The Protective Role of Hydrogen Sulfide Against Obesity-Associated Cellular Stress in Blood Glucose Regulation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. PA/FAF-BSA Conjugation Preparation
2.3. Intracellular ROS Detection
2.4. Cytotoxicity Assessment of Palmitate
2.5. Animals
2.6. In Vivo Experimental Design
2.7. Oral Glucose Tolerance Test and GLP-1 Determination
2.8. In Vivo Oxidative Stress Analysis
2.9. Statistical Analysis
3. Results
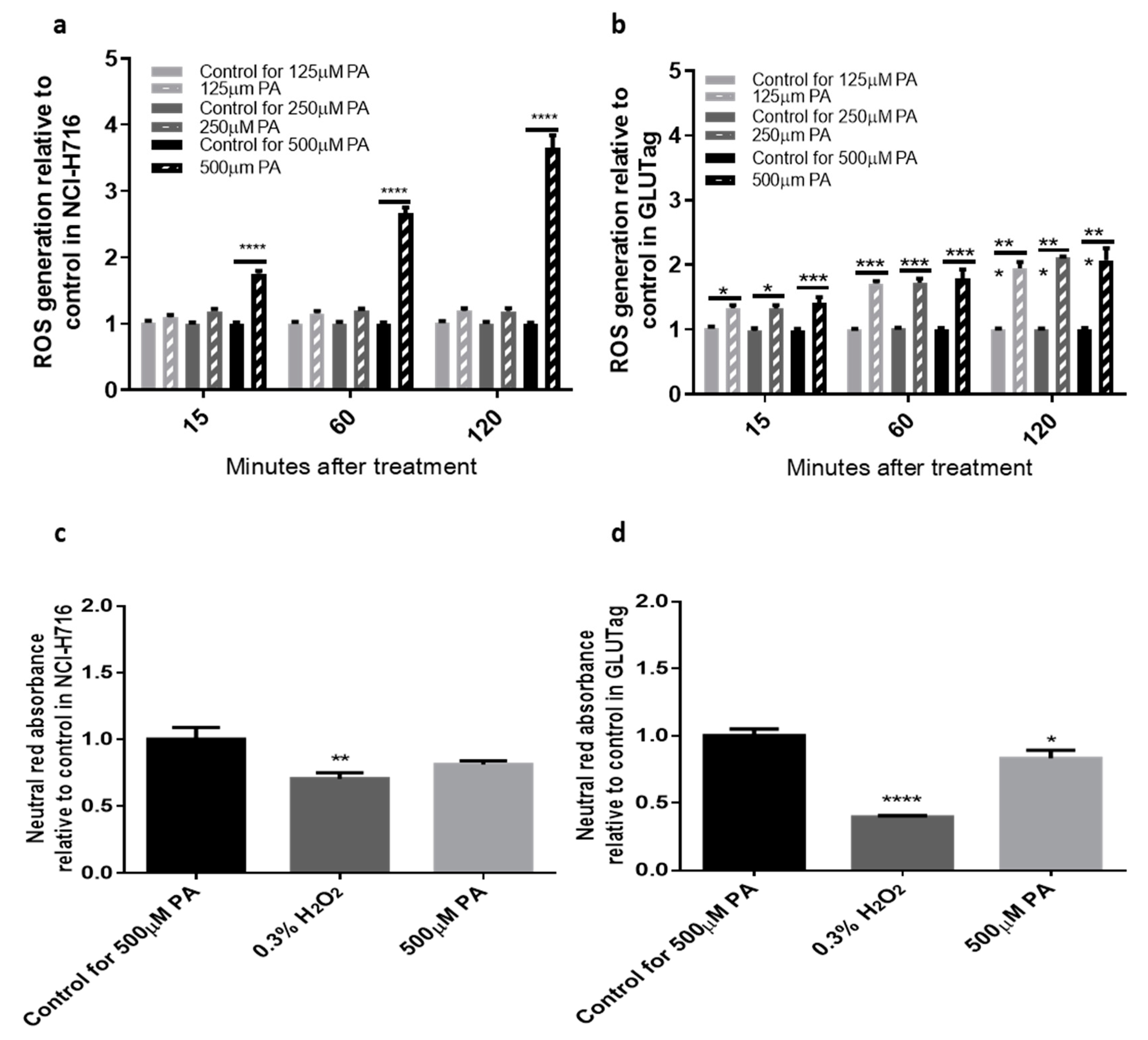
3.1. Palmitate Stimulates ROS Production in Both NCI-H716 and GLUTag Cells
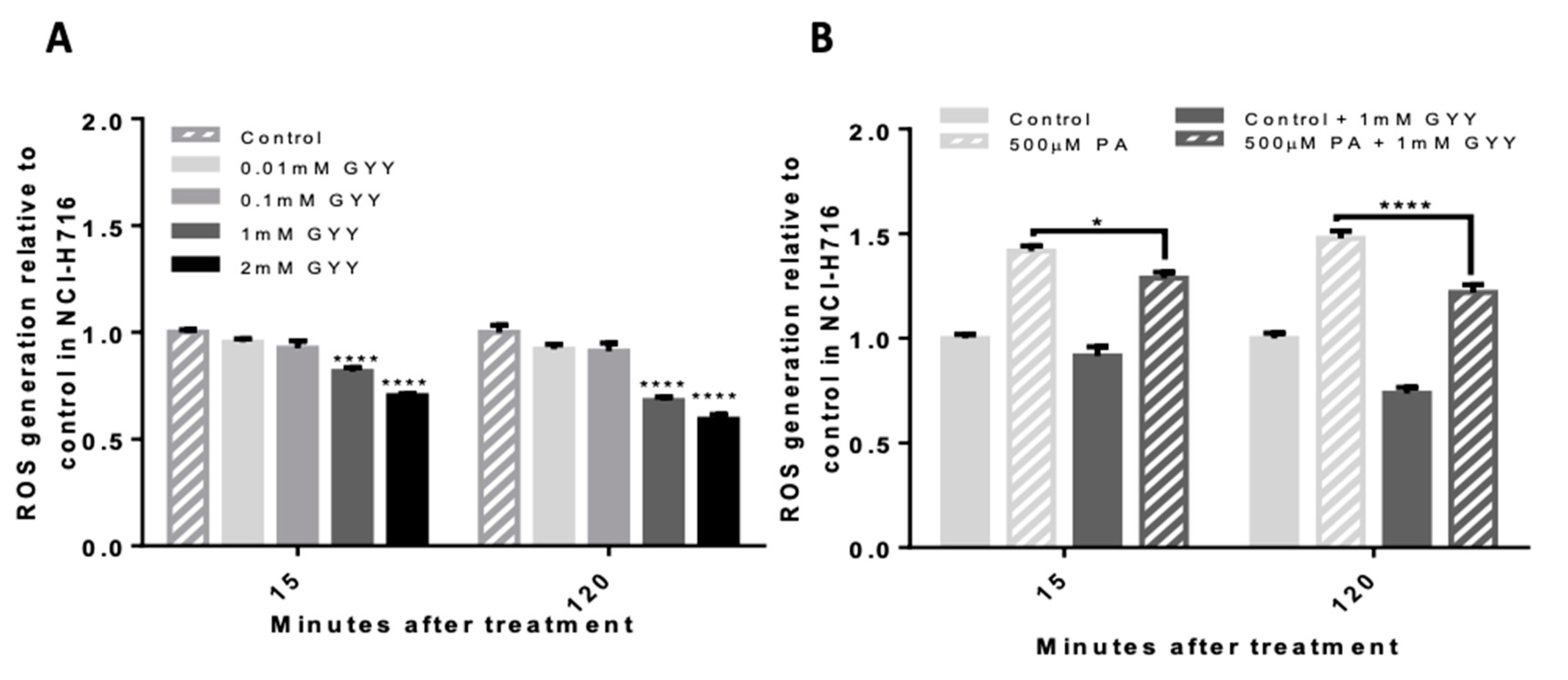
3.2. GYY4137, a Slow-Releasing H2S Donor, Reduces Basal and Palmitate-Induced ROS in NCI-H716 Cells
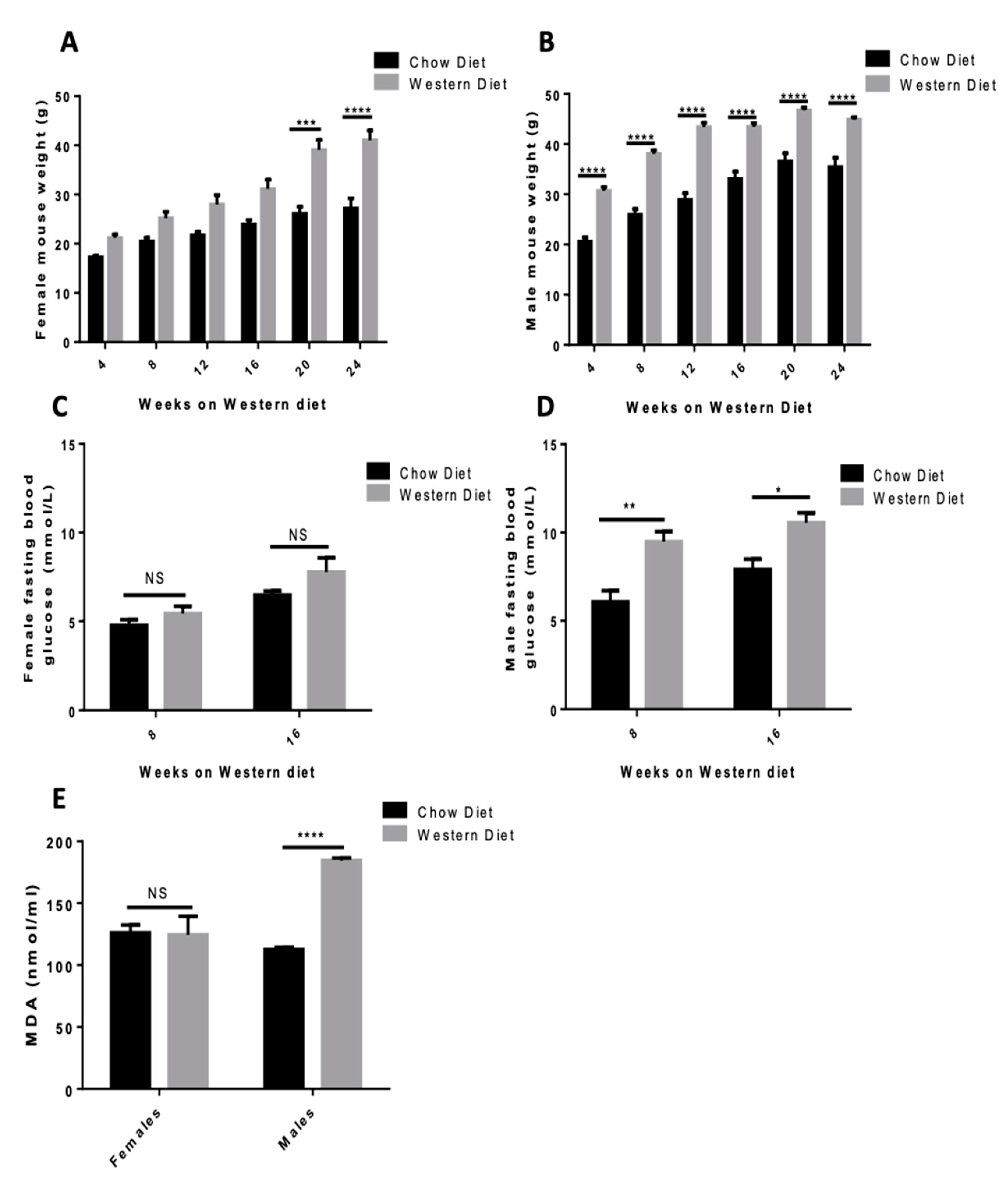
3.3. Western Diet Elevates Body Weight in Both Sexes and Elevates Fasting Blood Glucose and Lipid Peroxidation in Male Mice
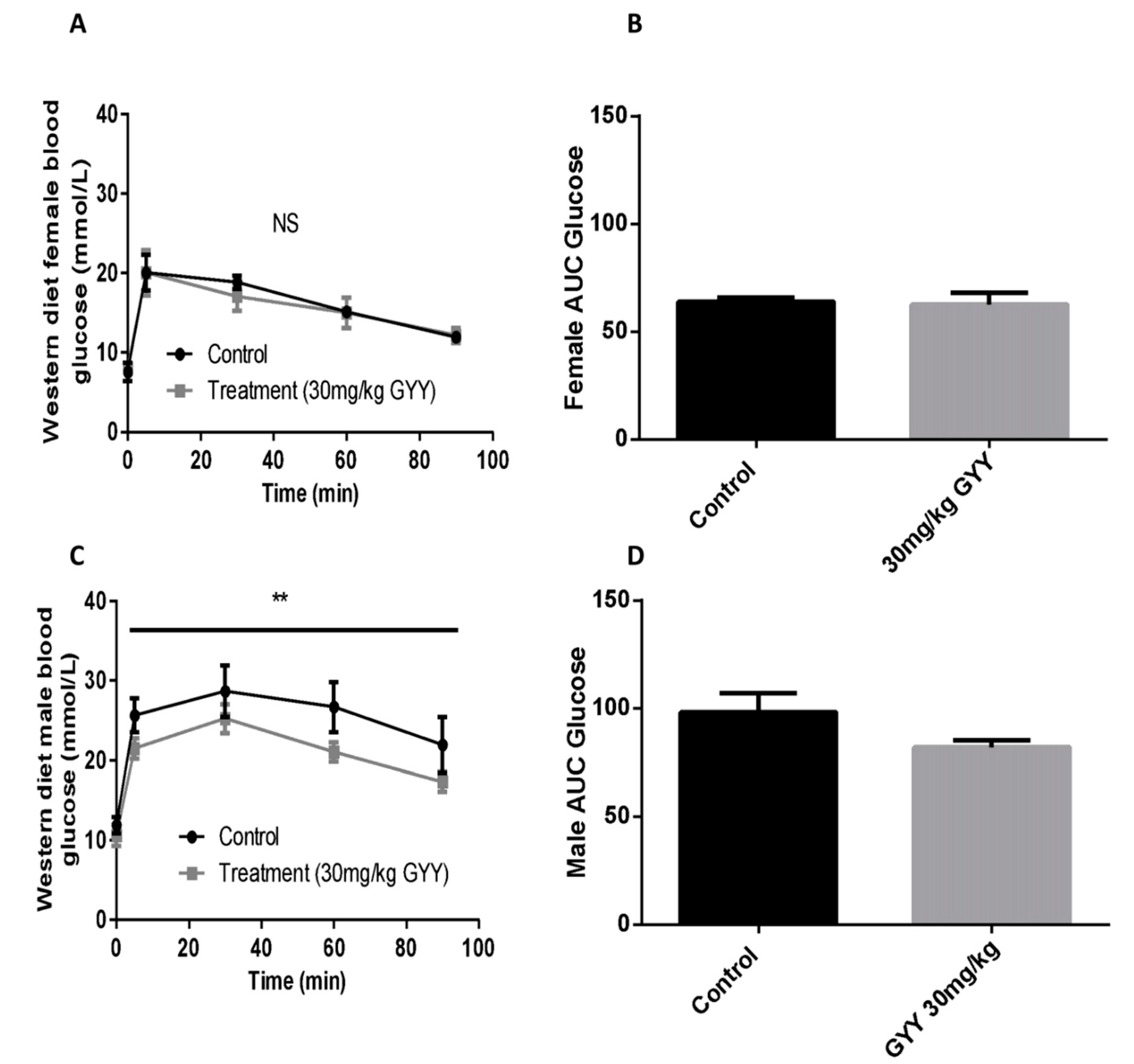
3.4. The H2S Donor, GYY4137, Improves Oral Glucose Tolerance in Western Diet-Fed Male Mice
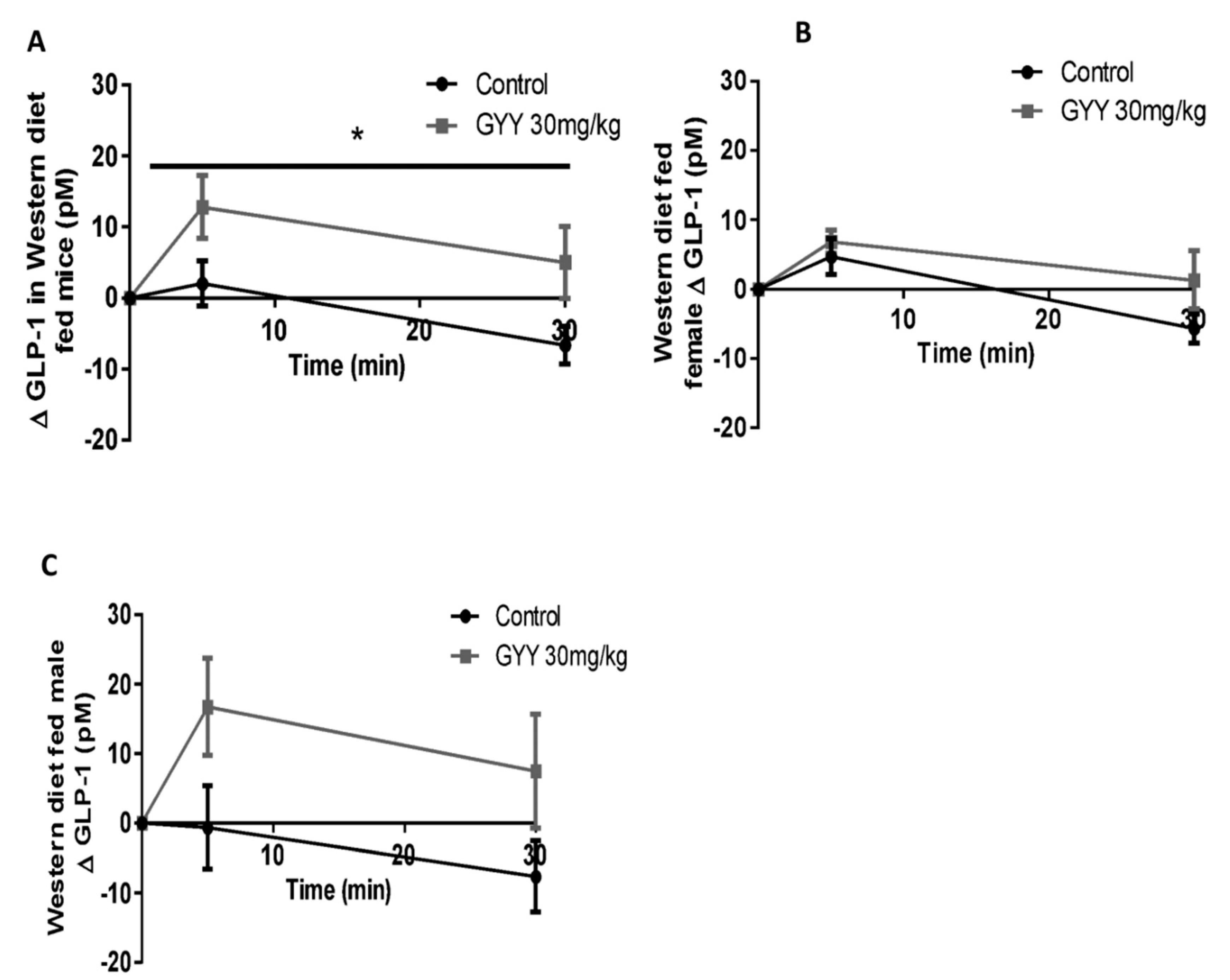
3.5. The H2S Donor, GYY4137, Enhances Glucose Stimulated GLP-1 Release in Western Diet-Fed Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebovitz, H.E.; Banerji, M.A. Point: Visceral Adiposity Is Causally Related to Insulin Resistance. Diabetes Care 2005, 28, 2322–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paccosi, S.; Cresci, B.; Pala, L.; Rotella, C.M.; Parenti, A. Obesity Therapy: How and Why? Curr. Med. Chem. 2020, 27, 174–186. [Google Scholar] [CrossRef]
- Brunton, S. GLP-1 Receptor Agonists vs. DPP-4 Inhibitors for Type 2 Diabetes: Is One Approach More Successful or Preferable than the Other? Int. J. Clin. Pract. 2014, 68, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallwitz, B. Clinical Use of DPP-4 Inhibitors. Front. Endocrinol. (Lausanne) 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Heppner, K.M.; Perez-Tilve, D. GLP-1 Based Therapeutics: Simultaneously Combating T2DM and Obesity. Front. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hira, T.; Pinyo, J.; Hara, H. What Is GLP-1 Really Doing in Obesity? Trends Endocrinol. Metab. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Drucker, D.J.; Nauck, M.A. The Incretin System: Glucagon-like Peptide-1 Receptor Agonists and Dipeptidyl Peptidase-4 Inhibitors in Type 2 Diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Madsbad, S. The Role of Glucagon-like Peptide-1 Impairment in Obesity and Potential Therapeutic Implications. Diabetes Obes. Metab. 2014, 16, 9–21. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and Its Impact on Metabolic Syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes Mellitus and Oxidative Stress—A Concise Review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, P.A.; Rutter, G.A. The Role of Oxidative Stress and Hypoxia in Pancreatic Beta-Cell Dysfunction in Diabetes Mellitus. Antioxid. Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, L.D.; Xu, S.; Choi, S.-K.; Ha, C.-M.; Thoudam, T.; Cha, S.-K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.-K.; Park, K.-S. Oxidative Stress and Calcium Dysregulation by Palmitate in Type 2 Diabetes. Exp. Mol. Med. 2017, 49, e291. [Google Scholar] [CrossRef] [PubMed]
- Shefa, U.; Kim, M.-S.; Jeong, N.Y.; Jung, J. Antioxidant and Cell-Signaling Functions of Hydrogen Sulfide in the Central Nervous System. Oxidative Med. Cell. Longev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chiku, T.; Padovani, D.; Zhu, W.; Singh, S.; Vitvitsky, V.; Banerjee, R. H2S Biogenesis by Human Cystathionine γ-Lyase Leads to the Novel Sulfur Metabolites Lanthionine and Homolanthionine and Is Responsive to the Grade of Hyperhomocysteinemia. J. Biol. Chem. 2009, 284, 11601–11612. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B.; Lin, H.C. Hydrogen Sulfide in Physiology and Diseases of the Digestive Tract. Microorganisms 2015, 3, 866–889. [Google Scholar] [CrossRef] [Green Version]
- Pichette, J.; Fynn-Sackey, N.; Gagnon, J. Hydrogen Sulfide and Sulfate Prebiotic Stimulates the Secretion of GLP-1 and Improves Glycemia in Male Mice. Endocrinology 2017, 158, 3416–3425. [Google Scholar] [CrossRef]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A Complex Protein Mixture Required for Optimal Growth of Cell Culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef]
- Cousin, S.P.; Hügl, S.R.; Wrede, C.E.; Kajio, H.; Myers, M.G., Jr.; Rhodes, C.J. Free fatty acid induced inhibition of glucose and insulin-like growth factor I-induced deoxyribonucleic acid synthesis in the pancreatic β-cell line INS-1. Endocrinology 2001, 142, 229–240. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral Red Uptake Assay for the Estimation of Cell Viability/Cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Arner, P.; Rydén, M. Fatty Acids, Obesity and Insulin Resistance. Obes. Facts 2015, 8, 147–155. [Google Scholar] [CrossRef]
- Vasu, S.; Moffett, R.C.; McClenaghan, N.H.; Flatt, P.R. Differential Molecular and Cellular Responses of GLP-1 Secreting L-Cells and Pancreatic Alpha Cells to Glucotoxicity and Lipotoxicity. Exp. Cell Res. 2015, 336, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Filippello, A.; Urbano, F.; Di Mauro, S.; Scamporrino, A.; Di Pino, A.; Scicali, R.; Rabuazzo, A.M.; Purrello, F.; Piro, S. Chronic Exposure to Palmitate Impairs Insulin Signaling in an Intestinal L-Cell Line: A Possible Shift from GLP-1 to Glucagon Production. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Kuhre, R.E.; Wewer Albrechtsen, N.J.; Deacon, C.F.; Balk-Møller, E.; Rehfeld, J.F.; Reimann, F.; Gribble, F.M.; Holst, J.J. Peptide Production and Secretion in GLUTag, NCI-H716, and STC-1 Cells: A Comparison to Native L-Cells. J. Mol. Endocrinol. 2016, 56, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Al-Magableh, M.R.; Kemp-Harper, B.K.; Hart, J.L. Hydrogen Sulfide Treatment Reduces Blood Pressure and Oxidative Stress in Angiotensin II-Induced Hypertensive Mice. Hypertens. Res. 2015, 38, 13–20. [Google Scholar] [CrossRef]
- Cheung, S.H.; Lau, J.Y.W. Hydrogen Sulfide Mediates Athero-Protection against Oxidative Stress via S-Sulfhydration. PLoS ONE 2018, 13, e0194176. [Google Scholar] [CrossRef] [Green Version]
- Bian, J.-S.; Olson, K.R.; Zhu, Y.-C. Hydrogen Sulfide: Biogenesis, Physiology, and Pathology. Oxidative Med. Cell. Longev. 2016, 2016, 6549625. [Google Scholar] [CrossRef]
- Sun, W.-H.; Liu, F.; Chen, Y.; Zhu, Y.-C. Hydrogen Sulfide Decreases the Levels of ROS by Inhibiting Mitochondrial Complex IV and Increasing SOD Activities in Cardiomyocytes under Ischemia/Reperfusion. Biochem. Biophys. Res. Commun. 2012, 421, 164–169. [Google Scholar] [CrossRef]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S Signals through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [Green Version]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Baena, M.; Sangüesa, G.; Hutter, N.; Beltrán, J.M.; Sánchez, R.M.; Roglans, N.; Alegret, M.; Laguna, J.C. Liquid Fructose in Western-Diet-Fed Mice Impairs Liver Insulin Signaling and Causes Cholesterol and Triglyceride Loading without Changing Calorie Intake and Body Weight. J. Nutr. Biochem. 2017, 40, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Yonny, M.E.; García, E.M.; López, A.; Arroquy, J.I.; Nazareno, M.A. Measurement of Malondialdehyde as Oxidative Stress Biomarker in Goat Plasma by HPLC-DAD. Microchem. J. 2016, 129, 281–285. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The Role of Estrogens in Control of Energy Balance and Glucose Homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F. Sex Differences in Metabolic Homeostasis, Diabetes, and Obesity. Biol. Sex Differ. 2015, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Lee, Z.W.; Zhou, J.; Chen, C.-S.; Zhao, Y.; Tan, C.-H.; Li, L.; Moore, P.K.; Deng, L.-W. The Slow-Releasing Hydrogen Sulfide Donor, GYY4137, Exhibits Novel Anti-Cancer Effects In Vitro and In Vivo. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Drucker, N.A.; Jensen, A.R.; Te Winkel, J.P.; Markel, T.A. Hydrogen Sulfide Donor GYY4137 Acts through Endothelial Nitric Oxide to Protect Intestine in Murine Models of Necrotizing Enterocolitis and Intestinal Ischemia. J. Surg Res. 2019, 234, 294–302. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Surmiak, M.; Hubalewska-Mazgaj, M.; Kwiecien, S.; Wallace, J.L.; Brzozowski, T. The Effect of Hydrogen Sulfide-Releasing Naproxen (ATB-346) versus Naproxen on Formation of Stress-Induced Gastric Lesions, the Regulation of Systemic Inflammation, Hypoxia and Alterations in Gastric Microcirculation. J. Physiol. Pharmacol. 2017, 68, 749–756. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezouari, A.; Nangia, R.; Gagnon, J. The Protective Role of Hydrogen Sulfide Against Obesity-Associated Cellular Stress in Blood Glucose Regulation. Antioxidants 2020, 9, 1038. https://doi.org/10.3390/antiox9111038
Mezouari A, Nangia R, Gagnon J. The Protective Role of Hydrogen Sulfide Against Obesity-Associated Cellular Stress in Blood Glucose Regulation. Antioxidants. 2020; 9(11):1038. https://doi.org/10.3390/antiox9111038
Chicago/Turabian StyleMezouari, Ania, Radhika Nangia, and Jeffrey Gagnon. 2020. "The Protective Role of Hydrogen Sulfide Against Obesity-Associated Cellular Stress in Blood Glucose Regulation" Antioxidants 9, no. 11: 1038. https://doi.org/10.3390/antiox9111038