Effects of Storage Time and Temperature on Lipid Oxidation and Protein Co-Oxidation of Low-Moisture Shredded Meat Products

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and methods
2.1. Materials
2.2. Preparation of Shredded Meat Products
2.3. Physicochemical Quality Assessment on Shredded Meat Products
2.3.1. Proximate Composition
2.3.2. Reducing Sugar Content
2.3.3. Fatty Acid Composition
2.3.4. Water Activity, aw
2.3.5. Colour Measurement
2.4. Lipid Oxidation Analysis
2.4.1. Lipid Extraction and Total Lipid Content
2.4.2. Conjugated Dienes (CD)
2.4.3. Thiobarbituric Acid Reactive Substances (TBARS) Value
2.5. Protein Co-oxidation Analysis
2.5.1. Soluble Protein Content
2.5.2. Amino Acid Composition
2.5.3. Protein Carbonyl
2.5.4. Tryptophan Loss and Schiff Base Fluorescence Spectroscopy
2.6. Statistical Analysis
3. Results and Discussions
3.1. Physicochemical Analysis of Shredded Meat Products
3.1.1. Proximate Composition and aw
3.1.2. Fatty Acid Composition (FAC)
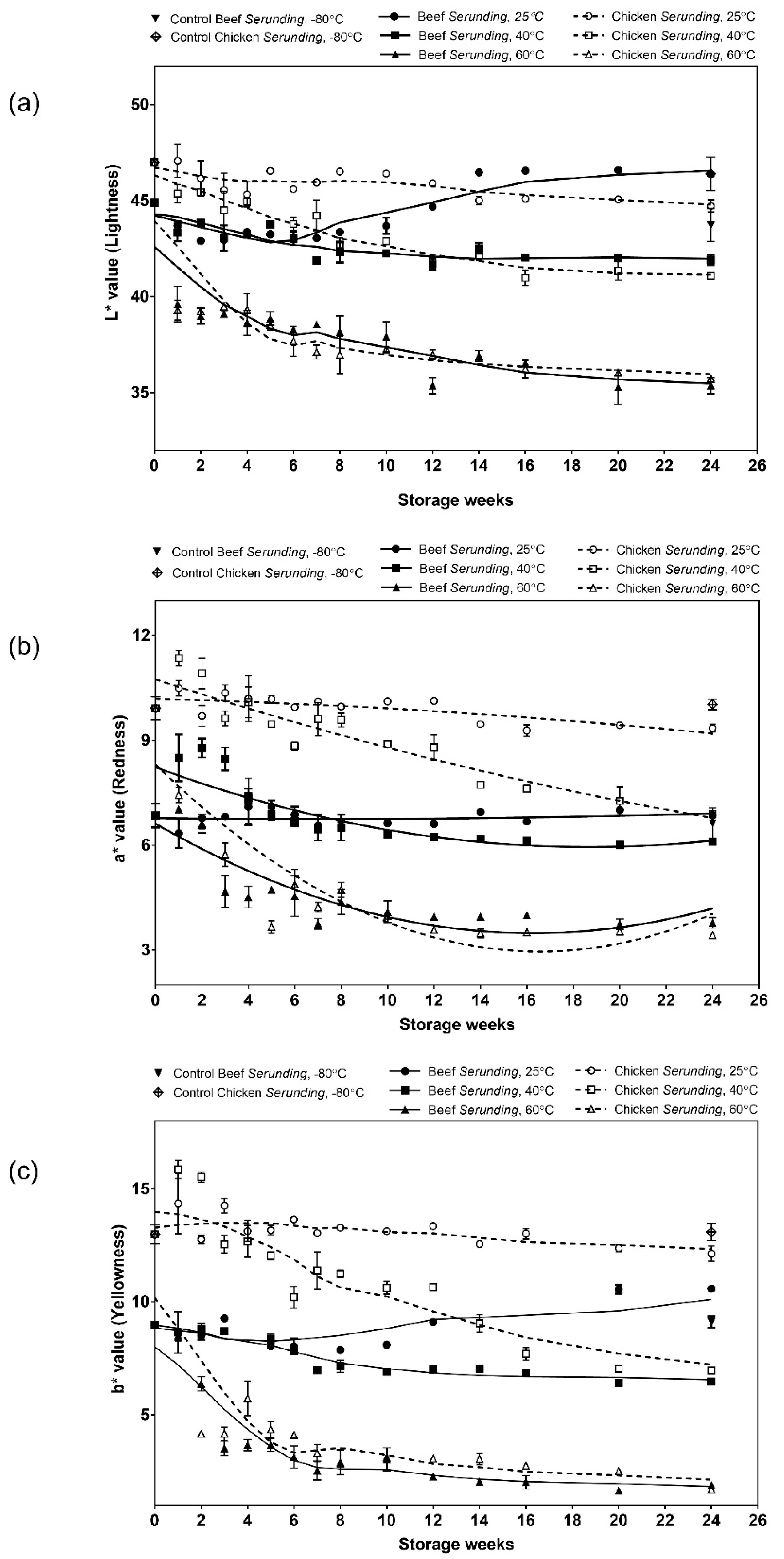
3.1.3. Colour
3.2. Lipid Oxidation
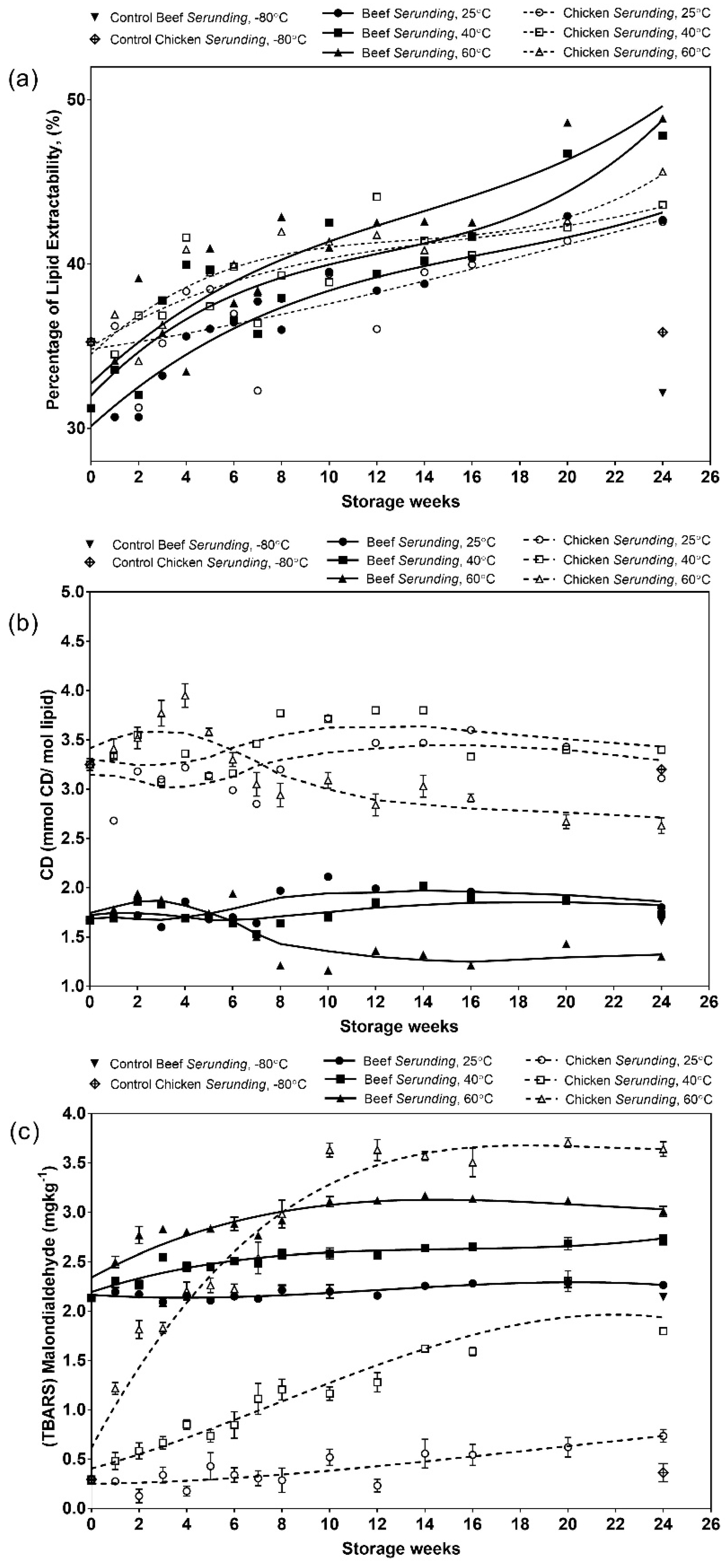
3.2.1. Extracted Lipid
3.2.2. Conjugated Dienes (CD)
3.2.3. Malondialdehydes (MDA)
3.3. Protein Co-oxidation
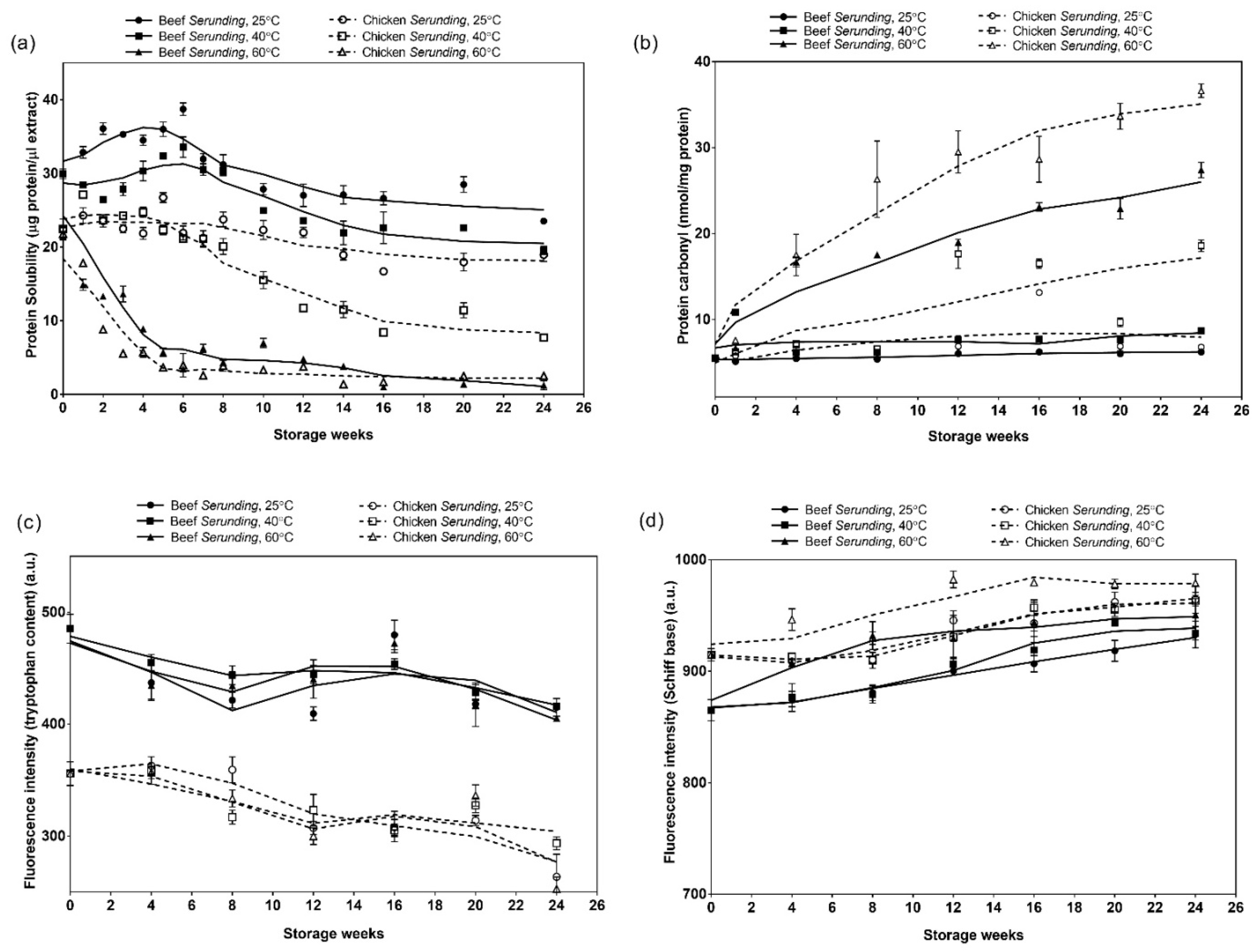
3.3.1. Soluble Protein Content
3.3.2. Amino Acid Composition
3.3.3. Protein Carbonyl
3.3.4. Tryptophan Loss and Schiff Base Fluorescence Spectroscopy
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baskaran, S.; Ayob, S.A.; Howe, N.C.; Mahadi, N. Understanding purchase intention of ready-to-eat food among Malaysian urbanites: A proposed framework. Int. J. Acad. Res. Bus. Soc. Sci. 2017, 7, 566–579. [Google Scholar] [CrossRef]
- Maity, T.; Bawa, A.; Raju, P. Optimization and quality assessment of ready-to-eat intermediate moisture compressed beetroot bar. J. Food Sci. Technol. 2016, 53, 3233–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, S.; Adams, J.; White, M. Nutritional content of supermarket ready meals and recipes by television chefs in the United Kingdom: Cross sectional study. Br. Med J. 2012, 345, e7607. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Verma, A.K.; Kumar, D.; Umaraw, P.; Mehta, N.; Malav, O. Meat snacks: A novel technological perspective. In Innovations in Traditional Foods; Elsevier Inc.: London, UK, 2019; pp. 293–321. [Google Scholar]
- Price, A.; Díaz, P.; Bañón, S.; Garrido, M.D. Natural extracts versus sodium ascorbate to extend the shelf life of meat-based ready-to-eat meals. Food Sci. Technol. Int. 2013, 19, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Hu, M. Oxidative stability and shelf life of low-moisture foods. In Oxidative Stability and Shelf Life of Foods Containing Oils and Fats; Elsevier Inc.: London, UK, 2016; pp. 313–371. [Google Scholar]
- Schaich, K.M. Co-oxidation of proteins by oxidizing lipids. In Lipid Oxidation Pathways; AOCS Press: Urbana, IL, USA, 2008; pp. 183–274. [Google Scholar]
- Barden, L.M. Understanding lipid oxidation in low-moisture food. Ph.D. Thesis, University of Massachusetts Amherst, Amherst, MA, USA, 2014. [Google Scholar]
- Tournour, H.H.; Segundo, M.A.; Magalhães, L.M.; Costa, A.S.; Cunha, L.M. Effect of Touriga nacional grape extract on characteristics of mechanically deboned chicken meat kept under frozen storage. J. Food Process. Eng. 2017, 40, e12434. [Google Scholar] [CrossRef]
- Estévez, M.; Xiong, Y. Intake of oxidized proteins and amino acids and causative oxidative stress and disease: Recent scientific evidences and hypotheses. J. Food Sci. 2019, 84, 387–396. [Google Scholar] [CrossRef]
- Villaverde, A.; Estévez, M. Carbonylation of myofibrillar proteins through the Maillard pathway: Effect of reducing sugars and reaction temperature. J. Agric. Food Chem. 2013, 61, 3140–3147. [Google Scholar] [CrossRef]
- AOAC International. Official methods of analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists (AOAC) International: Gaithersburg, MD, USA, 2010. [Google Scholar]
- Utrera, M.; Parra, V.; Estévez, M. Protein oxidation during frozen storage and subsequent processing of different beef muscles. Meat Sci. 2014, 96, 812–820. [Google Scholar] [CrossRef]
- AOCS. AOCS Official Method Ce 1-62; The American Oil Chemists’ Society: Urbana, IL, USA, 2005. [Google Scholar]
- Ibadullah, W.Z.W. Lipid protein interactions in peanut butter. Ph.D. Thesis, Rutgers University, New Brunswick, NJ, USA, 2013. [Google Scholar]
- AOCS. AOCS Th 1a-64; The American Oil Chemists’ Society: Urbana, IL, USA, 2011. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shang Gui, D.; Zhi Ying, P.; Fang, C.; Ping, Y.; Tie, W. Amino acid composition and anti-anaemia action of hydrolyzed offal protein from Harengula Zunasi Bleeker. Food Chem. 2004, 87, 97–102. [Google Scholar] [CrossRef]
- Soglia, F.; Petracci, M.; Ertbjerg, P. Novel DNPH-based method for determination of protein carbonylation in muscle and meat. Food Chem. 2016, 197, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. In Methods in Enzymology; Elsevier Inc.: London, UK, 1994; pp. 346–357. [Google Scholar]
- Wehr, N.B.; Levine, R.L. Quantification of protein carbonylation. In Methods in Molecular Biology; Clifton, N.J., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 265–281. [Google Scholar]
- FAO-WHO. Code of Hygienic Practice for Low-Moisture Foods (CXC 75-2015). In Codex Alimentarius: International Food Standards; Codex Secretariat: Rome, Italy, 2015. [Google Scholar]
- Huda, N.; Fatma, Y.; Fazillah, A.; Adzitey, F. Chemical composition, colour and sensory characteristics of commercial serunding (Shredded Meat) in Malaysia. Pak. J. Nutr. 2012, 11, 1–4. [Google Scholar] [CrossRef]
- Sukisman, A.; Rahim, A. Sifat fisik, kimia dan aktivitas antioksidan abon daging ayam di Kota Palu. J. Agroland. 2018, 25, 154–163. [Google Scholar]
- Sukisman, A.; Purnomo, H.; Rosyidi, D.; L, R. Quality properties, antioxidant capacity and total phenolic content of traditional deep fried shredded meat (abon) of Palu, Central Sulawesi. Am. J. Food Technol. 2014, 9, 80–88. [Google Scholar]
- Sun, Q.; Senecal, A.; Chinachoti, P.; Faustman, C. Effect of water activity on lipid oxidation and protein solubility in freeze-dried beef during storage. J. Food Sci. 2002, 67, 2512–2516. [Google Scholar] [CrossRef]
- Cheng, H.; Erichsen, H.; Soerensen, J.; Petersen, M.A.; Skibsted, L.H. Optimising water activity for storage of high lipid and high protein infant formula milk powder using multivariate analysis. Int. Dairy J. 2019, 93, 92–98. [Google Scholar] [CrossRef]
- Rial, S.; Karelis, A.; Bergeron, K.-F.; Mounier, C. Gut microbiota and metabolic health: The potential beneficial effects of a medium chain triglyceride diet in obese individuals. Nutrients 2016, 8, 281. [Google Scholar] [CrossRef]
- Khaw, K.-T.; Sharp, S.J.; Finikarides, L.; Afzal, I.; Lentjes, M.; Luben, R.; Forouhi, N.G. Randomised trial of coconut oil, olive oil or butter on blood lipids and other cardiovascular risk factors in healthy men and women. Br. Med. J. 2018, 8, e020167. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Z.; Han, Y.; Xu, J.; Huang, W.; Li, Z. Medium Chain Triglycerides enhances exercise endurance through the increased mitochondrial biogenesis and metabolism. PLoS ONE 2018, 13, e0191182. [Google Scholar] [CrossRef]
- Carrascal, J.R.; Hidalgo, J.; Morillo, M. Fatty acid composition of phospholipid classes in beef. In Proceedings of the 3rd Euro Fed Lipid Congress, Edinburgh, Scotland, 5–8 September 2004. [Google Scholar]
- Rhee, K.; Anderson, L.; Sams, A. Lipid oxidation potential of beef, chicken, and pork. J. Food Sci. 1996, 61, 8–12. [Google Scholar] [CrossRef]
- Moreno, M.M.; Olivares, D.M.; Lopez, F.A.; Adelantado, J.G.; Reig, F.B. Analytical evaluation of polyunsaturated fatty acids degradation during thermal oxidation of edible oils by Fourier transform infrared spectroscopy. Talanta 1999, 50, 269–275. [Google Scholar] [CrossRef]
- Azima, F. Chemical characteristic and fatty acid profile in rendang of minangkabau. Int. J. Adv. Sci. Eng. Inf. Technol. 2016, 6, 465–468. [Google Scholar] [CrossRef]
- Suman, S.P.; Joseph, P. Myoglobin chemistry and meat color. Annu. Rev. Food Sci. Technol. 2013, 4, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, G.; Saldanha, T.; Soares, R.; Torres, E. Effect of natural antioxidant combinations on lipid oxidation in cooked chicken meat during refrigerated storage. Food Chem. 2012, 135, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Nissen, L.R.; Månsson, L.; Bertelsen, G.; Huynh-Ba, T.; Skibsted, L.H. Protection of dehydrated chicken meat by natural antioxidants as evaluated by electron spin resonance spectrometry. J. Agric. Food Chem. 2000, 48, 5548–5556. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.; Marangoni, C.; Brum, D.; Vendruscolo, R.; Silva, M.; de Moura, H.; Rampelotto, C.; Wagner, R.; de Menezes, C.; Barin, J. Effect of dietary olive leaves on the lipid and protein oxidation and bacterial safety of chicken hamburgers during frozen storage. Int. Food Res. J. 2018, 25, 383–391. [Google Scholar]
- Uchida, K.; Sakai, K.; Itakura, K.; Osawa, T.; Toyokuni, S. Protein modification by lipid peroxidation products: Formation of malondialdehyde-derivednϵ-(2-propenal) lysine in proteins. Arch. Biochem. Biophys. 1997, 346, 45–52. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Emara, A.; Gan, X.; Li, H. Effects of malondialdehyde as a byproduct of lipid oxidation on protein oxidation in rabbit meat. Food Chem. 2019, 288, 405–412. [Google Scholar] [CrossRef]
- Sohaib, M.; Anjum, F.M.; Arshad, M.S.; Imran, M.; Imran, A.; Hussain, S. Oxidative stability and lipid oxidation flavoring volatiles in antioxidants treated chicken meat patties during storage. Lipids Health Dis. 2017, 16, 27. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y. Protein modifications in baked versus fried tortilla chips. Master’s Thesis, Rutgers University, New Brunswick, NJ, USA, 2011. [Google Scholar]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef]
- Nollet, L.; Toldra, F. Handbook of Processed Meats and Poultry Analysis; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Carvalho, R.H.; Ida, E.I.; Madruga, M.S.; Martínez, S.L.; Shimokomaki, M.; Estévez, M. Underlying connections between the redox system imbalance, protein oxidation and impaired quality traits in pale, soft and exudative (PSE) poultry meat. Food Chem. 2017, 215, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Soladoye, O.; Juárez, M.; Aalhus, J.; Shand, P.; Estévez, M. Protein oxidation in processed meat: Mechanisms and potential implications on human health. Compr. Rev. Food Sci. Food Saf. 2015, 14, 106–122. [Google Scholar] [CrossRef]
- Gatellier, P.; Kondjoyan, A.; Portanguen, S.; Sante-Lhoutellier, V. Effect of cooking on protein oxidation in n-3 polyunsaturated fatty acids enriched beef. Implication on nutritional quality. Meat Sci. 2010, 85, 645–650. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Component. | Present Data | Reported Data* | ||
|---|---|---|---|---|
| Beef | Chicken | Beef | Chicken | |
| Moisture | 6.36 ± 0.03 a | 7.46 ± 0.07 b | 4.20–12.12 | 4.32–13.56 |
| Protein | 23.60 ± 0.02 a | 21.77 ± 0.12 b | 19.86–36.39 | 29.71–40.72 |
| Fat | 31.99 ± 0.10 a | 32.30 ± 0.36 a | 3.20–39.00 | 6.04–21.98 |
| Ash | 5.20 ± 0.01 a | 4.97 ± 0.09 b | 4.08–5.16 | 3.17–5.91 |
| Carbohydrate (by difference) | 32.85 ± 0.15 a | 33.50 ± 0.53 a | N/A | N/A |
| Water activity Reducing sugar | 0.381 ± 0.003 a 1.09 ± 0.027 a | 0.431 ± 0.002 b 1.02 ± 0.033 b | 0.410–0.640 N/A | N/A N/A |
| Present Data | Reported Data | |||||
|---|---|---|---|---|---|---|
| Sample | Beef Serunding | Chicken Serunding | Raw | Cooked | ||
| Beef | Chicken | Beef | Chicken | |||
| C6:0 | 0.67 ± 0.01 a | 0.73 ± 0.01 b | - | - | - | - |
| C8:0 | 7.72 ± 0.10 a | 8.43 ± 0.08 b | - | - | - | - |
| C10:0 | 5.85 ± 0.05 a | 6.50 ± 0.09 b | - | - | - | - |
| C12:0 | 41.78 ± 0.33 a | 47.85 ± 0.70 b | - | - | - | - |
| C14:0 | 15.51 ± 0.08 a | 17.29 ± 0.17 b | - | - | - | - |
| C14:1 | 0.10 ± 0.01 | - | - | - | - | - |
| C16:0 | 11.01 ± 0.10 b | 8.32 ± 0.37 a | - | - | - | - |
| C16:1 | 0.45 ± 0.00 b | 0.11 ± 0.01 a | - | - | - | - |
| C17:0 | 0.20 ± 0.00 | - | - | - | - | - |
| C18:0 | 4.98 ± 0.14 b | 2.44 ± 0.29 a | - | - | - | - |
| C18:1, cis-9 | 5.29 ± 0.22 a | 5.98 ± 0.61 b | - | - | - | - |
| C18:2, cis-9,12 | 1.45 ± 0.11 a | 4.12 ± 0.61 b | 3.07 | - | - | - |
| Saturated | 87.73 ± 0.17 a | 91.65 ± 0.45 b | 48.65 | 31.27 | 40.15 | 32.11 |
| Monounsaturated | 5.84 ± 0.16 a | 6.09 ± 0.63 b | 47.65 | 43.31 | 50.76 | 45.76 |
| Polyunsaturated | 1.45 ± 0.11 a | 4.12 ± 0.05 b | 3.70 | 18.87 | 5.50 | 20.53 |
| Amino Acid (mg/g Protein) | Beef Serunding | Chicken Serunding | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Week 0 | Week 24 | Week 0 | Week 24 | ||||||
| Control | 25 ± 1 ⁰C | 40 ± 1 ⁰C | 60 ± 1 ⁰C | Control | 25 ± 1 ⁰C | 40 ± 1 ⁰C | 60 ± 1 ⁰C | ||
| 1 | Glycine | 12.69 ± 1.32 a | 12.57 ± 0.36 a | 10.94 ± 0.05 a | 11.86 ± 0.03 a | 9.95 ± 0.12 a | 9.52 ± 0.05 b | 8.74 ± 0.10 c | 9.61 ± 0.13 ab |
| 2 | Histidine | 12.97 ± 2.55 a | 7.18 ± 0.19 b | 6.58 ± 0.52 b | 6.38 ± 0.03 b | 6.75 ± 0.33 a | 6.16 ± 0.27 ab | 6.82 ± 0.11 a | 5.55 ± 0.14 b |
| 3 | Arginine | 27.64 ± 0.23 a | 21.88 ± 0.93 b | 18.01 ± 0.38 c | 16.70 ± 0.22 c | 23.46 ± 0.50 a | 20.68 ± 0.01 ab | 20.69 ± 0.27 ab | 19.97 ± 0.04 b |
| 4 | Threonine | 20.70 ± 0.98 a | 13.60 ± 0.36 b | 11.02 ± 0.16 c | 13.39 ± 0.09 b | 11.16 ± 1.22 a | 12.47 ± 0.17 a | 11.42 ± 0.09 a | 10.79 ± 0.01 a |
| 5 | Alanine | 21.33 ± 0.27 a | 19.09 ± 0.03 b | 15.71 ± 0.32 c | 15.61 ± 0.09 c | 23.28 ± 0.32 a | 20.65 ± 0.26 b | 20.10 ± 0.04 b | 14.38 ± 0.04 c |
| 6 | Proline | 11.05 ± 0.20 bc | 13.46 ± 0.51 a | 10.26 ± 0.24 c | 11.72 ± 0.11 b | 10.24 ± 0.47 a | 10.45 ± 0.11 a | 10.55 ± 0.17 a | 10.11 ± 0.0 a |
| 7 | Tyrosine | 17.18 ± 2.11 a | 11.49 ± 0.00 b | 8.87 ± 0.09 b | 11.51 ± 0.07 b | 10.62 ± 0.31 a | 9.09 ± 0.04 b | 8.01 ± 0.30 c | 8.61 ± 0.03 bc |
| 8 | Methionine | 9.95 ± 0.32 a | 6.27 ± 0.05 bc | 4.61 ± 0.46 c | 7.47 ± 0.64 b | 6.64 ± 0.01 a | 6.74 ± 0.25 b | 6.07 ± 0.01 b | 5.29 ± 0.35 b |
| 9 | Isoleucine | 13.75 ± 1.42 a | 13.75 ± 0.18 a | 10.29 ± 0.17 b | 12.68 ± 0.11 ab | 12.73 ± 0.26 a | 11.47 ± 0.17 b | 10.77 ± 0.39 b | 10.56 ± 0.00 b |
| 10 | Leucine | 16.36 ± 0.37 d | 25.40 ± 0.26 a | 18.72 ± 0.21 c | 21.69 ± 0.25 b | 19.35 ± 0.03 a | 17.32 ± 0.53 b | 18.14 ± 0.24a b | 17.33 ± 0.41 b |
| 11 | Phenylalanine | 119.59 ± 0.21 a | 114.27 ± 0.30 b | 63.50 ± 0.32 c | 61.80 ± 0.21 d | 98.55 ± 0.12 a | 98.37 ± 1.04 a | 85.10 ± 0.28 b | 83.27 ± 0.04 b |
| 12 | Lysine | 56.67 ± 0.43 a | 36.63 ± 0.24 b | 26.63 ± 0.96 c | 24.48 ± 0.36 d | 29.82 ± 0.09 a | 25.14 ± 0.71 b | 24.65 ± 0.11 b | 23.79 ± 0.14 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wazir, H.; Chay, S.Y.; Zarei, M.; Hussin, F.S.; Mustapha, N.A.; Wan Ibadullah, W.Z.; Saari, N. Effects of Storage Time and Temperature on Lipid Oxidation and Protein Co-Oxidation of Low-Moisture Shredded Meat Products. Antioxidants 2019, 8, 486. https://doi.org/10.3390/antiox8100486
Wazir H, Chay SY, Zarei M, Hussin FS, Mustapha NA, Wan Ibadullah WZ, Saari N. Effects of Storage Time and Temperature on Lipid Oxidation and Protein Co-Oxidation of Low-Moisture Shredded Meat Products. Antioxidants. 2019; 8(10):486. https://doi.org/10.3390/antiox8100486
Chicago/Turabian StyleWazir, Hazrati, Shyan Yea Chay, Mohammad Zarei, Farah Salina Hussin, Nor Afizah Mustapha, Wan Zunairah Wan Ibadullah, and Nazamid Saari. 2019. "Effects of Storage Time and Temperature on Lipid Oxidation and Protein Co-Oxidation of Low-Moisture Shredded Meat Products" Antioxidants 8, no. 10: 486. https://doi.org/10.3390/antiox8100486