
ΔMST and the Regulation of Cardiac CSE and OTR Expression in Trauma and Hemorrhage

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Polytrauma Model
2.2. Mitochondrial Respiration
2.3. Immunohistochemistry
2.4. Statistical Analysis
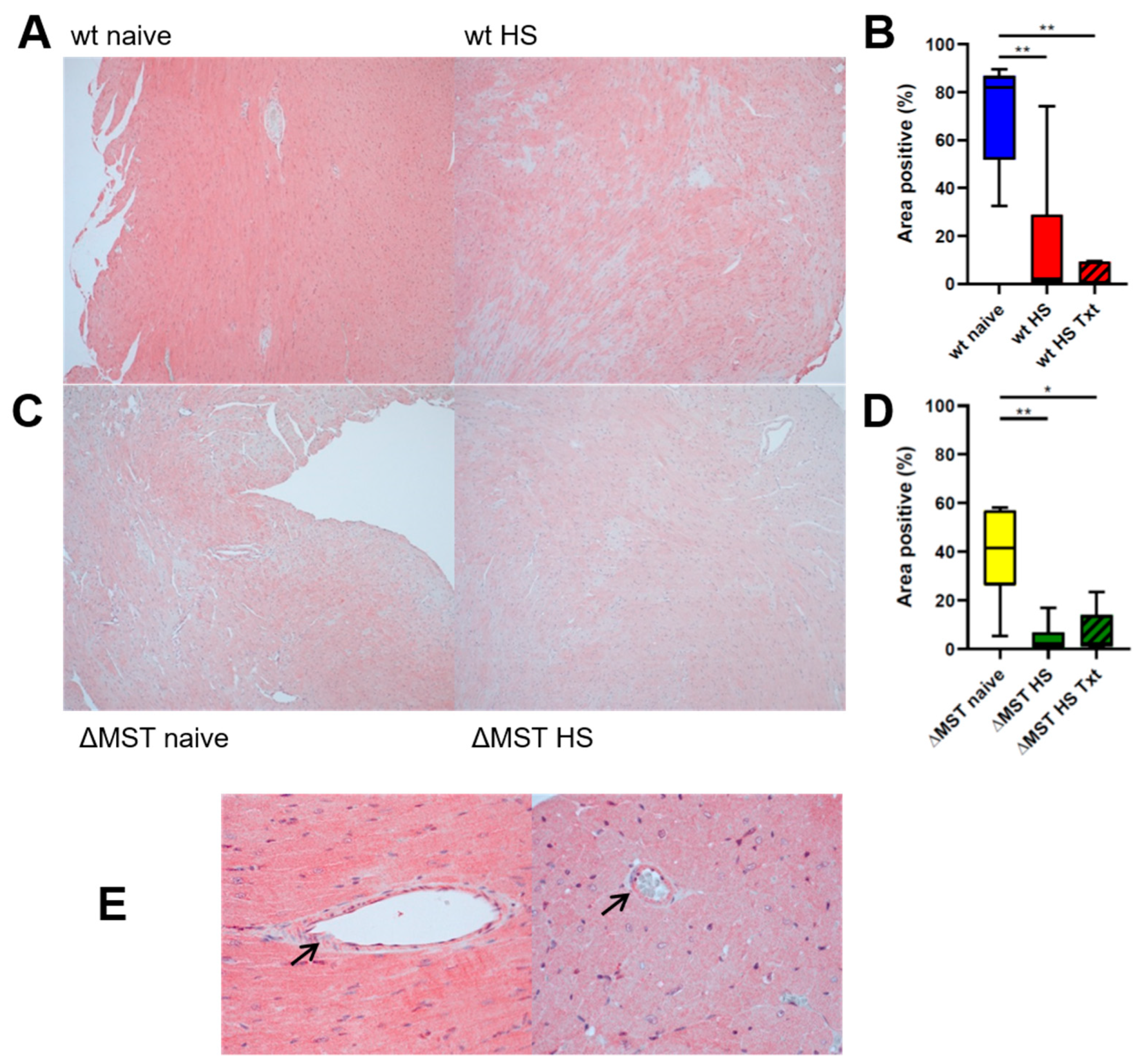
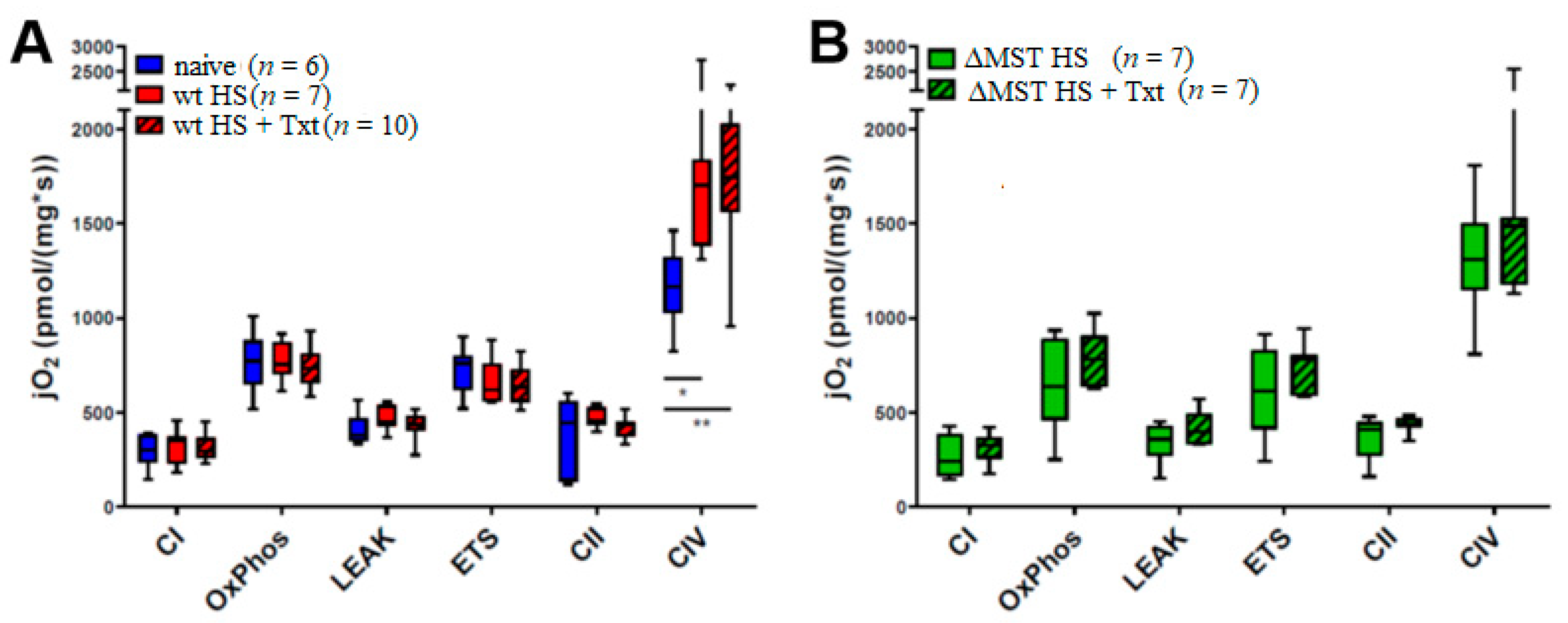
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of Variance |
| CBS | Cystathionine-β-synthase |
| CI | Mitochondrial oxygen consumption after addition of substrates for the stimulation of complex I |
| CII | Mitochondrial oxygen consumption depending on the activity of complex II after inhibition of complex I |
| CIV | Oxygen consumption after stimulation of complex IV |
| CSE | Cystathionine-γ-lyase |
| ΔMST | MST mutation |
| ELS | Early-life stress |
| ETS | Maximum respiratory capacity in the uncoupled state |
| FCCP | Carbonyl cyanide-4-(trifluoromethoxy)phenylhydrazone |
| HS | Hemorrhagic shock |
| H2S | Hydrogen sulfide |
| IHC | Immunohistochemistry |
| jO2 | Oxygen flux |
| ko | Genetic deletion/knock-out |
| LEAK | Mitochondrial oxygen consumption after inhibition of ATP synthase in the coupled state |
| LPS | Lipopolysaccharide |
| LTSS | Long-term separation stress |
| MAP | Mean arterial pressure |
| MIR | Myocardial ischemia/reperfusion |
| MST | 3-mercaptopyruvate sulfurtransferase |
| NO | Nitric oxide |
| OT | Oxytocin |
| OTR | Oxytocin receptor |
| OxPhos | Maximum respiratory capacity in the coupled state |
| STSS | Short-term separation stress |
| TMPD | N,N,N’,N’-Tetramethyl-p-phenylenediamine dihydrochloride |
| Txt | Blunt chest trauma |
| WB | Western blot |
| wt | Wild type |
References
- Denoix, N.; McCook, O.; Ecker, S.; Wang, R.; Waller, C.; Radermacher, P.; Merz, T. The Interaction of the Endogenous Hydrogen Sulfide and Oxytocin Systems in Fluid Regulation and the Cardiovascular System. Antioxidants 2020, 9, 748. [Google Scholar] [CrossRef] [PubMed]
- Peleli, M.; Bibli, S.I.; Li, Z.; Chatzianastasiou, A.; Varela, A.; Katsouda, A.; Zukunft, S.; Bucci, M.; Vellecco, V.; Davos, C.H.; et al. Cardiovascular phenotype of mice lacking 3-mercaptopyruvate sulfurtransferase. Biochem. Pharmacol. 2020, 176, 113833. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, N.; Nagano, M.; Ito, T.; Shimamura, K.; Akimoto, T.; Suzuki, H. Antioxidant enzyme, 3-mercaptopyruvate sulfurtransferase-knockout mice exhibit increased anxiety-like behaviors: A model for human mercaptolactate-cysteine disulfiduria. Sci. Rep. 2013, 3, 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröger, M.; Wepler, M.; Wachter, U.; Merz, T.; McCook, O.; Kress, S.; Lukaschewski, B.; Hafner, S.; Huber-Lang, M.; Calzia, E.; et al. The Effects of Genetic 3-Mercaptopyruvate Sulfurtransferase Deficiency in Murine Traumatic-Hemorrhagic Shock. Shock 2019, 51, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, X.L.; Liu, H.R.; Rose, P.; Zhu, Y.Z. Protective effects of cysteine analogues on acute myocardial ischemia: Novel modulators of endogenous H2S production. Antioxid. Redox. Signal. 2010, 12, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Kondo, K.; Bhushan, S.; King, A.L.; Prabhu, S.D.; Hamid, T.; Koenig, S.; Murohara, T.; Predmore, B.L.; Gojon, G., Sr.; Gojon, G., Jr.; et al. H2S protects against pressure overload-induced heart failure via upregulation of endothelial nitric oxide synthase. Circulation 2013, 127, 1116–1127. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, F.; Yin, J.; Wu, S.; Zhou, X. Protective mechanisms of hydrogen sulfide in myocardial ischemia. J. Cell. Physiol. 2020, 235, 9059–9070. [Google Scholar] [CrossRef]
- Ellmers, L.J.; Templeton, E.M.; Pilbrow, A.P.; Frampton, C.; Ishii, I.; Moore, P.K.; Bhatia, M.; Richards, A.M.; Cameron, V.A. Hydrogen Sulfide Treatment Improves Post-Infarct Remodeling and Long-Term Cardiac Function in CSE Knockout and Wild-Type Mice. Int. J. Mol. Sci. 2020, 21, 4284. [Google Scholar] [CrossRef]
- Szabo, C.; Ransy, C.; Módis, K.; Andriamihaja, M.; Murghes, B.; Coletta, C.; Olah, G.; Yanagi, K.; Bouillaud, F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part I. Biochemical and physiological mechanisms. Br. J. Pharmacol. 2014, 171, 2099–2122. [Google Scholar] [CrossRef] [Green Version]
- Denoix, N.; McCook, O.; Scheuerle, A.; Kapapa, T.; Hoffmann, A.; Gündel, H.; Waller, C.; Radermacher, P.; Merz, T. Brain histology and immunohistochemistry after resuscitation from hemorrhagic shock in swine with pre-existing atherosclerosis: Effects of sodium thiosulfate (Na2S2O3). Front. Pharmacol. 2021. under review. [Google Scholar]
- Amini-Khoei, H.; Amiri, S.; Mohammadi-Asl, A.; Alijanpour, S.; Poursaman, S.; Haj-Mirzaian, A.; Rastegar, M.; Mesdaghinia, A.; Banafshe, H.R.; Sadeghi, E.; et al. Experiencing neonatal maternal separation increased pain sensitivity in adult male mice: Involvement of oxytocinergic system. Neuropeptides 2017, 61, 77–85. [Google Scholar] [CrossRef]
- Albus, C.; Herrmann-Lingen, C.; Jensen, K.; Hackbusch, M.; Münch, N.; Kuncewicz, C.; Grilli, M.; Schwaab, B.; Rauch, B. For the German Society of Cardiovascular Prevention & Rehabilitation (DGPR). Additional effects of psychological interventions on subjective and objective outcomes compared with exercise-based cardiac rehabilitation alone in patients with cardiovascular disease: A systematic review and meta-analysis. Eur. J. Prev. Cardiol. 2019, 26, 1035–1049. [Google Scholar]
- D’Andrea, W.; Sharma, R.; Zelechoski, A.D.; Spinazzola, J. Physical health problems after single trauma exposure: When stress takes root in the body. J. Am. Psychiatr. Nurses Assoc. 2011, 17, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Wigger, D.C.; Gröger, N.; Lesse, A.; Krause, S.; Merz, T.; Gündel, H.; Braun, K.; McCook, O.; Radermacher, P.; Bock, J.; et al. Maternal Separation Induces Long-Term Alterations in the Cardiac Oxytocin Receptor and Cystathionine γ-Lyase Expression in Mice. Oxid. Med. Cell Longev. 2020, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merz, T.; Lukaschewski, B.; Wigger, D.; Rupprecht, A.; Wepler, M.; Gröger, M.; Hartmann, C.; Whiteman, M.; Szabo, C.; Wang, R.; et al. Interaction of the hydrogen sulfide system with the oxytocin system in the injured mouse heart. Intensive Care Med. Exp. 2018, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenema, A.H.; Bredewold, R.; Neumann, I.D. Opposite effects of maternal separation on intermale and maternal aggression in C57BL/6 mice: Link to hypothalamic vasopressin and oxytocin immunoreactivity. Psychoneuroendocrinology 2007, 32, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-J.; Chen, Y.-C.; Chien, T.-H.; Chang, H.-Y.; Chen, Y.-H.; Chien, C.-Y.; Huang, T.-S. Impact of comorbidities on the prognoses of trauma patients: Analysis of a hospital-based trauma registry database. PLoS ONE 2018, 13, e0194749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraris, V.A.; Ferraris, S.P.; Saha, S.P. The relationship between mortality and preexisting cardiac disease in 5971 trauma patients. J. Trauma Acute Care Surg. 2010, 69, 645–652. [Google Scholar]
- Auxéméry, Y. Post-traumatic psychiatric disorders: PTSD is not the only diagnosis. Presse. Med. 2018, 47, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Gero, D.; Ahmad, A.; Brunyanszki, A.; Olah, G.; Szczesny, B.; Szabo, C. 3-Mercaptopyruvate sulfurtransferase deficient mice show accelerated glucose uptake and a dysregulated metabolic profile. Nitric Oxide 2015, 47, S35–S36. [Google Scholar] [CrossRef]
- Ahmad, A.; Gerö, D.; Ola, G.; Szabo, C. Effect of endotoxemia in mice genetically deficient in cystathionine-γ-lyase, cystathionine-β-synthase or 3-mercaptopyruvate sulfurtransferase. Int. J. Mol. Med. 2016, 38, 1683–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wepler, M.; Merz, T.; Wachter, U.; Vogt, J.; Calzia, E.; Scheuerle, A.; Möller, P.; Gröger, M.; Kress, S.; Fink, M.; et al. The Mitochondria-Targeted H2S-Donor AP39 in a Murine Model of Combined Hemorrhagic Shock and Blunt Chest Trauma. Shock 2019, 52, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Gröger, M.; Noirhomme, J.P.; Scheuerle, A.; Möller, P.; Wachter, U.; Huber-Lang, M.; Nussbaum, B.; Jung, B.; Merz, T.; et al. In-Depth Characterization of the Effects of Cigarette Smoke Exposure on the Acute Trauma Response and Hemorrhage in Mice. Shock 2019, 51, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Merz, T.; Vogt, J.A.; Wachter, U.; Calzia, E.; Szabo, C.; Wang, R.; Radermacher, P.; McCook, O. Impact of hyperglycemia on cystathionine-γ-lyase expression during resuscitated murine septic shock. Intensive Care Med. Exp. 2017, 5, 1–14. [Google Scholar] [CrossRef]
- Merz, T.; Wepler, M.; Nußbaum, B.; Vogt, J.; Calzia, E.; Wang, R.; Szabo, C.; Radermacher, P.; McCook, O. Cystathionine-γ-lyase expression is associated with mitochondrial respiration during sepsis-induced acute kidney injury in swine with atherosclerosis. Intensive Care Med. Exp. 2018, 6, 43. [Google Scholar] [CrossRef]
- Nagahara, N.; Ito, T.; Kitamura, H. Tissue and subcellular distribution of mercaptopyruvate sulfurtransferasein the rat: Confocal laser fluorescence and immunoelectron microscopic studies combined with biochemicalanalysis. Histochem. Cell Biol. 1998, 110, 243–250. [Google Scholar] [CrossRef]
- Krause, S.; Boeck, C.; Gumpp, A.M.; Rottler, E.; Schury, K.; Karabatsiakis, A.; Buchheim, A.; Gündel, H.; Kolassa, I.T.; Waller, C. Child Maltreatment Is Associated with a Reduction of the Oxytocin Receptor in Peripheral Blood Mononuclear Cells. Front. Psychol. 2018, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Druzhyna, N.; Szabo, C. Effect of 3-mercaptopyruvate sulfurtransferase deficiency on the development of multiorgan failure, inflammation and wound healing in mice subjected to burn injury. J. Burn. Care Res. 2019, 40, 148–156. [Google Scholar] [CrossRef]
- Li, Z.; Xia, H.; Sharp, T.; Hidalgo, H.; Nagahara, N.; Elrod, J.W.; Lefer, D.J. Deficiency of 3-Mercaptopyruvate Sulfurtransferase Results in Impaired Mitochondrial Function and Increased Heart Failure Severity. Circulation 2019, 140, A15987. [Google Scholar]
- Bordt, E.A.; Smith, C.J.; Demarest, T.G.; Bilbo, S.D.; Kingsbury, M.A. Mitochondria, Oxytocin, and Vasopressin: Unfolding the Inflammatory Protein Response. Neurotox. Res. 2019, 36, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Pappas, C.; Tajiri, N.; Borlongan, C.V. Oxytocin modulates GABAAR subunits to confer neuroprotection in stroke in vitro. Sci. Rep. 2016, 6, 35659. [Google Scholar] [CrossRef] [Green Version]
- Nussbaum, B.L.; McCook, O.; Hartmann, C.; Matallo, J.; Wepler, M.; Antonucci, E.; Kalbitz, M.; Huber-Lang, M.; Georgieff, M.; Calzia, E.; et al. Left ventricular function during porcine resuscitated septic shock with pre-existing atherosclerosis. Intensive Care Med. Exp. 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Gutkowska, J.; Jankowski, M.; Antunes-Rodrigues, J. The role of oxytocin in cardiovascular regulation. Braz. J. Med. Biol. Res. 2014, 47, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merz, T.; Stenzel, T.; Nussbaum, B.; Wepler, M.; Szabo, C.; Wang, R.; Radermacher, P.; McCook, O. Cardiovascular disease and resuscitated septic shock lead to the downregulation of the H2S producing enzyme cystathionine-γ-lyase in the porcine coronary artery. Intensive Care Med. Exp. 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghimian, M.; Faghihi, M.; Karimian, S.M.; Imani, A.; Houshmand, F.; Azizi, Y. The role of central oxytocin in stress-induced cardioprotection in ischemic-reperfused heart model. J. Cardiol. 2013, 61, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrod, R.J.; Gozal, Y.; Navot, N.; Zuo, Z. Activation of adenosine triphosphate-regulated potassium channels during reperfusion restores isoflurane postconditioning-induced cardiac protection in acutely hyperglycemic rabbits. Anesthesiology 2015, 122, 1299–1311. [Google Scholar]
- Jin, S.; Teng, X.; Xiao, L.; Xue, H.; Guo, Q.; Duan, X.; Chen, Y.; Wu, Y. Hydrogen sulfide ameliorated L-NAME-induced hypertensive heart disease by the Akt/eNOS/NO pathway. Exp. Biol. Med. 2017, 242, 1831–1841. [Google Scholar] [CrossRef]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [Green Version]
- Merz, T.; Denoix, N.; Wigger, D.; Waller, C.; Wepler, M.; Vettorazzi, S.; Tuckermann, J.; Radermacher, P.; McCook, O. The Role of Glucocorticoid Receptor and Oxytocin Receptor in the Septic Heart in a Clinically Relevant, Resuscitated Porcine Model With Underlying Atherosclerosis. Front. Endocrinol. 2020, 11, 299. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.C.; Yang, H.; Lv, C.; Jia, S.; Liu, X.; Wang, X.; Meng, D.; Qin, D.; Zhu, H.; et al. Therapeutic Potential of Oxytocin in Atherosclerotic Cardiovascular Disease: Mechanisms and Signaling Pathways. Front. Neurosci. 2019, 13, 454. [Google Scholar] [CrossRef] [Green Version]
- Coletti, R.; Almeida-Pereira, G.; Elias, L.L.K.; Antunes-Rodrigues, J.E. Effects of hydrogen sulfide (H2S) on water intake and vasopressin and oxytocin secretion induced by fluid deprivation. Horm. Behav. 2015, 67, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Coletti, R.; de Lima, J.; Vechiato, F.; de Oliveira, F.L.; Debarba, L.K.; Almeida-Pereira, G.; Elias, L.; Antunes-Rodrigues, J. Nitric oxide acutely modulates hypothalamic and neurohypophyseal carbon monoxide and hydrogen sulphide production to control vasopressin, oxytocin and atrial natriuretic peptide release in rats. J. Neuroendocr. 2019, 31, e12686. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Thibonnier, M.; Conarty, D.M.; Preston, J.A.; Plesnicher, C.L.; Dweik, R.A.; Erzurum, S.C. Human vascular endothelial cells express oxytocin receptors. Endocrinology 1999, 140, 1301–1309. [Google Scholar] [CrossRef]
- Jankowski, M.; Wang, D.; Hajjar, F.; Mukaddam-Daher, S.; McCann, S.M.; Gutkowska, J. Oxytocin and its receptors are synthesized in the rat vasculature. Proc. Natl. Acad. Sci. USA 2000, 97, 6207–6211. [Google Scholar] [CrossRef] [Green Version]
- Menaouar, A.; Florian, M.; Wang, D.; Danalache, B.; Jankowski, M.; Gutkowska, J. Anti-hypertrophic effects of oxytocin in rat ventricular myocytes. Int. J. Cardiol. 2014, 175, 38–49. [Google Scholar] [CrossRef]
- Mancardi, D.; Pla, A.F.; Moccia, F.; Tanzi, F.; Munaron, L. Old and new gasotransmitters in the cardiovascular system: Focus on the role of nitric oxide and hydrogen sulfide in endothelial cells and cardiomyocytes. Curr. Pharm. Biotechnol. 2011, 12, 1406–1415. [Google Scholar] [CrossRef]
- Holst, S.; Uvnas-Moberg, K.; Petersson, M. Postnatal oxytocin treatment and postnatal stroking of rats reduce blood pressure in adulthood. Auton. Neurosci. 2002, 99, 85–90. [Google Scholar] [CrossRef]
- Całka, J. The role of nitric oxide in the hypothalamic control of LHRH and oxytocin release, sexual behavior and aging of the LHRH and oxytocin neurons. Folia Histochem. Cytobiol. 2006, 44, 3–12. [Google Scholar]
- Petersson, M.; Uvnas-Moberg, K. Postnatal oxytocin treatment of spontaneously hypertensive male rats decreases blood pressure and body weight in adulthood. Neurosci. Lett. 2008, 2, 166–169. [Google Scholar] [CrossRef]
- Ali, M.Y.; Ping, C.Y.; Mok, Y.Y.P.; Ling, L.; Whiteman, M.; Bhatia, M.; Moore, P.K. Regulation of vascular nitric oxide in vitro and in vivo: A new role for endogenous hydrogen sulphide? Br. J. Pharmacol. 2006, 149, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Dawe, G.S.; Han, S.P.; Bian, J.S.; Moore, P.K. Hydrogen sulphide in the hypothalamus causes an ATP-sensitive K channel-dependent decrease in blood pressure in freely moving rats. Neuroscience 2008, 152, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Q.; Chai, C.; Li, X.Y.; Yuan, W.J.; Wang, W.Z.; Lu, Y. The cardiovascular effects of central hydrogen sulfide are related to KATP channels activation. Physiol. Res. 2011, 60, 729–738. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trautwein, B.; Merz, T.; Denoix, N.; Szabo, C.; Calzia, E.; Radermacher, P.; McCook, O. ΔMST and the Regulation of Cardiac CSE and OTR Expression in Trauma and Hemorrhage. Antioxidants 2021, 10, 233. https://doi.org/10.3390/antiox10020233
Trautwein B, Merz T, Denoix N, Szabo C, Calzia E, Radermacher P, McCook O. ΔMST and the Regulation of Cardiac CSE and OTR Expression in Trauma and Hemorrhage. Antioxidants. 2021; 10(2):233. https://doi.org/10.3390/antiox10020233
Chicago/Turabian StyleTrautwein, Britta, Tamara Merz, Nicole Denoix, Csaba Szabo, Enrico Calzia, Peter Radermacher, and Oscar McCook. 2021. "ΔMST and the Regulation of Cardiac CSE and OTR Expression in Trauma and Hemorrhage" Antioxidants 10, no. 2: 233. https://doi.org/10.3390/antiox10020233