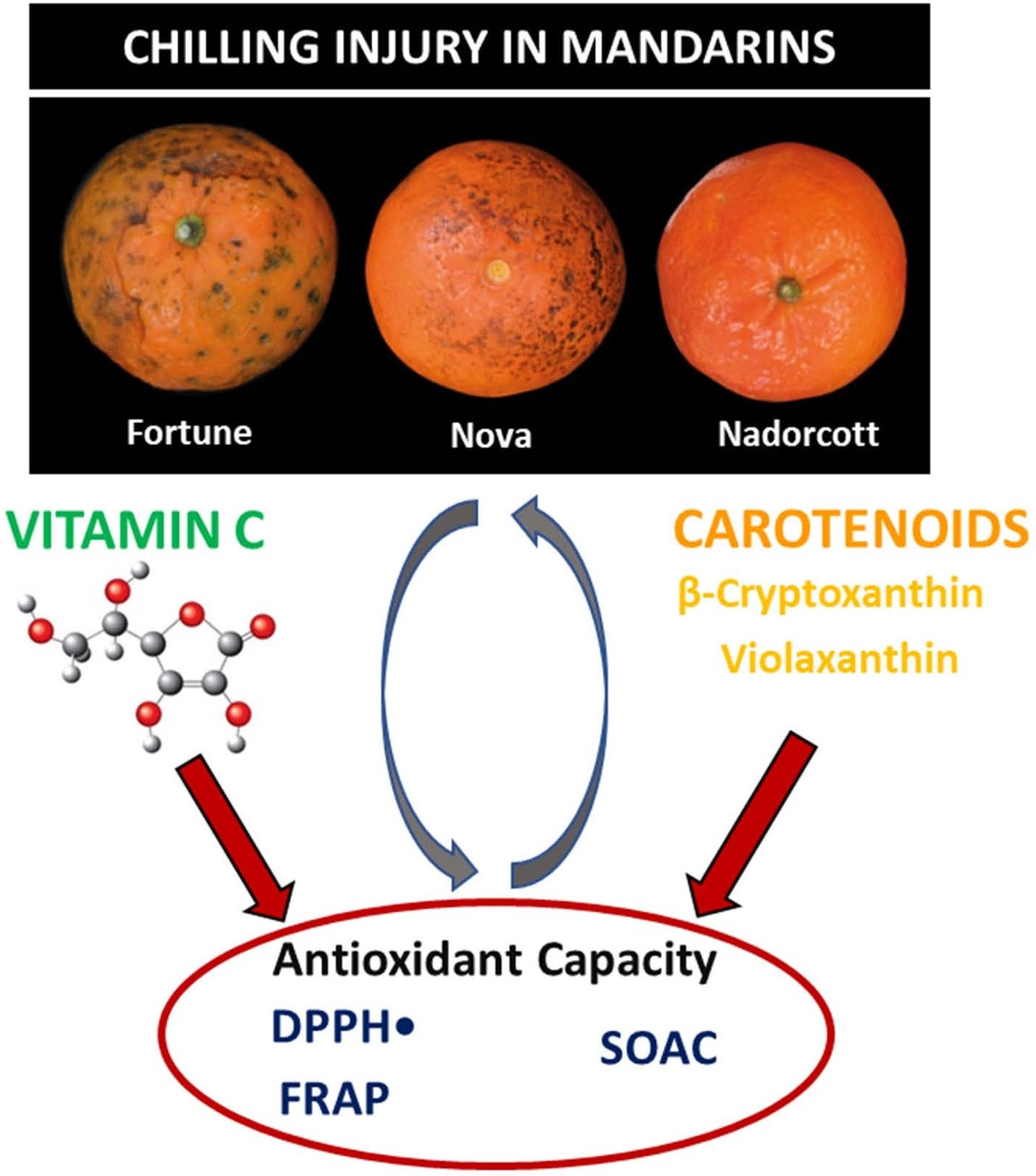
Carotenoids, Vitamin C, and Antioxidant Capacity in the Peel of Mandarin Fruit in Relation to the Susceptibility to Chilling Injury during Postharvest Cold Storage


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Storage Conditions
2.2. Chilling Injury Evaluation
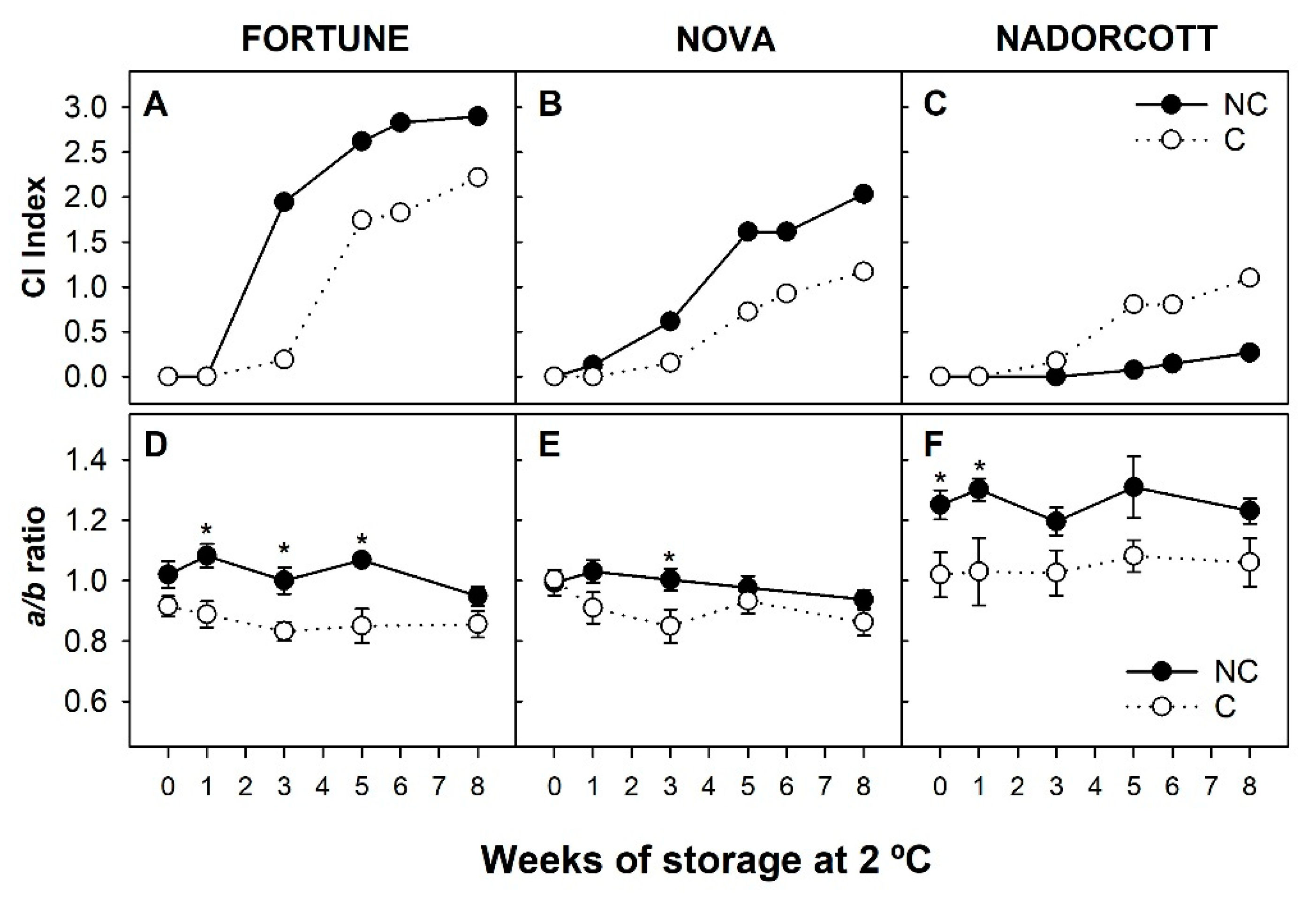
2.3. External Peel Color Evaluation
2.4. Carotenoid Extraction and Analysis
2.5. Vitamin C Determination
2.6. Antioxidants Assays
2.6.1. DPPH and FRAP Assays
2.6.2. Singlet Oxygen Absorption Capacity (SOAC)
2.7. Statistical Analyses
3. Results
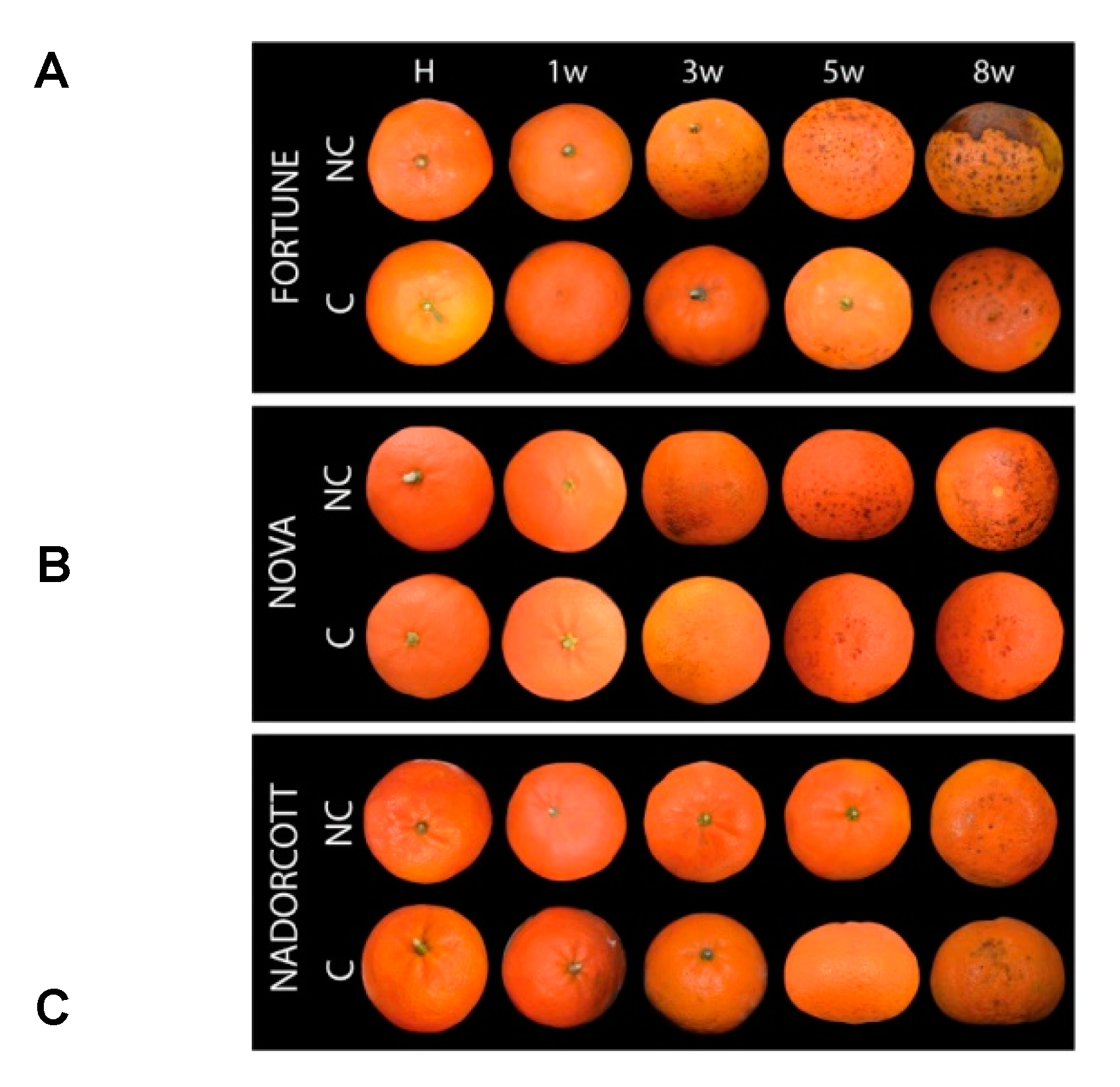
3.1. Incidence of Chilling Injury and Changes in Peel Pigmentation of Fortune, Nova and Nadorcott Mandarins during Cold Storage
3.2. Changes in Carotenoids and Vitamin C in the Peel of Fortune, Nova, and Nadorcott Mandarins during Cold Storage
3.2.1. Total Carotenoid Content and Carotenoid Composition
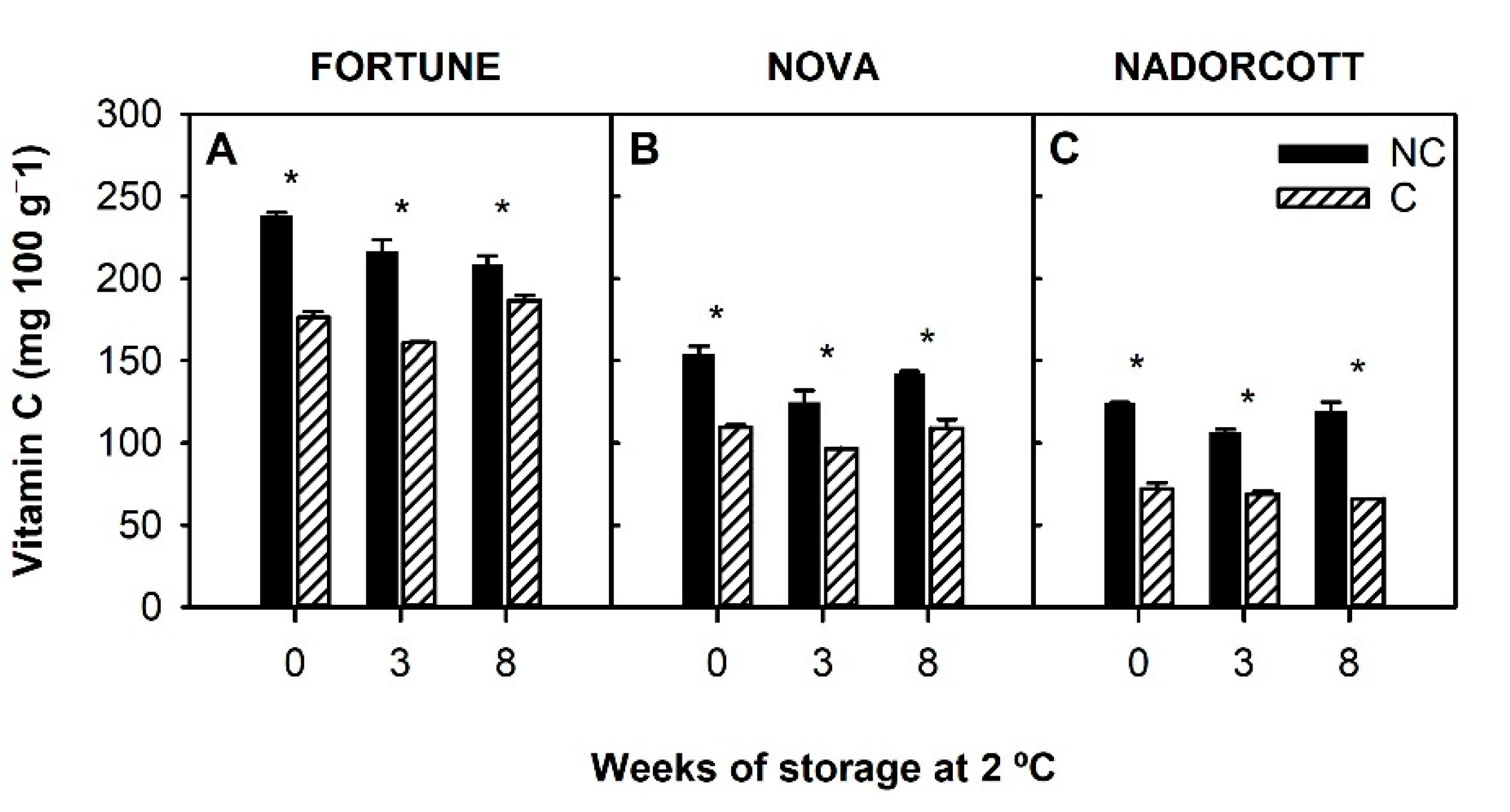
3.2.2. Vitamin C Content
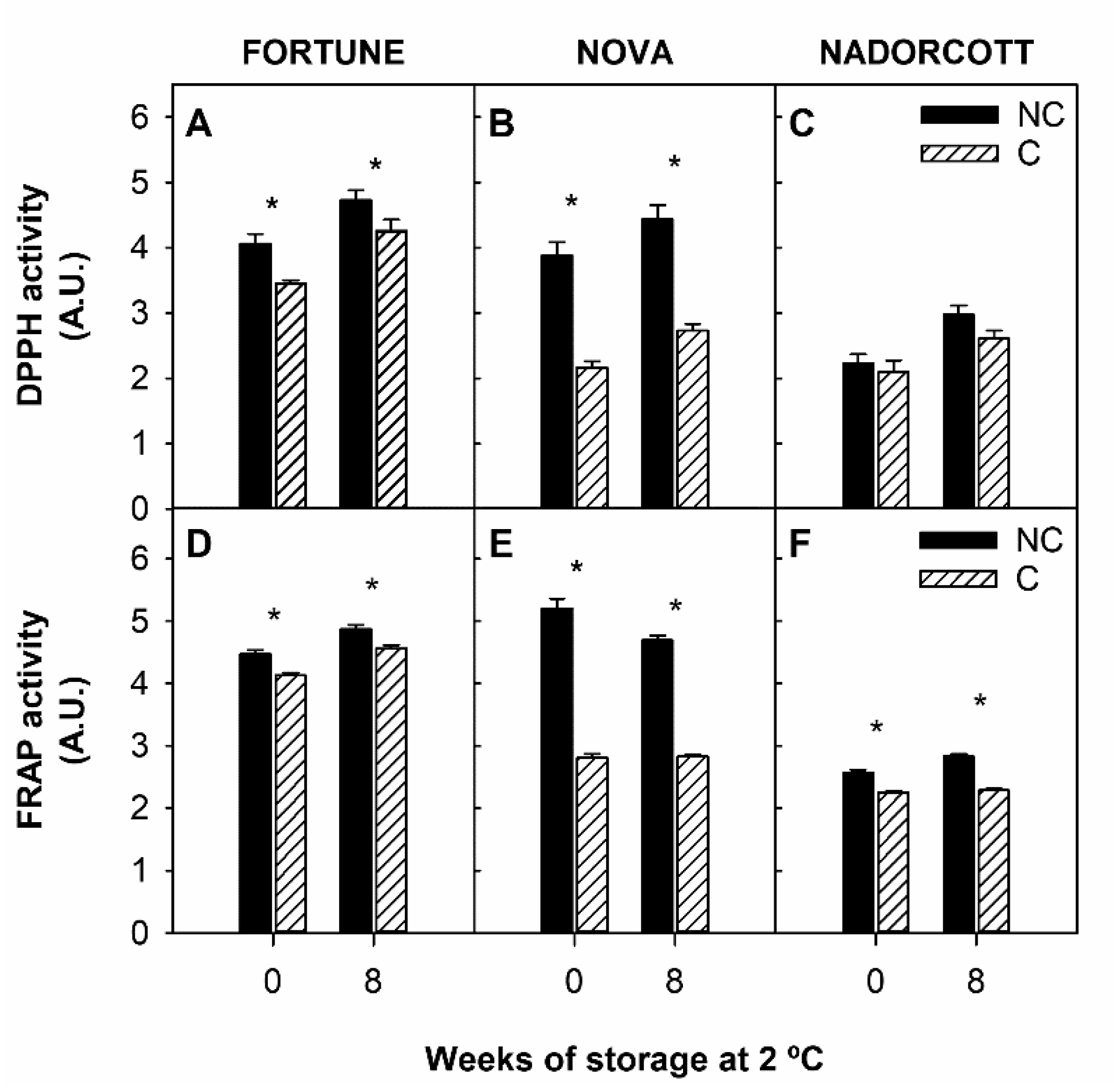
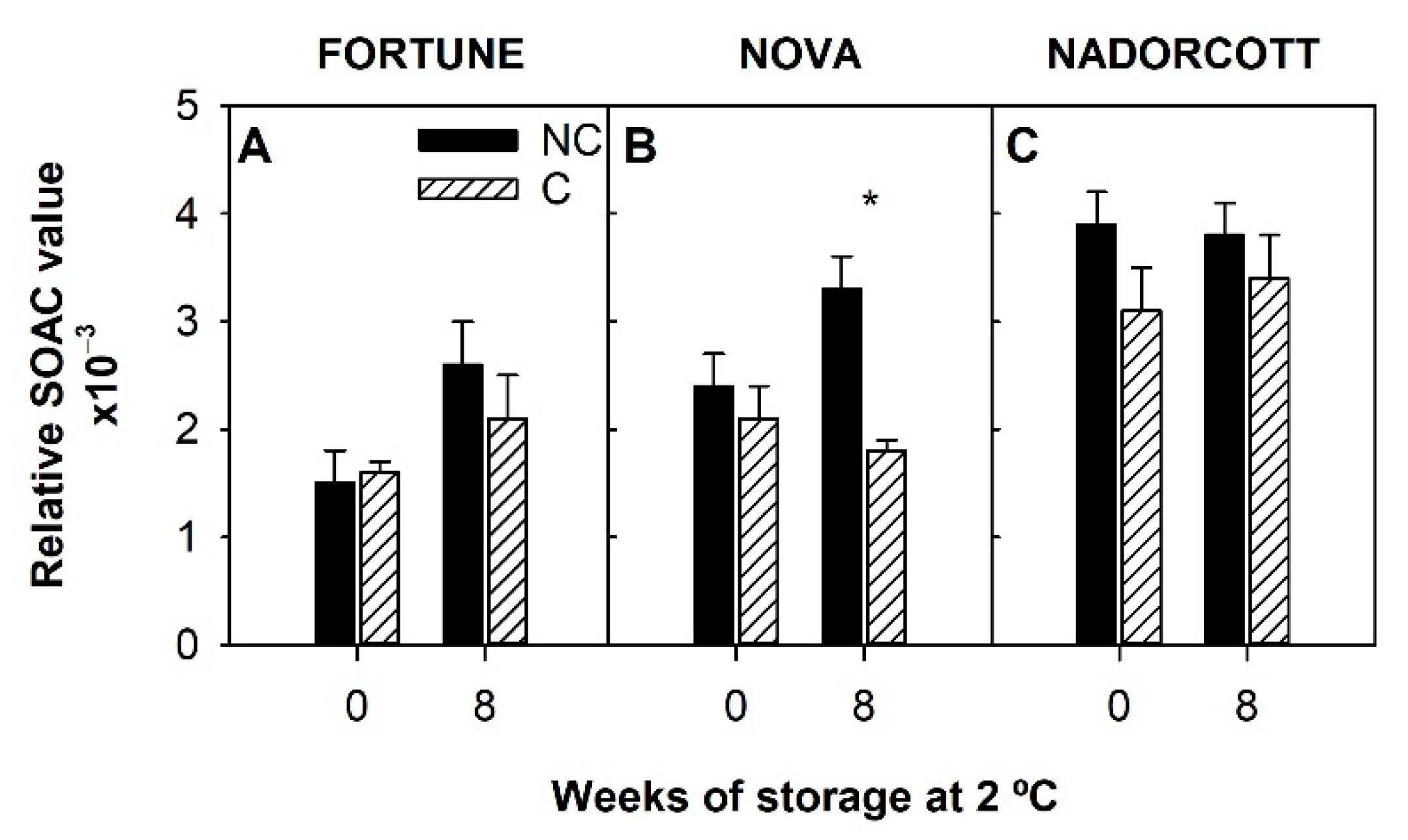
3.3. Antioxidant Activity and Singlet Oxygen Absorption Capacity in the Peel of Fortune, Nova, and Nadorcott Mandarins during Cold Storage
3.4. Correlation Analysis between Chilling Injury Index, Carotenoids, Vitamin C and DPPH•, FRAP and SOAC Antioxidant Activity/Capacity in the Peel of Fortune, Nova, and Nadorcott during Cold Storage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lafuente, M.T.; Zacarías, L. Postharvest physiological disorders in citrus fruit. Stewart Postharvest Rev. 2006, 2, 1–9. [Google Scholar] [CrossRef]
- Zacarias, L.; Cronje, P.J.R.; Palou, L. Postharvest technology of citrus fruits. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 421–446. ISBN 978-0-12-812163-4. [Google Scholar]
- Lado, J.; Cronje, P.J.; Rodrigo, M.J.; Zacarías, L. Citrus. In Postharvest Physiological Disorders in Fruits and Vegetables; Sergio, T.D.F., Sunil, P., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2019; pp. 377–398. ISBN 9781315267470. [Google Scholar]
- Cronje, P.J.R.; Barry, G.H.; Huysamer, M. Postharvest rind breakdown of ‘Nules Clementine’ mandarin is influenced by ethylene application, storage temperature and storage duration. Postharvest Biol. Technol. 2011, 60, 192–201. [Google Scholar] [CrossRef]
- Lado, J.; Rodrigo, M.J.; Cronje, P.; Zacarías, L. Involvement of lycopene in the induction of tolerance to chilling injury in grapefruit. Postharvest Biol. Technol. 2015, 100, 176–186. [Google Scholar] [CrossRef]
- Cronje, P.J.R.; Barry, G.H.; Huysamer, M. Canopy Position Affects Pigment Expression and Accumulation of Flavedo Carbohydrates of ‘Nules Clementine’ Mandarin Fruit, Thereby Affecting Rind Condition. J. Am. Soc. Hortic. Sci. 2013, 138, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Magwaza, L.S.; Opara, U.L.; Cronje, P.J.R.; Landahl, S.; Terry, L.A. Canopy position affects rind biochemical profile of “Nules Clementine” mandarin fruit during postharvest storage. Postharvest Biol. Technol. 2013, 86, 300–308. [Google Scholar] [CrossRef]
- Holland, N.; Menezes, H.C.; Lafuente, M.T. Carbohydrate Metabolism as Related to High-Temperature Conditioning and Peel Disorders Occurring during Storage of Citrus Fruit. J. Agric. Food Chem. 2005, 53, 8790–8796. [Google Scholar] [CrossRef]
- Holland, N.; Menezes, H.C.; Lafuente, M.T. Carbohydrates as related to the heat-induced chilling tolerance and respiratory rate of ‘Fortune’ mandarin fruit harvested at different maturity stages. Postharvest Biol. Technol. 2002, 25, 181–191. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Alquézar, B.; Alós, E.; Lado, J.; Zacarías, L. Biochemical bases and molecular regulation of pigmentation in the peel of Citrus fruit. Sci. Hortic. 2013, 163, 46–62. [Google Scholar] [CrossRef]
- Kato, M. Mechanism of Carotenoid Accumulation in Citrus Fruit. J. Jpn. Soc. Hortic. Sci. 2012, 81, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Llorente, B.; D’Andrea, L.; Ruiz-Sola, M.A.; Botterweg, E.; Pulido, P.; Andilla, J.; Loza-Alvarez, P.; Rodriguez-Concepcion, M. Tomato fruit carotenoid biosynthesis is adjusted to actual ripening progression by a light-dependent mechanism. Plant J. 2016, 85, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Lado, J.; Alós, E.; Manzi, M.; Cronje, P.J.R.; Gómez-Cadenas, A.; Rodrigo, M.J.; Zacarías, L. Light Regulation of Carotenoid Biosynthesis in the Peel of Mandarin and Sweet Orange Fruits. Front. Plant Sci. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2012, 60, 197–201. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, G.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ohta, S.; Kato, M. Effect of blue LED light intensity on carotenoid accumulation in citrus juice sacs. J. Plant Physiol. 2015, 188, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Lado, J.; Cronje, P.; Alquézar, B.; Page, A.; Manzi, M.; Gómez-Cadenas, A.; Stead, A.D.; Zacarías, L.; Rodrigo, M.J. Fruit shading enhances peel color, carotenes accumulation and chromoplast differentiation in red grapefruit. Physiol. Plant. 2015, 154, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevillano, L.; Sanchez-Ballest, M.T.; Romojaro, F.; Flores, F.B. Physiological, hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. J. Sci. Food Agric. 2009, 89, 555–573. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Cantrell, A.; McGarvey, D.; George Truscott, T.; Rancan, F.; Böhm, F. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch. Biochem. Biophys. 2003, 412, 47–54. [Google Scholar] [CrossRef]
- Lado, J.; Rodrigo, M.J.; López-Climent, M.; Gómez-Cadenas, A.; Zacarías, L. Implication of the antioxidant system in chilling injury tolerance in the red peel of grapefruit. Postharvest Biol. Technol. 2016, 111, 214–223. [Google Scholar] [CrossRef]
- Smirnoff, N. Vitamin C: The Metabolism and Functions of Ascorbic Acid in Plants. In Advances in Botanical Research; Rébeillé, F., Douce, R., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 59, pp. 107–177. ISBN 9780123858535. [Google Scholar]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Lado, J.; Alós, E.; Rodrigo, M.J.; Zacarías, L. Light avoidance reduces ascorbic acid accumulation in the peel of Citrus fruit. Plant Sci. 2015, 231, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Alós, E.; Rodrigo, M.J.; Zacarías, L. Differential transcriptional regulation of l-ascorbic acid content in peel and pulp of citrus fruits during development and maturation. Planta 2014, 239, 1113–1128. [Google Scholar] [CrossRef]
- Magwaza, L.S.; Mditshwa, A.; Tesfay, S.Z.; Opara, U.L. An overview of preharvest factors affecting vitamin C content of citrus fruit. Sci. Hortic. 2017, 216, 12–21. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Bassal, M.; El-Hamahmy, M. Hot water dip and preconditioning treatments to reduce chilling injury and maintain postharvest quality of Navel and Valencia oranges during cold quarantine. Postharvest Biol. Technol. 2011, 60, 186–191. [Google Scholar] [CrossRef]
- Erkan, M.; Pekmezci, M.; Karaşahin, I.; Uslu, H. Reducing chilling injury and decay in stored “Clementine” mandarins with hot water and curing treatments. Eur. J. Hortic. Sci. 2005, 70, 183–188. [Google Scholar] [CrossRef]
- Erkan, M.; Pekmezci, M.; Wang, C.Y. Hot water and curing treatments reduce chilling injury and maintain post-harvest quality of “Valencia” oranges. Int. J. Food Sci. Technol. 2005, 40, 91–96. [Google Scholar] [CrossRef]
- Chaudhary, P.R.; Jayaprakasha, G.K.; Porat, R.; Patil, B.S. Low temperature conditioning reduces chilling injury while maintaining quality and certain bioactive compounds of ‘Star Ruby’ grapefruit. Food Chem. 2014, 153, 243–249. [Google Scholar] [CrossRef]
- Yang, X.Y.; Xie, J.X.; Wang, F.F.; Zhong, J.; Liu, Y.Z.; Li, G.H.; Peng, S.A. Comparison of ascorbate metabolism in fruits of two citrus species with obvious difference in ascorbate content in pulp. J. Plant Physiol. 2011, 168, 2196–2205. [Google Scholar] [CrossRef] [PubMed]
- Lafuente, M.T.; Martínez-Téllez, M.A.; Zacarías, L. Abscisic acid in the response of ‘Fortune’ mandarins to chilling. Effect of maturity and high-temperature conditioning. J. Sci. Food Agric. 1997, 73, 494–502. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Cilla, A.; Barberá, R.; Zacarías, L. Carotenoid bioaccessibility in pulp and fresh juice from carotenoid-rich sweet oranges and mandarins. Food Funct. 2015, 6, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J.; Lado, J.; Alós, E.; Alquézar, B.; Dery, O.; Hirschberg, J.; Zacarías, L. A mutant allele of ζ-carotene isomerase (Z-ISO) is associated with the yellow pigmentation of the “Pinalate” sweet orange mutant and reveals new insights into its role in fruit carotenogenesis. BMC Plant Biol. 2019, 19, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.W.; Montagu, M.V.; Inzé, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.; Strain, J.J.; Favell, D.; Fletcher, J. PlantL-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Parra-Rivero, O.; Paes de Barros, M.; Prado, M.d.M.; Gil, J.-V.; Hornero-Méndez, D.; Zacarías, L.; Rodrigo, M.J.; Limón, M.C.; Avalos, J. Neurosporaxanthin Overproduction by Fusarium fujikuroi and Evaluation of Its Antioxidant Properties. Antioxidants 2020, 9, 528. [Google Scholar] [CrossRef]
- Ouchi, A.; Aizawa, K.; Iwasaki, Y.; Inakuma, T.; Terao, J.; Nagaoka, S.I.; Mukai, K. Kinetic study of the quenching reaction of singlet oxygen by carotenoids and food extracts in solution. development of a singlet oxygen absorption capacity (SOAC) assay method. J. Agric. Food Chem. 2010, 58, 9967–9978. [Google Scholar] [CrossRef]
- Takahashi, S.; Iwasaki-Kino, Y.; Aizawa, K.; Terao, J.; Mukai, K. Development of singlet oxygen absorption capacity (SOAC) assay method using a microplate reader. J. AOAC Int. 2016, 99, 193–197. [Google Scholar] [CrossRef]
- Sala, J.M. Involvement of oxidative stress in chilling injury in cold-storedmandarin fruits. Postharvest Biol. Technol. 1998, 13, 255–261. [Google Scholar] [CrossRef]
- Cronjé, P.J.R. Postharvest rind disorders of “Nadorcott” mandarin are affected by rootstock in addition to postharvest treatments. Acta Hortic. 2013, 1007, 111–117. [Google Scholar] [CrossRef]
- Barry, G.H.; Caruso, M.; Gmitter, F.G. Commercial scion varieties. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 83–104. ISBN 978-0-12-812163-4. [Google Scholar]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids, Volume 1B: Spectroscopy; Carotenoids Series; Birkhäuser: Basel, Switzerland, 1994; ISBN 9783764329082. [Google Scholar]
- Magwaza, L.S.; Opara, U.L.; Cronje, P.J.R.; Landahl, S.; Nieuwoudt, H.H.; Mouazen, A.M.; Nicolaï, B.M.; Terry, L.A. Assessment of rind quality of ‘Nules Clementine’ mandarin during postharvest storage: 1. Vis/NIRS PCA models and relationship with canopy position. Sci. Hortic. 2014, 165, 410–420. [Google Scholar] [CrossRef]
- Li, Y.; Golding, J.B.; Arcot, J.; Wills, R.B.H. Continuous exposure to ethylene in the storage environment adversely affects ‘Afourer’ mandarin fruit quality. Food Chem. 2018, 242, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Lado, J.; Cuellar, F.; Rodrigo, M.J.; Zacarías, L. Nutritional Composition of Mandarins. In Nutritional Composition of Fruit Cultivars; Elsevier: Amsterdam, The Netherlands, 2016; pp. 419–443. ISBN 9780124081178. [Google Scholar]
- Alquézar, B.; Rodrigo, M.J.; Zacarías, L. Carotenoid biosynthesis and their regulation in Citrus fruits. Tree For. Sci. Biotechnol. 2008, 2, 23–35. [Google Scholar]
- Rodrigo, M.J.; Alquézar, B.; Alós, E.; Medina, V.; Carmona, L.; Bruno, M.; Al-Babili, S.; Zacarías, L. A novel carotenoid cleavage activity involved in the biosynthesis of Citrus fruit-specific apocarotenoid pigments. J. Exp. Bot. 2013, 64, 4461–4478. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, L.; Matsuta, A.; Matsutani, K.; Yamawaki, K.; Yahata, M.; Wahyudi, A.; Motohashi, R.; Kato, M. Enzymatic Formation of -Citraurin from -Cryptoxanthin and Zeaxanthin by Carotenoid Cleavage Dioxygenase4 in the Flavedo of Citrus Fruit. Plant Physiol. 2013, 163, 682–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Zhu, K.; Sun, Q.; Zhang, W.; Wang, X.; Cao, H.; Tan, M.; Xie, Z.; Zeng, Y.; Ye, J.; et al. Natural Variation in CCD4 Promoter Underpins Species-Specific Evolution of Red Coloration in Citrus Peel. Mol. Plant 2019, 12, 1294–1307. [Google Scholar] [CrossRef] [PubMed]
- Fanciullino, A.-L.; Dhuique-Mayer, C.; Luro, F.; Casanova, J.; Morillon, R.; Ollitrault, P. Carotenoid Diversity in Cultivated Citrus Is Highly Influenced by Genetic Factors. J. Agric. Food Chem. 2006, 54, 4397–4406. [Google Scholar] [CrossRef] [PubMed]
- Tietel, Z.; Lewinsohn, E.; Fallik, E.; Porat, R. Importance of storage temperatures in maintaining flavor and quality of mandarins. Postharvest Biol. Technol. 2012, 64, 175–182. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ikoma, Y.; Kato, M.; Nakajima, N.; Hasegawa, Y. Effect of postharvest temperature and ethylene on carotenoid accumulation in the flavedo and juice sacs of satsuma mandarin (Citrus unshiu Marc.) Fruit. J. Agric. Food Chem. 2009, 57, 4724–4732. [Google Scholar] [CrossRef]
- El Airaj, H.; Gest, N.; Truffault, V.; Garchery, C.; Riqueau, G.; Gouble, B.; Page, D.; Stevens, R. Decreased monodehydroascorbate reductase activity reduces tolerance to cold storage in tomato and affects fruit antioxidant levels. Postharvest Biol. Technol. 2013, 86, 502–510. [Google Scholar] [CrossRef]
- Qian, C.; He, Z.; Zhao, Y.; Mi, H.; Chen, X.; Mao, L. Maturity-dependent chilling tolerance regulated by the antioxidative capacity in postharvest cucumber (Cucumis sativus L.) fruits. J. Sci. Food Agric. 2013, 93, 626–633. [Google Scholar] [CrossRef]
- Chongchatuporn, U.; Ketsa, S.; van Doorn, W.G. Chilling injury in mango (Mangifera indica) fruit peel: Relationship with ascorbic acid concentrations and antioxidant enzyme activities. Postharvest Biol. Technol. 2013, 86, 409–417. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreno, C.; Plaza, L.; De Ancos, B.; Cano, M.P. Quantitative bioactive compounds assessment and their relative contribution to the antioxidant capacity of commercial orange juices. J. Sci. Food Agric. 2003, 83, 430–439. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Takahashi, S.; Aizawa, K.; Mukai, K. Development of singlet oxygen absorption capacity (SOAC) assay method. 4. Measurements of the SOAC values for vegetable and fruit extracts. Biosci. Biotechnol. Biochem. 2015, 79, 280–291. [Google Scholar] [CrossRef]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant activity of Citrus fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Niki, E.; Noguchi, N.; Tsuchihashi, H.; Gotoh, N. Interaction among vitamin C, vitamin E, and beta-carotene. Am. J. Clin. Nutr. 1995, 62, 1322S–1326S. [Google Scholar] [CrossRef] [PubMed]
- Mathaba, N.; Bower, J.P.; Bertling, I. Effect of production site, storage duration, and hot water and molybdenum dips on bioactive compounds with antioxidant properties in lemon flavedo during cold storage. S. Afr. J. Plant Soil 2014, 31, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Lafuente, M.T.; Zacarias, L.; Martıínez-Téllez, M.A.; Sanchez-Ballesta, M.T.; Granell, A. Phenylalanine ammonia-lyase and ethylene in relation to chilling injury as affected by fruit age in citrus. Postharvest Biol. Technol. 2003, 29, 309–318. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Zacarias, L.; Granell, A.; Lafuente, M.T. Accumulation of PAL transcript and PAL activity as affected by heat-conditioning and low-temperature storage and its relation to chilling sensitivity in mandarin fruits. J. Agric. Food Chem. 2000, 48, 2726–2731. [Google Scholar] [CrossRef]
- Sala, J.M.; Lafuente, T. Catalase enzyme activity is related to tolerance of mandarin fruits to chilling. Postharvest Biol. Technol. 2000, 20, 81–89. [Google Scholar] [CrossRef]
- De Ancos, B.; Rodrigo, M.J.; Sánchez-Moreno, C.; Pilar Cano, M.; Zacarías, L. Effect of high-pressure processing applied as pretreatment on carotenoids, flavonoids and vitamin C in juice of the sweet oranges “Navel” and the red-fleshed “Cara Cara”. Food Res. Int. 2020, 132, 109105. [Google Scholar] [CrossRef] [PubMed]
- Zacarías-García, J.; Rey, F.; Gil, J.-V.; Rodrigo, M.J.; Zacarías, L. Antioxidant capacity in fruit of Citrus cultivars with marked differences in pulp coloration: Contribution of carotenoids and vitamin C. Food Sci. Technol. Int. 2020, 1082013220944018. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as anti-oxidants—A review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Conn, P.F.; Schalch, W.; Truscott, T.G. The singlet oxygen and carotenoid interaction. J. Photochem. Photobiol. B Biol. 1991, 11, 41–47. [Google Scholar] [CrossRef]
- Katsuura, S.; Imamura, T.; Bando, N.; Yamanishi, R. β-Carotene and β-cryptoxanthin but not lutein evoke redox and immune changes in RAW264 murine macrophages. Mol. Nutr. Food Res. 2009, 53, 1396–1405. [Google Scholar] [CrossRef] [PubMed]
- Unno, K.; Sugiura, M.; Ogawa, K.; Takabayashi, F.; Toda, M.; Sakuma, M.; Maeda, K.I.; Fujitani, K.; Miyazaki, H.; Yamamoto, H.; et al. Beta-cryptoxanthin, plentiful in Japanese mandarin orange, prevents age-related cognitive dysfunction and oxidative damage in senescence-accelerated mouse brain. Biol. Pharm. Bull. 2011, 34, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Llopis, S.; Rodrigo, M.J.; González, N.; Genovés, S.; Zacarías, L.; Ramón, D.; Martorell, P. β-Cryptoxanthin Reduces Body Fat and Increases Oxidative Stress Response in Caenorhabditis elegans Model. Nutrients 2019, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Stinco, C.M.; Escudero-Gilete, M.L.; Heredia, F.J.; Vicario, I.M.; Meléndez-Martínez, A.J. Multivariate analyses of a wide selection of orange varieties based on carotenoid contents, color and in vitro antioxidant capacity. Food Res. Int. 2016, 90, 194–204. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoids (µg g−1 FW) | FORTUNE | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 8 | ||||||||||||||||||||||
| NC | C | NC | C | NC | C | |||||||||||||||||||
| Phytoene | 6.34 | ± | 0.40 | c | 17.02 | ± | 2.53 ** | A | 12.61 | ± | 2.09 * | b | 7.64 | ± | 0.20 | B | 19.62 | ± | 1.24 ** | a | 5.09 | ± | 2.20 | B |
| Phytofluene | 1.95 | ± | 0.28 | a | 4.31 | ± | 1.03 * | A | 1.51 | ± | 0.19 | a | 1.44 | ± | 0.17 | B | 2.59 | ± | 0.87 | a | 1.00 | ± | 0.23 | B |
| β-cryptoxanthin | 3.44 | ± | 1.02 | a | 2.23 | ± | 0.30 | A | 3.56 | ± | 2.27 | a | 5.29 | ± | 2.63 | A | 1.96 | ± | 0.43 | a | 8.23 | ± | 3.19 * | A |
| β-citraurin | 41.83 | ± | 8.39 | a | 29.79 | ± | 5.48 | A | 31.68 | ± | 4.79 | a | 23.06 | ± | 1.23 | A | 39.89 | ± | 0.50 ** | a | 24.50 | ± | 3.16 | A |
| Antheraxanthin | 14.45 | ± | 1.34 | b | 14.25 | ± | 1.13 | B | 24.49 | ± | 1.64 ** | a | 7.91 | ± | 0.24 | C | 31.14 | ± | 2.61 | a | 24.74 | ± | 1.69 | B |
| Luteoxanthin | 2.74 | ± | 0.68 | b | 2.61 | ± | 0.25 | B | 2.55 | ± | 0.11 * | b | 1.87 | ± | 0.22 | B | 4.75 | ± | 0.10 | a | 5.73 | ± | 0.98 | A |
| Violaxanthin | 99.37 | ± | 2.61 | a | 86.56 | ± | 8.96 | A | 100.35 | ± | 6.97 | a | 97.50 | ± | 14.53 | A | 119.28 | ± | 4.17 | a | 120.79 | ± | 2.84 | A |
| Lutein | 0.58 | ± | 0.34 | a | 0.88 | ± | 0.40 | A | 1.19 | ± | 0.01 | a | nd | 2.07 | ± | 0.57 | a | 1.07 | ± | 0.20 | A | |||
| Other β.β-xanthophylls | 12.99 | ± | 2.58 | b | 25.62 | ± | 5.65 * | A | 23.66 | ± | 3.24 * | a | 8.96 | ± | 4.39 | A | 29.52 | ± | 0.96 ** | a | 10.05 | ± | 3.82 | A |
| Total | 185.90 | ± | 17.86 | b | 189.53 | ± | 13.67 | A | 204.03 | ± | 15.33 * | ab | 157.28 | ± | 14.24 | A | 256.32 | ± | 9.12 ** | a | 201.88 | ± | 3.75 | A |
| Carotenoids (µg g−1 FW) | NOVA | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 8 | ||||||||||||||||||||||
| NC | C | NC | C | NC | C | |||||||||||||||||||
| Phytoene | 98.12 | ± | 8.60 ** | a | 41.19 | ± | 4.01 | C | 120.23 | ± | 4.37 ** | a | 81.11 | ± | 4.83 | A | 103.43 | ± | 9.12 ** | a | 59.65 | ± | 3.30 | B |
| Phytofluene | 24.17 | ± | 2.40 ** | a | 12.71 | ± | 0.76 | C | 36.26 | ± | 0.48 ** | a | 28.34 | ± | 1.40 | A | 32.32 | ± | 4.38 * | a | 18.56 | ± | 1.18 | B |
| β-cryptoxanthin | 25.36 | ± | 1.43 * | a | 19.47 | ± | 1.43 | A | 44.54 | ± | 0.95 | b | 27.24 | ± | 14.12 | A | 24.09 | ± | 4.18 * | b | 12.54 | ± | 2.02 | A |
| β-citraurin | 37.77 | ± | 2.97 | a | 58.69 | ± | 5.68 ** | A | 23.32 | ± | 0.55 | b | 39.22 | ± | 11.51 | A | 36.26 | ± | 0.59 | a | 36.29 | ± | 0.43 | A |
| Antheraxanthin | 29.70 | ± | 2.15 ** | a | 8.22 | ± | 2.27 | B | 34.90 | ± | 4.01 * | a | 22.85 | ± | 0.86 | A | 34.93 | ± | 0.80 ** | a | 17.55 | ± | 0.46 | A |
| Luteoxanthin | 5.35 | ± | 2.64 | a | 2.44 | ± | 0.02 | A | 4.15 | ± | 0.19 ** | a | 1.83 | ± | 0.40 | A | 5.65 | ± | 0.25 ** | a | 2.41 | ± | 0.39 | A |
| Violaxanthin | 169.09 | ± | 4.00 ** | a | 133.12 | ± | 0.56 | A | 159.17 | ± | 6.42 ** | a | 116.84 | ± | 8.21 | A | 168.07 | ± | 1.13 ** | a | 134.73 | ± | 2.20 | A |
| Lutein | 7.56 | ± | 0.09 * | a | 2.59 | ± | 1.95 | A | 9.06 | ± | 1.10 ** | a | 4.11 | ± | 0.96 | A | 7.67 | ± | 1.03 * | a | 3.22 | ± | 0.03 | A |
| Other β.β-xanthophylls | 39.52 | ± | 10.13 | a | 25.00 | ± | 14.30 | A | 32.96 | ± | 6.58 ** | a | 7.38 | ± | 0.66 | A | 38.09 | ± | 5.56 * | a | 14.83 | ± | 6.96 | A |
| Total | 444.36 | ± | 1.59 ** | a | 308.85 | ± | 17.79 | A | 478.06 | ± | 16.59 ** | a | 338.44 | ± | 41.35 | A | 463.61 | ± | 15.52 ** | a | 304.30 | ± | 9.26 | A |
| Carotenoids (µg g−1 FW) | NADORCOTT | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 8 | ||||||||||||||||||||||
| NC | C | NC | C | NC | C | |||||||||||||||||||
| Phytoene | 59.90 | ± | 1.18 ** | a | 29.75 | ± | 1.07 | A | 48.06 | ± | 0.21 ** | b | 24.43 | ± | 0.84 | A | 21.06 | ± | 3.87 * | c | 12.29 | ± | 1.77 | B |
| Phytofluene | 17.30 | ± | 2.21 ** | a | 7.70 | ± | 0.30 | A | 12.88 | ± | 1.44 ** | a | 3.69 | ± | 0.63 | B | 11.28 | ± | 0.36 ** | a | 5.51 | ± | 0.49 | B |
| β-cryptoxanthin | 61.46 | ± | 8.05 | ab | 68.87 | ± | 16.53 | A | 84.87 | ± | 5.54 ** | a | 66.98 | ± | 0.83 | A | 54.15 | ± | 4.73 | b | 68.38 | ± | 2.46 * | A |
| β-citraurin | 56.75 | ± | 9.55 | a | 46.22 | ± | 6.36 | A | 32.69 | ± | 1.26 | a | 30.13 | ± | 5.38 | A | 48.52 | ± | 5.48 ** | a | 31.15 | ± | 1.02 | A |
| Antheraxanthin | 42.74 | ± | 2.15 ** | b | 26.37 | ± | 2.22 | B | 58.21 | ± | 1.70 ** | a | 42.56 | ± | 0.64 | A | 50.96 | ± | 4.52 ** | ab | 33.08 | ± | 1.59 | B |
| Luteoxanthin | 7.54 | ± | 3.36 | a | 2.85 | ± | 1.00 | A | 3.87 | ± | 2.97 | a | 5.41 | ± | 0.63 | A | 5.06 | ± | 1.37 | a | 2.05 | ± | 0.15 | A |
| Violaxanthin | 264.06 | ± | 16.51 ** | a | 209.95 | ± | 2.14 | A | 242.69 | ± | 3.51 ** | ab | 225.78 | ± | 1.06 | A | 218.18 | ± | 3.71 | b | 202.19 | ± | 19.83 | A |
| Lutein | 9.82 | ± | 1.82 | a | 8.30 | ± | 0.65 | B | 17.41 | ± | 2.54 | a | 13.33 | ± | 0.03 | A | 11.23 | ± | 2.28 | a | 12.19 | ± | 1.19 | A |
| Other β.β-xanthophylls | 51.10 | ± | 5.83 ** | a | 18.72 | ± | 2.21 | A | 54.06 | ± | 9.67 ** | a | 13.65 | ± | 0.94 | AB | 34.14 | ± | 5.62 * | a | 12.58 | ± | 0.08 | B |
| Total | 582.20 | ± | 22.85 ** | a | 424.99 | ± | 16.80 | AB | 565.05 | ± | 9.10 ** | a | 430.16 | ± | 2.50 | A | 479.82 | ± | 14.39 ** | b | 381.69 | ± | 25.44 | B |
| CI | TCC | β-Cryptoxanthin | β-Citraurin | Violaxanthin | Vitamin C | SOAC | DPPH• | FRAP | |
|---|---|---|---|---|---|---|---|---|---|
| CI | 1 | −0.81 * | −0.82 * | −0.74 | −0.85 * | 0.82 * | −0.88 * | 0.89 * | 0.81 * |
| TCC | −0.81 * | 1 | 0.85 * | 0.52 | 0.97 * | −0.62 | 0.96 * | −0.52 | −0.41 |
| β-cryptoxanthin | −0.82 * | 0.85 * | 1 | 0.51 | 0.93 * | −0.78 | 0.92 * | −0.73 | −0.71 |
| β-citraurin | −0.74 | 0.52 | 0.51 | 1 | 0.57 | −0.51 | 0.55 | −0.75 | −0.71 |
| Violaxanthin | −0.85 * | 0.97 * | 0.93 * | 0.57 | 1 | −0.64 | 0.99 * | −0.62 | −0.57 |
| Vitamin C | 0.82 * | −0.62 | −0.78 | −0.51 | −0.64 | 1 | −0.66 | 0.87 * | 0.75 |
| SOAC | −0.88 * | 0.96 * | 0.92 * | 0.55 | 0.99 * | −0.66 | 1 | −0.65 | −0.60 |
| DPPH• | 0.89 * | −0.52 | −0.73 | −0.75 | −0.62 | 0.87 * | −0.65 | 1 | 0.96 * |
| FRAP | 0.81 * | −0.41 | −0.71 | −0.71 | −0.57 | 0.75 | −0.60 | 0.96 * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rey, F.; Zacarías, L.; Rodrigo, M.J. Carotenoids, Vitamin C, and Antioxidant Capacity in the Peel of Mandarin Fruit in Relation to the Susceptibility to Chilling Injury during Postharvest Cold Storage. Antioxidants 2020, 9, 1296. https://doi.org/10.3390/antiox9121296
Rey F, Zacarías L, Rodrigo MJ. Carotenoids, Vitamin C, and Antioxidant Capacity in the Peel of Mandarin Fruit in Relation to the Susceptibility to Chilling Injury during Postharvest Cold Storage. Antioxidants. 2020; 9(12):1296. https://doi.org/10.3390/antiox9121296
Chicago/Turabian StyleRey, Florencia, Lorenzo Zacarías, and María J. Rodrigo. 2020. "Carotenoids, Vitamin C, and Antioxidant Capacity in the Peel of Mandarin Fruit in Relation to the Susceptibility to Chilling Injury during Postharvest Cold Storage" Antioxidants 9, no. 12: 1296. https://doi.org/10.3390/antiox9121296