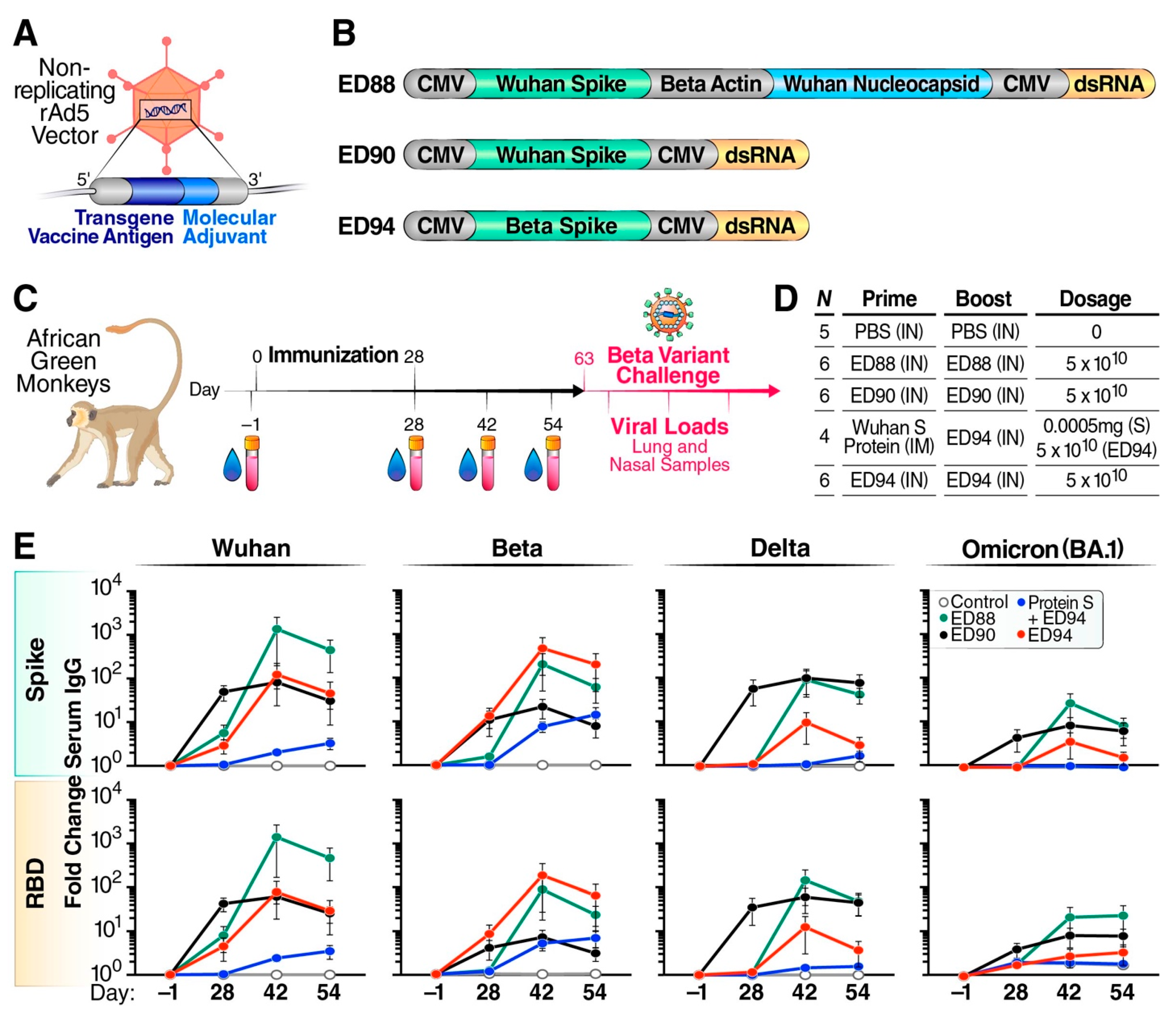
2.1. Adenoviral Vaccine Constructs
2.2. Ethics Statement
2.3. Study Design, Immunization, and Challenge
2.4. Serum and Mucosal Antibody Responses to SARS-CoV-2 Variants
2.5. Nasal and BALF Angiotensin-Converting Enzyme-2 (ACE-2) Functional Antibody Response by Surrogate Virus Neutralization Test (sVNT)
2.6. Serum Neutralizing Antibodies by Plaque Reduction Neutralization Test (PRNT)
2.7. Spike-Specific T-Cell Secretion of IFN-γ by ELISpot
2.8. Viral Load and Shedding
2.9. Statistical Analysis
3. Results
3.1. Mucosal Vaccination Elicits Strong Cross-Reactive Systemic Immunity
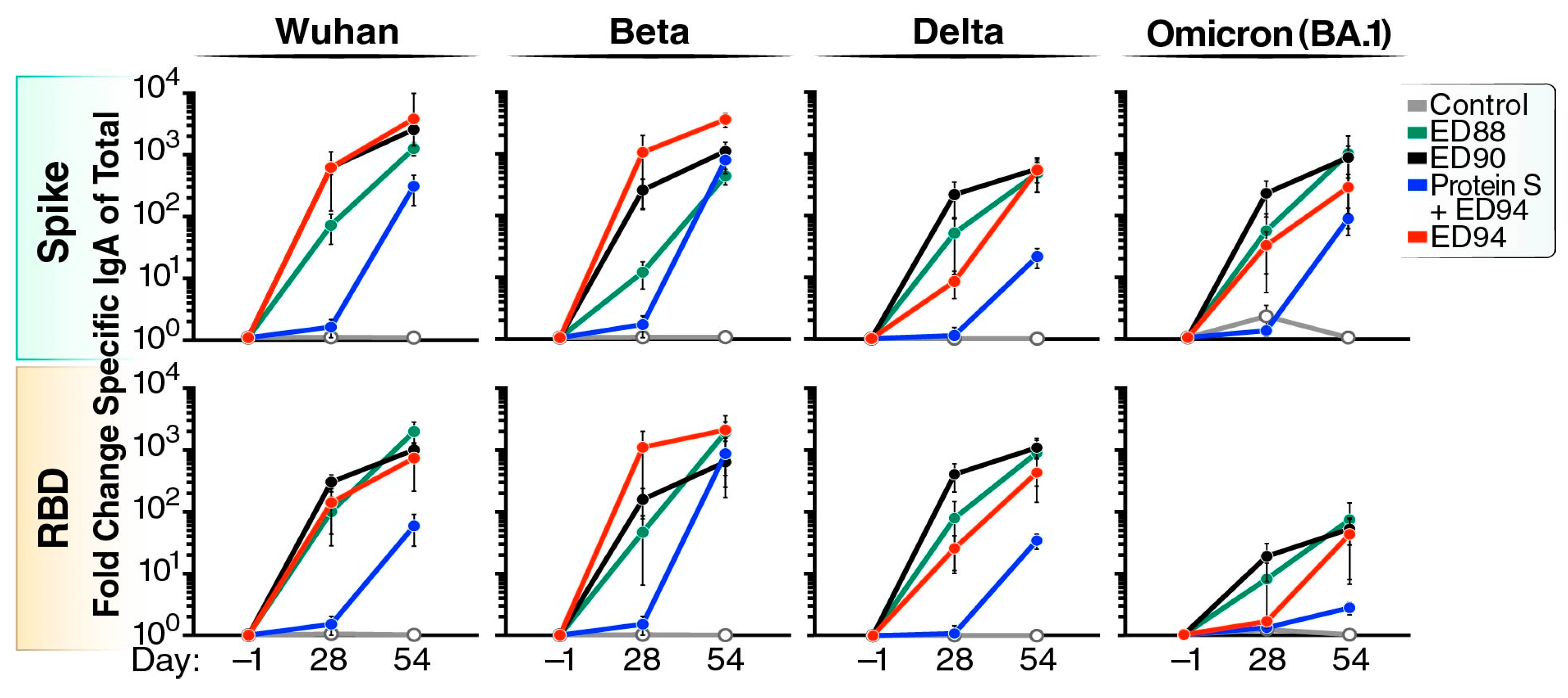
3.2. Robust Cross-Reactive Nasal IgA Is Elicited Following Mucosal Vaccination
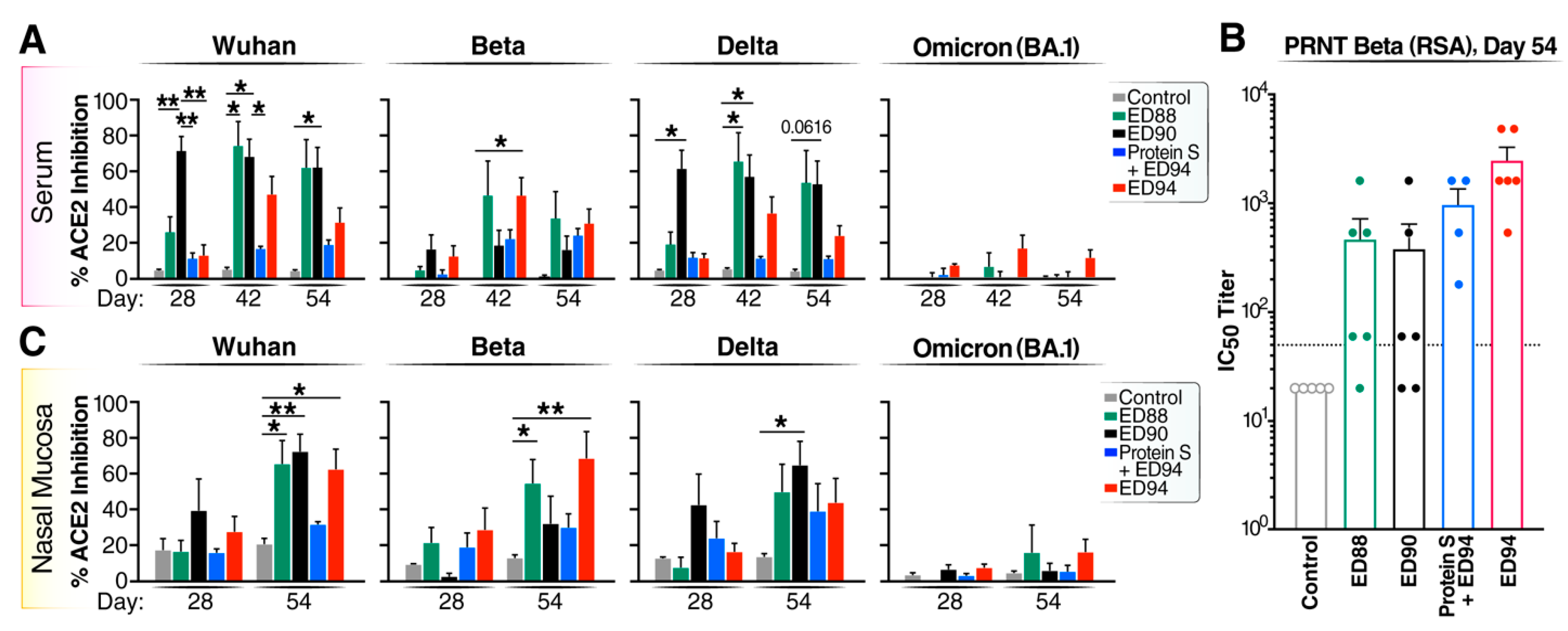
3.3. Boost Immunization Enhances Neutralizing Antibody Activity in Both the Peripheral and Mucosal Compartments
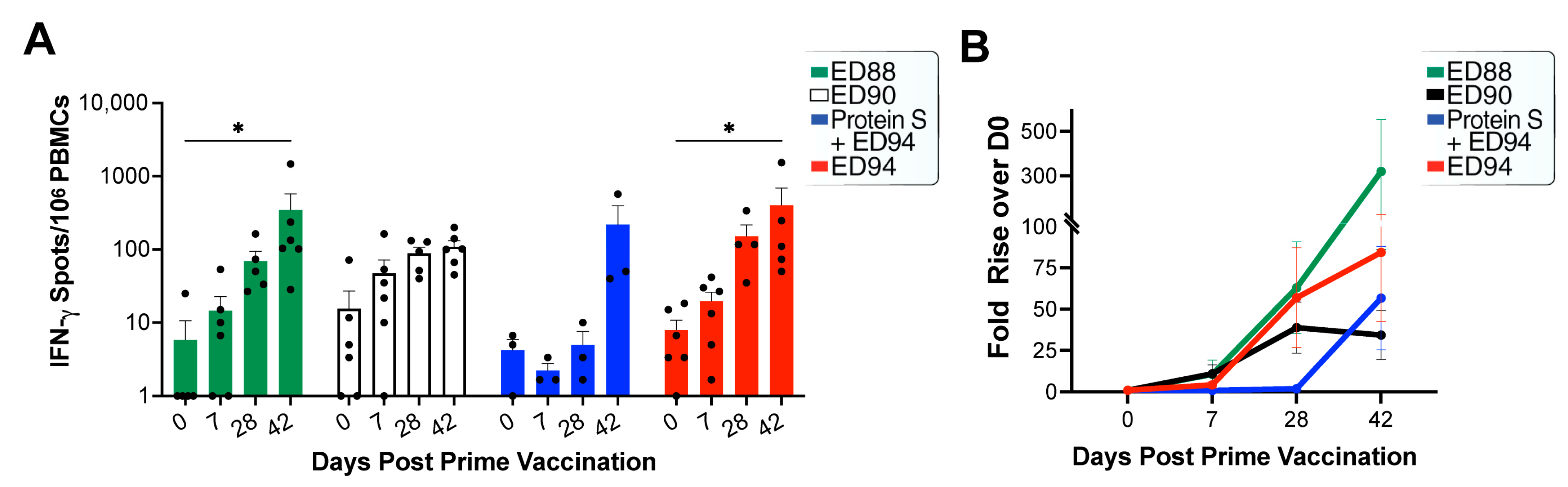
3.4. Mucosal Administration of rAd5 Vaccines Elicits Antigen-Specific T-cells
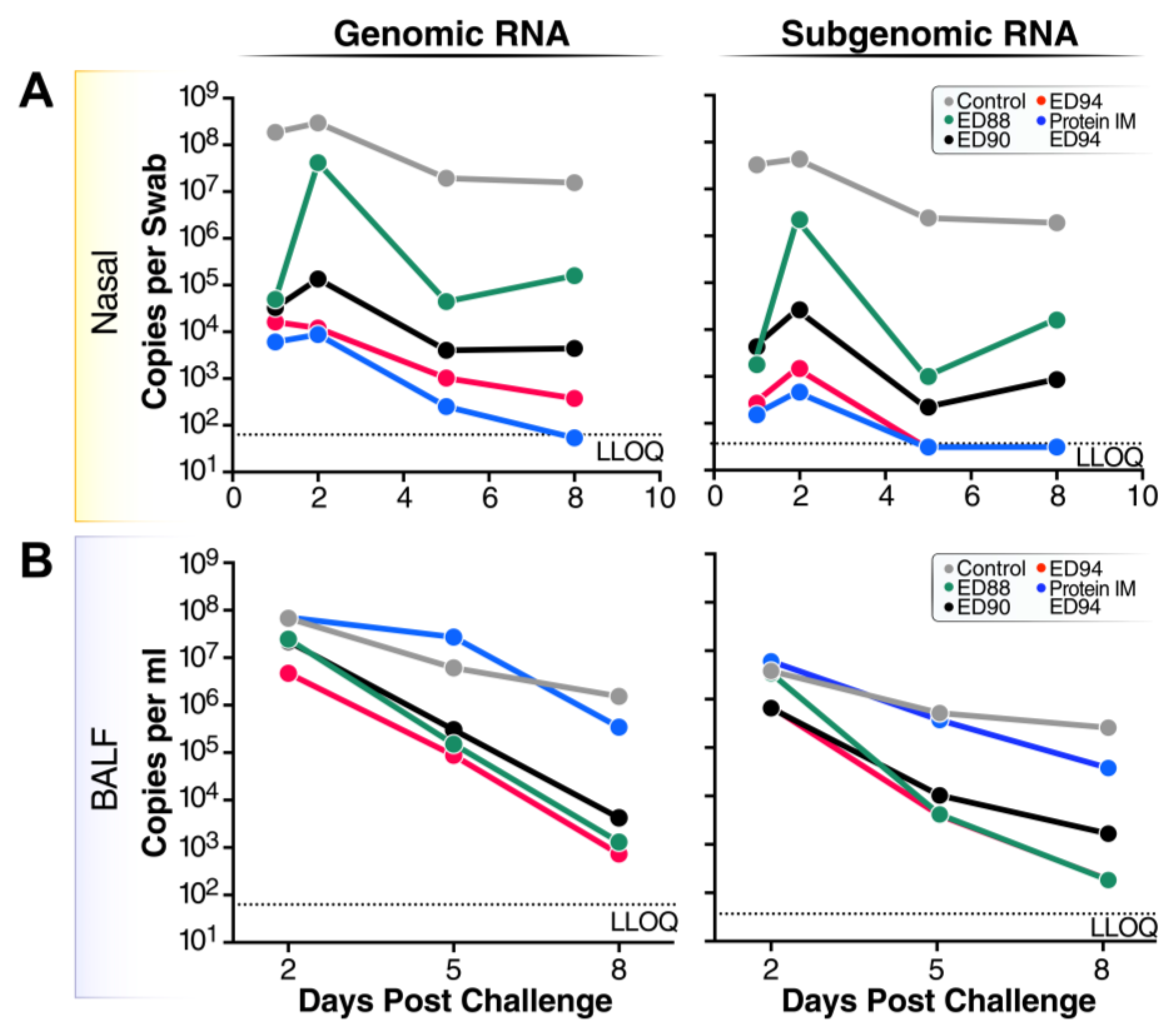
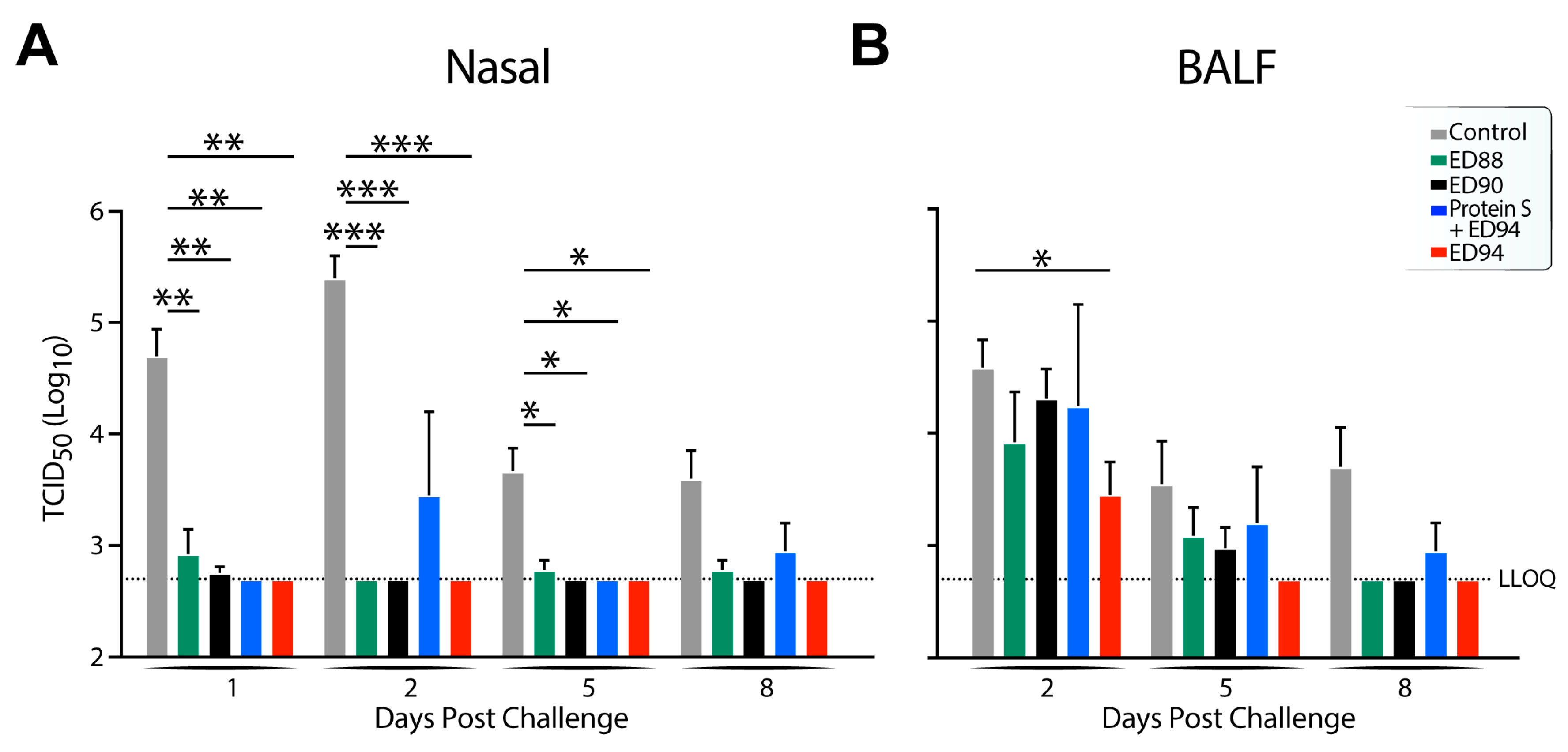
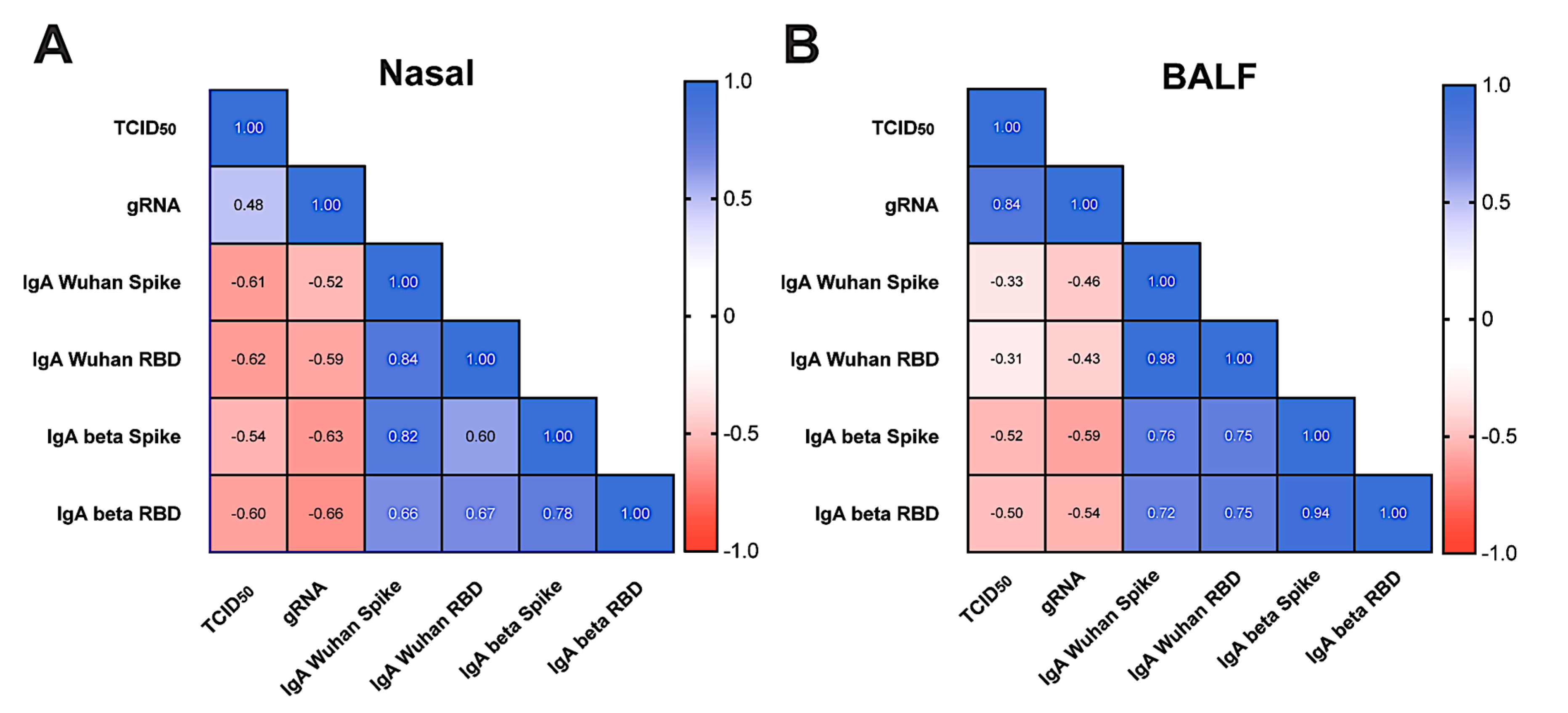
3.5. Viral Replication and Shedding Is Significantly Reduced in Immunized Animals after Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Planas, D.; Staropoli, I.; Porot, F.; Guivel-Benhassine, F.; Handala, L.; Prot, M.; Bolland, W.H.; Puech, J.; Pere, H.; Veyer, D.; et al. Duration of BA.5 neutralization in sera and nasal swabs from SARS-CoV-2 vaccinated individuals, with or without omicron breakthrough infection. Med 2022, 3, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—fortifying the frontiers. Nat. Rev. Immunol. 2022, 22, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Neutra, M.R.; Kozlowski, P.A. Mucosal vaccines: The promise and the challenge. Nat. Rev. Immunol. 2006, 6, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; Logan, N.; Lorenzo, G.D.; Furnon, W.; Scott, S.; Manali, M.; Szemiel, A.; et al. The hyper-transmissible SARS-CoV-2 Omicron variant exhibits significant antigenic change, vaccine escape and a switch in cell entry mechanism. medRxiv 2022. 2022.2001.2003.21268111. [Google Scholar] [CrossRef]
- Buchan, S.A.; Chung, H.; Brown, K.A.; Austin, P.C.; Fell, D.B.; Gubbay, J.B.; Nasreen, S.; Schwartz, K.L.; Sundaram, M.E.; Tadrous, M.; et al. Effectiveness of COVID-19 vaccines against Omicron or Delta infection. medRxiv 2022. 2021.2012.2030.21268565. [Google Scholar] [CrossRef]
- Corthesy, B. Multi-faceted functions of secretory IgA at mucosal surfaces. Front. Immunol. 2013, 4, 185. [Google Scholar] [CrossRef]
- Muramatsu, M.; Yoshida, R.; Yokoyama, A.; Miyamoto, H.; Kajihara, M.; Maruyama, J.; Nao, N.; Manzoor, R.; Takada, A. Comparison of antiviral activity between IgA and IgG specific to influenza virus hemagglutinin: Increased potential of IgA for heterosubtypic immunity. PLoS ONE 2014, 9, e85582. [Google Scholar] [CrossRef]
- Saito, S.; Sano, K.; Suzuki, T.; Ainai, A.; Taga, Y.; Ueno, T.; Tabata, K.; Saito, K.; Wada, Y.; Ohara, Y.; et al. IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody. PLoS Pathog. 2019, 15, e1007427. [Google Scholar] [CrossRef]
- Binsker, U.; Lees, J.A.; Hammond, A.J.; Weiser, J.N. Immune exclusion by naturally acquired secretory IgA against pneumococcal pilus-1. J. Clin. Investig. 2020, 130, 927–941. [Google Scholar] [CrossRef]
- Okuya, K.; Yoshida, R.; Manzoor, R.; Saito, S.; Suzuki, T.; Sasaki, M.; Saito, T.; Kida, Y.; Mori-Kajihara, A.; Kondoh, T.; et al. Potential Role of Nonneutralizing IgA Antibodies in Cross-Protective Immunity against Influenza A Viruses of Multiple Hemagglutinin Subtypes. J. Virol. 2020, 94, 12. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Viant, C.; Gaebler, C.; Cipolla, M.; Hoffmann, H.H.; Oliveira, T.Y.; Oren, D.A.; et al. Enhanced SARS-CoV-2 neutralization by dimeric IgA. Sci. Transl. Med. 2021, 13, eabf1555. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, H.; Cao, Y.; Zuo, F.; Simonelli, L.; Sammartino, J.C.; Pedotti, M.; Sun, R.; Cassaniti, I.; Hagbom, M.; Piralla, A.; et al. Conversion of monoclonal IgG to dimeric and secretory IgA restores neutralizing ability and prevents infection of Omicron lineages. Proc. Natl. Acad. Sci. USA 2024, 121, e2315354120. [Google Scholar] [CrossRef] [PubMed]
- Flitter, B.A.; Braun, M.R.; Tucker, S.N. Drop the Needle; A Temperature Stable Oral Tablet Vaccine Is Protective against Respiratory Viral Pathogens. Vaccines 2022, 10, 593. [Google Scholar] [CrossRef] [PubMed]
- Liebowitz, D.; Gottlieb, K.; Kolhatkar, N.S.; Garg, S.J.; Asher, J.M.; Nazareno, J.; Kim, K.; McIIwain, D.R.; Tucker, S.N. Efficacy, immunogenicity, and safety of an oral influenza vaccine: A placebo-controlled and active-controlled phase 2 human challenge study. Lancet Infect. Dis. 2020, 20, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Liebowitz, D.; Lin, K.; Kasparek, K.; Pasetti, M.F.; Garg, S.J.; Gottlieb, K.; Trager, G.; Tucker, S.N. Safety and immunogenicity of an oral tablet norovirus vaccine, a phase I randomized, placebo-controlled trial. JCI Insight 2018, 3, e121077. [Google Scholar] [CrossRef]
- Liebowitz, D.; Lindbloom, J.D.; Brandl, J.R.; Garg, S.J.; Tucker, S.N. High titre neutralising antibodies to influenza after oral tablet immunisation: A phase 1, randomised, placebo-controlled trial. Lancet Infect. Dis. 2015, 15, 1041–1048. [Google Scholar] [CrossRef]
- Langel, S.N.; Johnson, S.; Martinez, C.I.; Tedjakusuma, S.N.; Peinovich, N.; Dora, E.G.; Kuehl, P.J.; Irshad, H.; Barrett, E.G.; Werts, A.D.; et al. Adenovirus type 5 SARS-CoV-2 vaccines delivered orally or intranasally reduced disease severity and transmission in a hamster model. Sci. Transl. Med. 2022, 14, eabn6868. [Google Scholar] [CrossRef]
- Awasthi, M.; Macaluso, A.; Myscofski, D.; Prigge, J.; Koide, F.; Noyce, R.S.; Fogarty, S.; Stillwell, H.; Goebel, S.J.; Daugherty, B.; et al. Immunogenicity and Efficacy of TNX-1800, A Live Virus Recombinant Poxvirus Vaccine Candidate, against SARS-CoV-2 Challenge in Nonhuman Primates. Vaccines 2023, 11, 1682. [Google Scholar] [CrossRef]
- Flitter, B.A.; Lester, C.A.; Tedjakusuma, S.N.; Dora, E.G.; Peinovich, N.; Cortese, M.C.; Martinez, C.I.; Jegede, C.B.; Neuhaus, E.D.; Tucker, S.N. Mucosal Immunization of Cynomolgus Macaques with Adenoviral Vector Vaccine Elicits Neutralizing Nasal and Serum Antibody to Several SARS-CoV-2 Variants. bioRxiv 2022. [Google Scholar] [CrossRef]
- McAuliffe, J.; Vogel, L.; Roberts, A.; Fahle, G.; Fischer, S.; Shieh, W.J.; Butler, E.; Zaki, S.; St Claire, M.; Murphy, B.; et al. Replication of SARS coronavirus administered into the respiratory tract of African Green, rhesus and cynomolgus monkeys. Virology 2004, 330, 8–15. [Google Scholar] [CrossRef]
- Hartman, A.L.; Nambulli, S.; McMillen, C.M.; White, A.G.; Tilston-Lunel, N.L.; Albe, J.R.; Cottle, E.; Dunn, M.D.; Frye, L.J.; Gilliland, T.H.; et al. SARS-CoV-2 infection of African green monkeys results in mild respiratory disease discernible by PET/CT imaging and shedding of infectious virus from both respiratory and gastrointestinal tracts. PLoS Pathog. 2020, 16, e1008903. [Google Scholar] [CrossRef] [PubMed]
- Woolsey, C.; Borisevich, V.; Prasad, A.N.; Agans, K.N.; Deer, D.J.; Dobias, N.S.; Heymann, J.C.; Foster, S.L.; Levine, C.B.; Medina, L.; et al. Establishment of an African green monkey model for COVID-19 and protection against re-infection. Nat. Immunol. 2021, 22, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Speranza, E.; Williamson, B.N.; Feldmann, F.; Sturdevant, G.L.; Perez-Perez, L.; Meade-White, K.; Smith, B.J.; Lovaglio, J.; Martens, C.; Munster, V.J.; et al. Single-cell RNA sequencing reveals SARS-CoV-2 infection dynamics in lungs of African green monkeys. Sci. Transl. Med. 2021, 13, eabe8146. [Google Scholar] [CrossRef]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef]
- Decru, B.; Van Elslande, J.; Steels, S.; Van Pottelbergh, G.; Godderis, L.; Van Holm, B.; Bossuyt, X.; Van Weyenbergh, J.; Maes, P.; Vermeersch, P. IgG Anti-Spike Antibodies and Surrogate Neutralizing Antibody Levels Decline Faster 3 to 10 Months After BNT162b2 Vaccination Than After SARS-CoV-2 Infection in Healthcare Workers. Front. Immunol. 2022, 13, 909910. [Google Scholar] [CrossRef]
- Blom, K.; Marking, U.; Havervall, S.; Norin, N.G.; Gordon, M.; García, M.; Tecleab, T.; Christ, W.; Forsell, M.; Phillipson, M.; et al. Immune responses after omicron infection in triple-vaccinated health-care workers with and without previous SARS-CoV-2 infection. Lancet Infect. Dis. 2022, 22, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chan, J.F.; Zhou, J.; Wang, M.; Wang, Q.; Zhang, G.; Xu, W.; Chik, K.K.; Zhang, Y.; Wang, Y.; et al. A pan-sarbecovirus vaccine induces highly potent and durable neutralizing antibody responses in non-human primates against SARS-CoV-2 Omicron variant. Cell Res. 2022, 32, 495–497. [Google Scholar] [CrossRef]
- Seibert, C.W.; Rahmat, S.; Krause, J.C.; Eggink, D.; Albrecht, R.A.; Goff, P.H.; Krammer, F.; Duty, J.A.; Bouvier, N.M.; Garcia-Sastre, A.; et al. Recombinant IgA is sufficient to prevent influenza virus transmission in guinea pigs. J. Virol. 2013, 87, 7793–7804. [Google Scholar] [CrossRef]
- McMahan, K.; Wegmann, F.; Aid, M.; Sciacca, M.; Liu, J.; Hachmann, N.P.; Miller, J.; Jacob-Dolan, C.; Powers, O.; Hope, D.; et al. Mucosal boosting enhances vaccine protection against SARS-CoV-2 in macaques. Nature 2023. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.O.; Feldmann, F.; Zhao, H.; Curiel, D.T.; Okumura, A.; Tang-Huau, T.L.; Case, J.B.; Meade-White, K.; Callison, J.; Chen, R.E.; et al. A single intranasal dose of chimpanzee adenovirus-vectored vaccine protects against SARS-CoV-2 infection in rhesus macaques. Cell Rep. Med. 2021, 2, 100230. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccination prevents SARS-CoV-2 pneumonia in rhesus macaques. bioRxiv 2020. [Google Scholar] [CrossRef]
- Monto, A.S.; Ohmit, S.E.; Petrie, J.G.; Johnson, E.; Truscon, R.; Teich, E.; Rotthoff, J.; Boulton, M.; Victor, J.C. Comparative efficacy of inactivated and live attenuated influenza vaccines. N. Engl. J. Med. 2009, 361, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, M.; Ritchie, A.J.; Aboagye, J.; Jenkin, D.; Provstgaad-Morys, S.; Tarbet, I.; Woods, D.; Davies, S.; Baker, M.; Platt, A.; et al. Tolerability and immunogenicity of an intranasally-administered adenovirus-vectored COVID-19 vaccine: An open-label partially-randomised ascending dose phase I trial. EBioMedicine 2022, 85, 104298. [Google Scholar] [CrossRef] [PubMed]
- Altimmune. Altimmune Announces Update On AdCOVID™ Phase 1 Clinical Trial. 29 June 2021. Available online: https://ir.altimmune.com/news-releases/news-release-details/altimmune-announces-update-adcovidtm-phase-1-clinical-trial (accessed on 20 December 2023).
- Singh, C.; Verma, S.; Reddy, P.; Diamond, M.S.; Curiel, D.T.; Chintan, P.; Jain, M.K.; Redkar, S.V.; Bhate, A.S.; Gundappa, V.; et al. Immunogenicity and Tolerability of BBV154 (iNCOVACC®), an Intranasal SARS-CoV-2 Vaccine, Compared with Intramuscular Covaxin® in Healthy Adults: A Randomised, Open-Label, Phase 3 Clinical Trial. Lancet 2023, preprint. [Google Scholar]
- Jin, L.; Tang, R.; Wu, S.; Guo, X.; Hou, L.; Chen, X.; Zhu, T.; Gou, J.; Huang, H.; Zhong, J.; et al. Antibody Persistence and Safety through 6 Months after Heterologous Orally Aerosolised Ad5-nCoV in individuals primed with two-dose CoronaVac previously. medRxiv 2022. [Google Scholar] [CrossRef]
- Lyngse, F.P.; Molbak, K.; Skov, R.L.; Christiansen, L.E.; Mortensen, L.H.; Albertsen, M.; Moller, C.H.; Krause, T.G.; Rasmussen, M.; Michaelsen, T.Y.; et al. Increased transmissibility of SARS-CoV-2 lineage B.1.1.7 by age and viral load. Nat. Commun. 2021, 12, 7251. [Google Scholar] [CrossRef]
- King, J.C., Jr.; Cummings, G.E.; Stoddard, J.; Readmond, B.X.; Magder, L.S.; Stong, M.; Hoffmaster, M.; Rubin, J.; Tsai, T.; Ruff, E.; et al. A pilot study of the effectiveness of a school-based influenza vaccination program. Pediatrics 2005, 116, e868–e873. [Google Scholar] [CrossRef]
- Ambrose, C.S.; Wu, X.; Jones, T.; Mallory, R.M. The role of nasal IgA in children vaccinated with live attenuated influenza vaccine. Vaccine 2012, 30, 6794–6801. [Google Scholar] [CrossRef]
- Solforosi, L.; Costes, L.M.M.; Tolboom, J.T.B.M.; McMahan, K.; Anioke, T.; Hope, D.; Murdza, T.; Sciacca, M.; Bouffard, E.; Barrett, J.; et al. Booster with Ad26.COV2.S or Omicron-adapted vaccine enhanced immunity and efficacy against SARS-CoV-2 Omicron in macaques. Nat. Commun. 2023, 14, 1944. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedjakusuma, S.N.; Lester, C.A.; Neuhaus, E.D.; Dora, E.G.; Gutierrez, S.; Braun, M.R.; Tucker, S.N.; Flitter, B.A. A Next-Generation Adenoviral Vaccine Elicits Mucosal and Systemic Immunogenicity and Reduces Viral Shedding after SARS-CoV-2 Challenge in Nonhuman Primates. Vaccines 2024, 12, 132. https://doi.org/10.3390/vaccines12020132
Tedjakusuma SN, Lester CA, Neuhaus ED, Dora EG, Gutierrez S, Braun MR, Tucker SN, Flitter BA. A Next-Generation Adenoviral Vaccine Elicits Mucosal and Systemic Immunogenicity and Reduces Viral Shedding after SARS-CoV-2 Challenge in Nonhuman Primates. Vaccines. 2024; 12(2):132. https://doi.org/10.3390/vaccines12020132
Chicago/Turabian StyleTedjakusuma, Sarah N., Colin A. Lester, Elena D. Neuhaus, Emery G. Dora, Samanta Gutierrez, Molly R. Braun, Sean N. Tucker, and Becca A. Flitter. 2024. "A Next-Generation Adenoviral Vaccine Elicits Mucosal and Systemic Immunogenicity and Reduces Viral Shedding after SARS-CoV-2 Challenge in Nonhuman Primates" Vaccines 12, no. 2: 132. https://doi.org/10.3390/vaccines12020132