(−)-Loliolide Isolated from Sargassum horneri Protects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes

 ,
,  ,
,  and
and 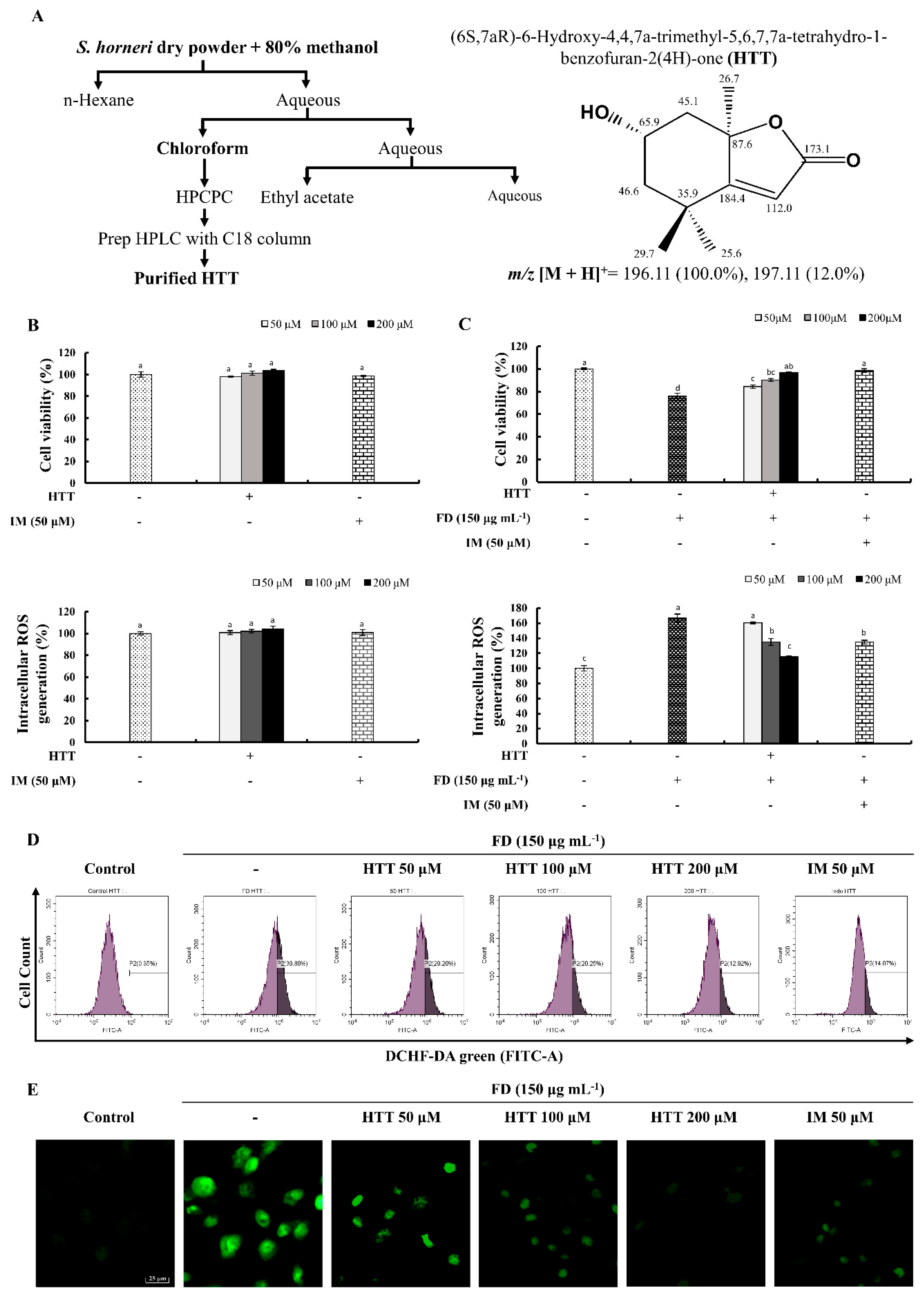{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials, Chemicals, and Reagents
2.2. Sample Preparation and Isolation of HTT
2.3. In Vitro Analysis
2.3.1. Cell Culture and Maintenance
2.3.2. Cell Viability and ROS Production
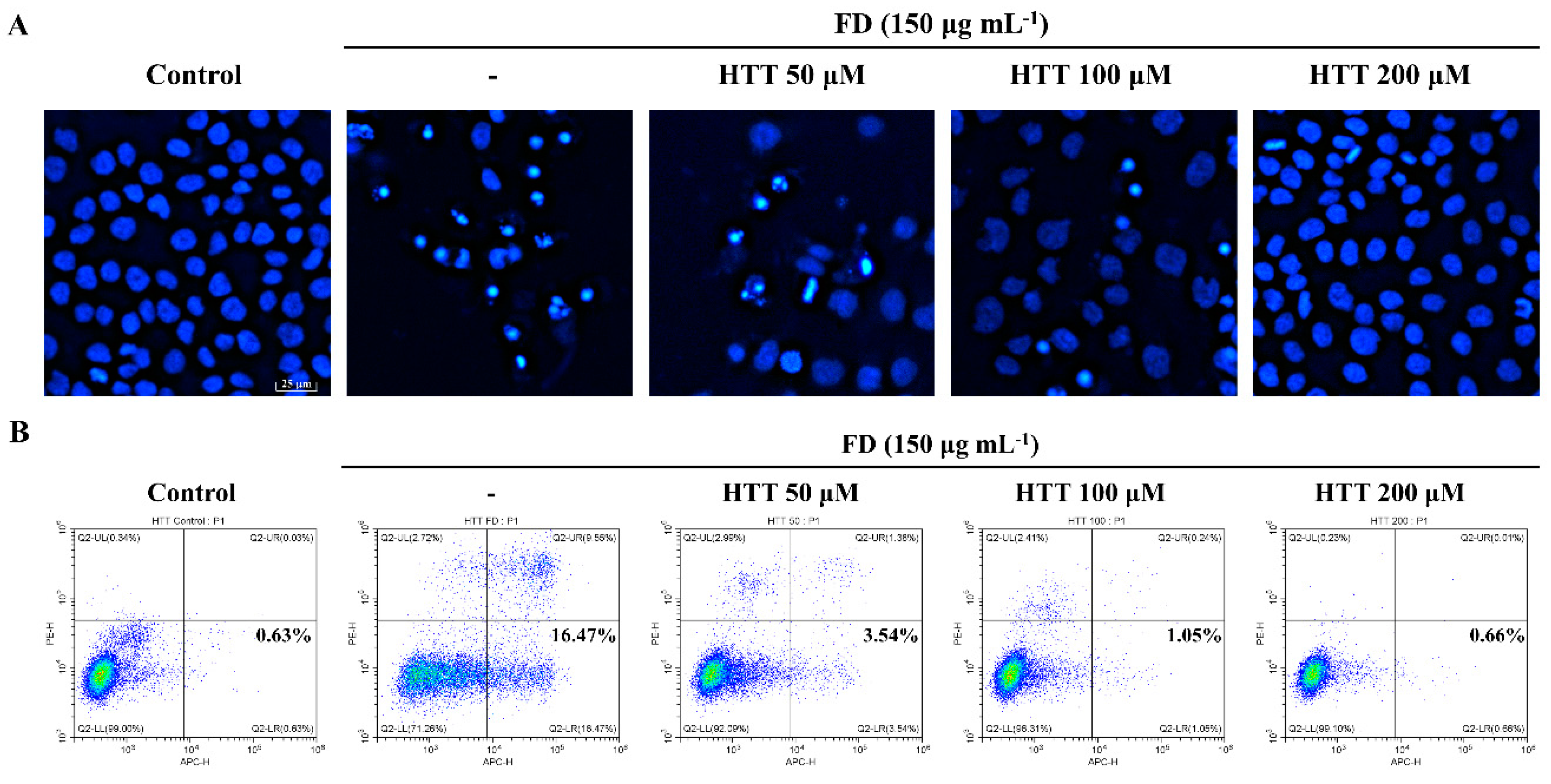
2.3.3. Nuclear Morphological Analysis
2.3.4. Annexin V Assay
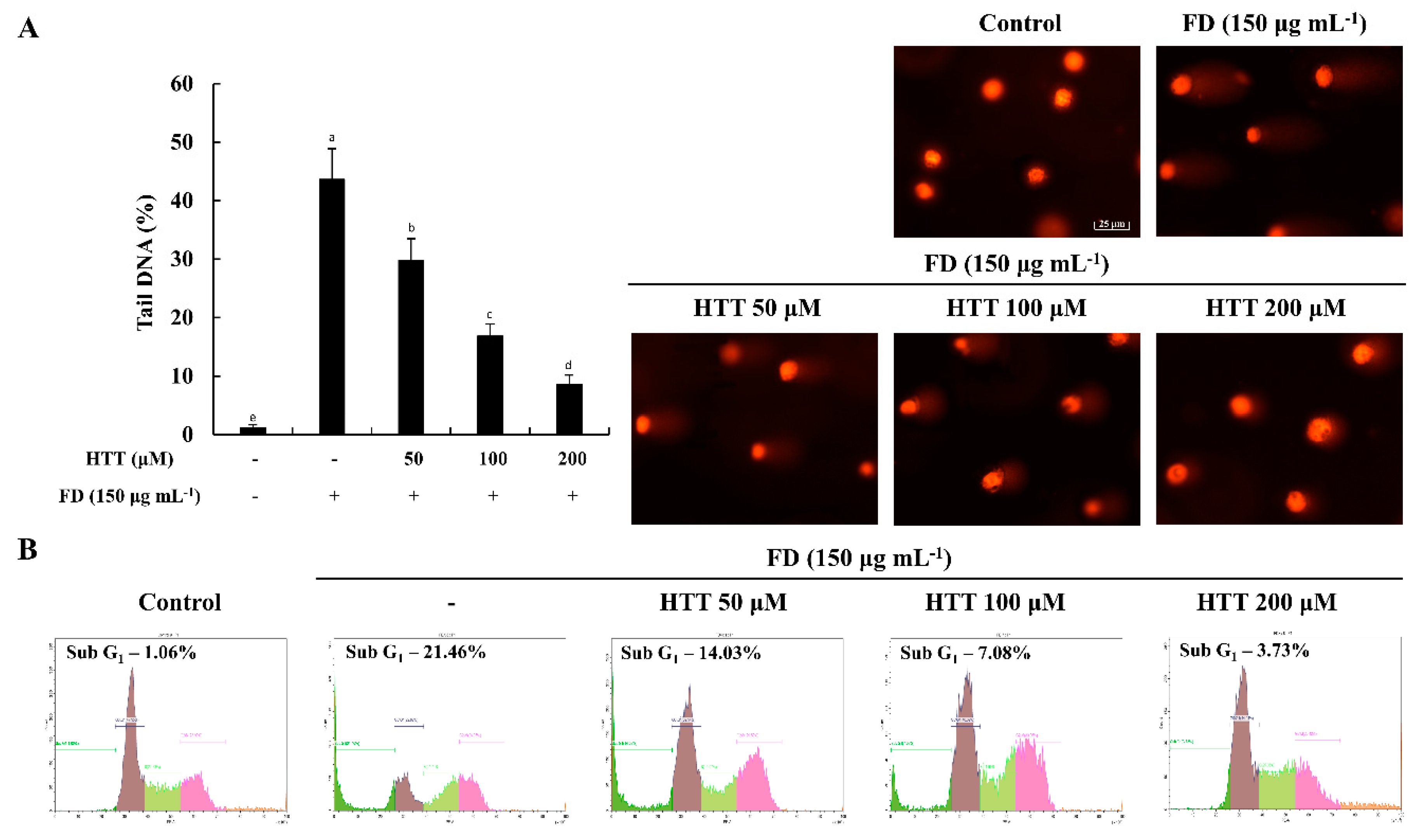
2.3.5. Cell Cycle Analysis
2.3.6. Alkaline Comet Assay
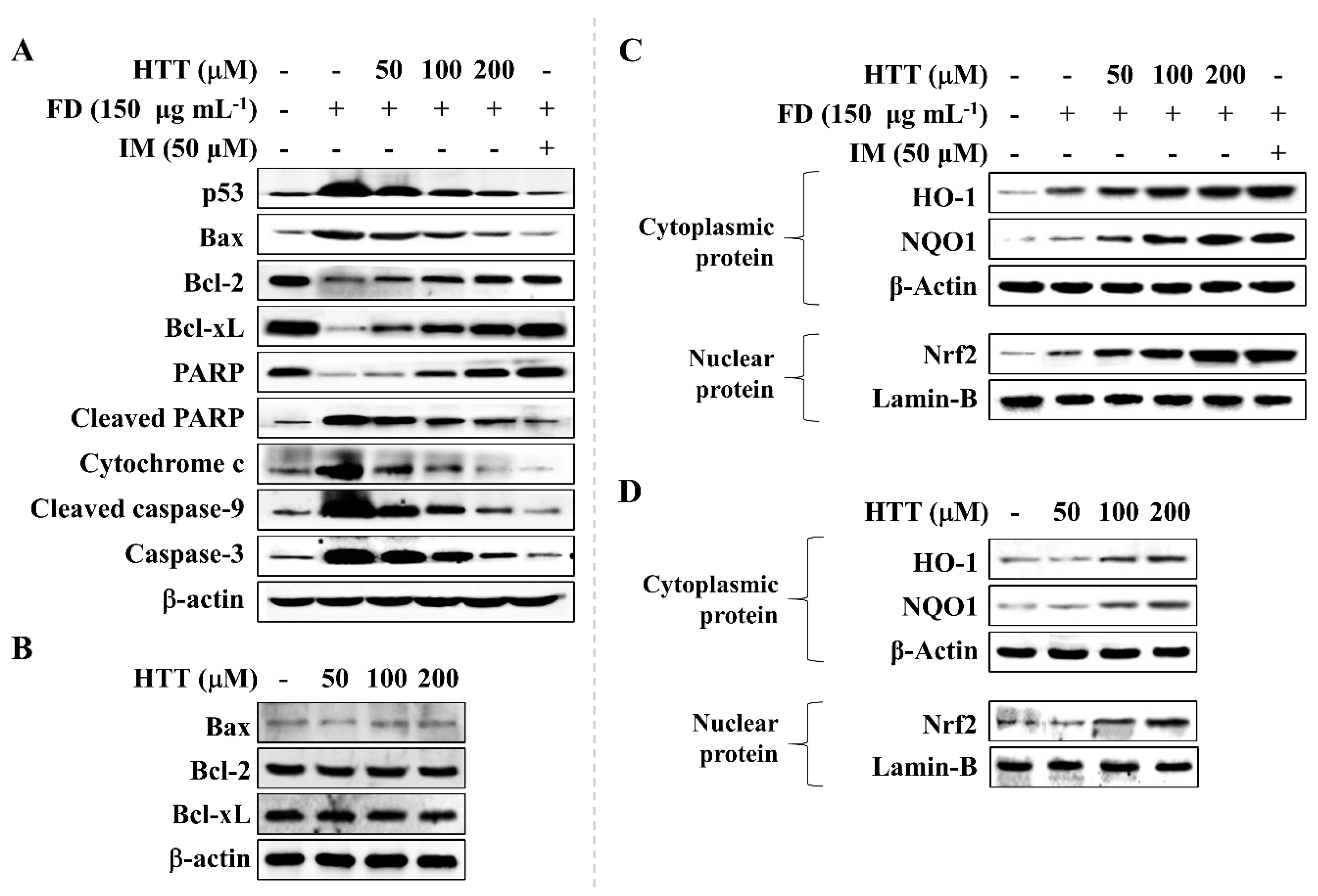
2.3.7. Western Blot Analysis
2.4. Statistical Analysis
3. Results
3.1. Effect of HTT on Cytotoxicity, Cell Viability and Intracellular ROS Production of FD-Induced HaCaT Cells
3.2. HTT Inhibited Early Apoptosis and Apoptotic Body Formation in FD-Induced HaCaT Cells
3.3. HTT Attenuates DNA Damage and Apoptotic Cells in the Sub-G1 Phase
3.4. HTT Attenuates Apoptosis via the Mitochondrial Pathway
3.5. Effect of HTT on Upregulation of Nrf2/HO-1 Pathway Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kang, D.; Kim, J.E. Fine, ultrafine, and yellow dust: Emerging health problems in Korea. J. Korean Med. Sci. 2014, 29, 621–622. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Kim, H.S.; Sanjeewa, K.K.A.; Oh, J.Y.; Jeon, Y.J.; Lee, W.W. Inhibition of inflammatory responses elicited by urban fine dust particles in keratinocytes and macrophages by diphlorethohydroxycarmalol isolated from a brown alga Ishige okamurae. Algae 2017, 32, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Kim, D.S.; Kim, H.; Yi, S.M. Relationship between mortality and fine particles during Asian dust, smog-Asian dust, and smog days in Korea. Int. J. Environ. Health Res. 2012, 22, 518–530. [Google Scholar] [CrossRef] [PubMed]
- VanCuren, R.A.; Cahill, T.A. Asian aerosols in North America: Frequency and concentration of fine dust. J. Geophys. Res. Atmos. 2002, 107, 4804. [Google Scholar] [CrossRef] [Green Version]
- Raloff, J. Ill winds Dust storms ferry toxic agents between countries and even continents. Sci. News 2001, 160, 218–220. [Google Scholar] [CrossRef]
- Choi, H.; Shin, D.W.; Kim, W.; Doh, S.J.; Lee, S.H.; Noh, M. Asian dust storm particles induce a broad toxicological transcriptional program in human epidermal keratinocytes. Toxicol. Lett. 2011, 200, 92–99. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Jeon, Y.J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotox. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef]
- Yoshioka, H.; Ishida, M.; Nishi, K.; Oda, H.; Toyohara, H.; Sugahara, T. Studies on anti-allergic activity of Sargassum horneri extract. J. Funct. Foods 2014, 10, 154–160. [Google Scholar] [CrossRef]
- Wen, Z.S.; Liu, L.J.; OuYang, X.K.; Qu, Y.L.; Chen, Y.; Ding, G.F. Protective effect of polysaccharides from Sargassum horneri against oxidative stress in RAW 264.7 cells. Int. J. Biol. Macromol. 2014, 68, 98–106. [Google Scholar] [CrossRef]
- Kim, H.S.; Sanjeewa, K.K.A.; Fernando, I.P.S.; Ryu, B.; Yang, H.W.; Ahn, G.; Kang, M.C.; Heo, S.T.; Je, J.G.; Jeon, Y.J. A comparative study of Sargassum horneri Korea and China strains collected along the coast of Jeju Island South Korea: Its components and bioactive properties. Algae 2018, 33, 341–349. [Google Scholar] [CrossRef]
- Marks, L.M.; Reed, D.C.; Holbrook, S.J. Life history traits of the invasive seaweed Sargassum horneri at Santa Catalina Island, California. Aquat Invasions 2018, 13, 339–350. [Google Scholar] [CrossRef]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabarczyk, M.; Wińska, K.; Mączka, W.; Potaniec, B.; Anioł, M. Loliolide—The most ubiquitous lactone. Folia Biol. Oecol. 2015, 11, 1–8. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.C.; Lee, K.W.; Kang, S.M.; Lee, W.W.; Jeon, Y.J. Antioxidant activity and cell protective effect of loliolide isolated from Sargassum ringgoldianum subsp. coreanum. Algae 2011, 26, 201–208. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Kim, H.-S.; Sanjeewa, K.K.A.; Kim, S.-Y.; Rho, J.-R.; Jee, Y.; Ahn, G.; Jeon, Y.-J. Sargassum horneri and isolated 6-hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)-one (HTT); LPS-induced inflammation attenuation via suppressing NF-κB, MAPK and oxidative stress through Nrf2/HO-1 pathways in RAW 264.7 macrophages. Algal Res. 2019, 40, 101513. [Google Scholar] [CrossRef]
- Wang, L.; Ryu, B.; Kim, W.S.; Kim, G.H.; Jeon, Y.J. Protective effect of gallic acid derivatives from the freshwater green alga Spirogyra sp. against ultraviolet B-induced apoptosis through reactive oxygen species clearance in human keratinocytes and zebrafish. Algae 2017, 32, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, H.S.; Wang, L.; Lee, W.W.; Jeon, Y.J. Apoptotic and antiproliferative properties of 3β-hydroxy-Δ5-steroidal congeners from a partially purified column fraction of Dendronephthya gigantea against HL-60 and MCF-7 cancer cells. J. Appl. Toxicol. 2017, 38, 527–536. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, A.-C.; Wang, M.-S.; Peng, X. Detection of apoptosis induced by new type gosling viral enteritis virus in vitro through fluorescein annexin V-FITC/PI double labeling. World J. Gastroenterol. 2008, 14, 2174–2178. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Haberzettl, P.; Bhatnagar, A.; Conklin, D.J. Particulate matter and oxidative stress—Pulmonary and cardiovascular targets and consequences. In Systems Biology of Free Radicals and Antioxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1557–1586. [Google Scholar]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Lakey, P.S.J.; Berkemeier, T.; Tong, H.J.; Arangio, A.M.; Lucas, K.; Poschl, U.; Shiraiwa, M. Chemical exposure-response relationship between air pollutants and reactive oxygen species in the human respiratory tract. Sci. Rep. UK 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Bigagli, E. Oxidative stress and air pollution exposure. J. Toxicol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, J.A.; Wang, L.L.; Chen, C.C.; Yang, D.; Jin, M.L.; Bai, C.X.; Song, Y.L. Urban particulate matter triggers lung inflammation via the ROS-MAPK- NF-kappa B signaling pathway. J. Thorac Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, I.; Sun, Z.; Ukachi, M.; Nagano, K.; McLeod, C.W.; Cox, A.G.; Nishikawa, M. Development and certification of the new NIES CRM 28: Urban aerosols for the determination of multielements. Anal. Bioanal. Chem. 2008, 391, 1997–2003. [Google Scholar] [CrossRef] [PubMed]
- Piaścik, M.; Przyk, E.P.; Held, A. The Certification of the Mass Fractions of Selected Polycyclic Aromatic Hydrocarbons (PAHs) in Fine Dust (PM10-Like Matrix) Certified Reference Material ERM®-CZ100; Institute for Reference Materials and Measurements: Geel, Belgium; Publications Office of the European Union: Brussels, Belgium, 2010; pp. 1–50. [Google Scholar]
- Mitkus, R.J.; Powell, J.L.; Zeisler, R.; Squibb, K.S. Comparative physicochemical and biological characterization of NIST Interim Reference Material PM2.5 and SRM 1648 in human A549 and mouse RAW264.7 cells. Toxicol. In Vitro 2013, 27, 2289–2298. [Google Scholar] [CrossRef]
- Kijima, M.; Mizuta, R. Histone H1 quantity determines the efficiencies of apoptotic DNA fragmentation and chromatin condensation. Biomed. Res. Tokyo 2019, 40, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Kajstura, M.; Halicka, H.D.; Pryjma, J.; Darzynkiewicz, Z. Discontinuous fragmentation of nuclear DNA during apoptosis revealed by discrete "sub-G(1)" peaks on DNA content histograms. Cytom. Part. A 2007, 71, 125–131. [Google Scholar] [CrossRef]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.F.; Shen, X.Y.; Lio, C.K.; Dai, Y.; Cheng, C.S.; Liu, J.X.; Yao, Y.D.; Yu, Y.; Xie, Y.; Luo, P.; et al. Activation of Nrf2/HO-1 Pathway by Nardochinoid C Inhibits Inflammation and Oxidative Stress in Lipopolysaccharide-Stimulated Macrophages. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Jeon, Y.-J.; Kim, H.-S.; Fernando, I.P.S.; Ahn, G. (−)-Loliolide Isolated from Sargassum horneri Protects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes. Antioxidants 2020, 9, 474. https://doi.org/10.3390/antiox9060474
Dias MKHM, Madusanka DMD, Han EJ, Kim MJ, Jeon Y-J, Kim H-S, Fernando IPS, Ahn G. (−)-Loliolide Isolated from Sargassum horneri Protects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes. Antioxidants. 2020; 9(6):474. https://doi.org/10.3390/antiox9060474
Chicago/Turabian StyleDias, Mawalle Kankanamge Hasitha Madhawa, Dissanayaka Mudiyanselage Dinesh Madusanka, Eui Jeong Han, Min Ju Kim, You-Jin Jeon, Hyun-Soo Kim, Ilekuttige Priyan Shanura Fernando, and Ginnae Ahn. 2020. "(−)-Loliolide Isolated from Sargassum horneri Protects against Fine Dust-Induced Oxidative Stress in Human Keratinocytes" Antioxidants 9, no. 6: 474. https://doi.org/10.3390/antiox9060474