
Chitosan-Loaded Lagenaria siceraria and Thymus vulgaris Potentiate Antibacterial, Antioxidant, and Immunomodulatory Activities against Extensive Drug-Resistant Pseudomonas aeruginosa and Vancomycin-Resistant Staphylococcus aureus: In Vitro and In Vivo Approaches
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antimicrobial Susceptibility Testing of Bacterial Strains
2.3. Natural Plants and Chitosan
2.4. Extraction of Lagenaria siceraria and Thymus vulgaris Essential Oils
2.5. Gas Chromatography–Mass Spectrometry Analysis (GC-MS)
2.6. Preparation of Essential Oil-Loaded Chitosan Nanoparticles [27]
2.7. Characterization of Essential Oil-Loaded Chitosan Nanoparticles
2.8. Antibacterial Activity of Chitosan Nanoparticles Loaded Either Lagenaria siceraria or Thymus vulgaris
2.9. Experimental Animals
2.10. Experimental Design
2.11. Biochemical and Serum Analysis
2.12. Measurement of Hematological Parameters
2.13. Enumeration of Bacteria
2.14. Pathological Examination
2.15. Real-Time Quantitative Polymerase Chain Reaction (RT–qPCR)
2.16. Statistical Analysis
3. Results
3.1. Antibiogram of P. aeruginosa and VRSA Strains
3.2. Characterization of Lagenaria siceraria and Thymus vulgaris Using GC-MS Analysis
3.3. Transmission Electron Microscopy and FTIR for Characterization of the Nanoemulsion
3.4. Antimicrobial Activities of the Essential Oils and Their Nanoemulsions against XDR P. aeruginosa and VRSA Strains
3.5. Results of the In Vivo Study
3.5.1. Clinical Observation and Postmortem Lesions
3.5.2. Results of Biochemical and Hematological Parameters
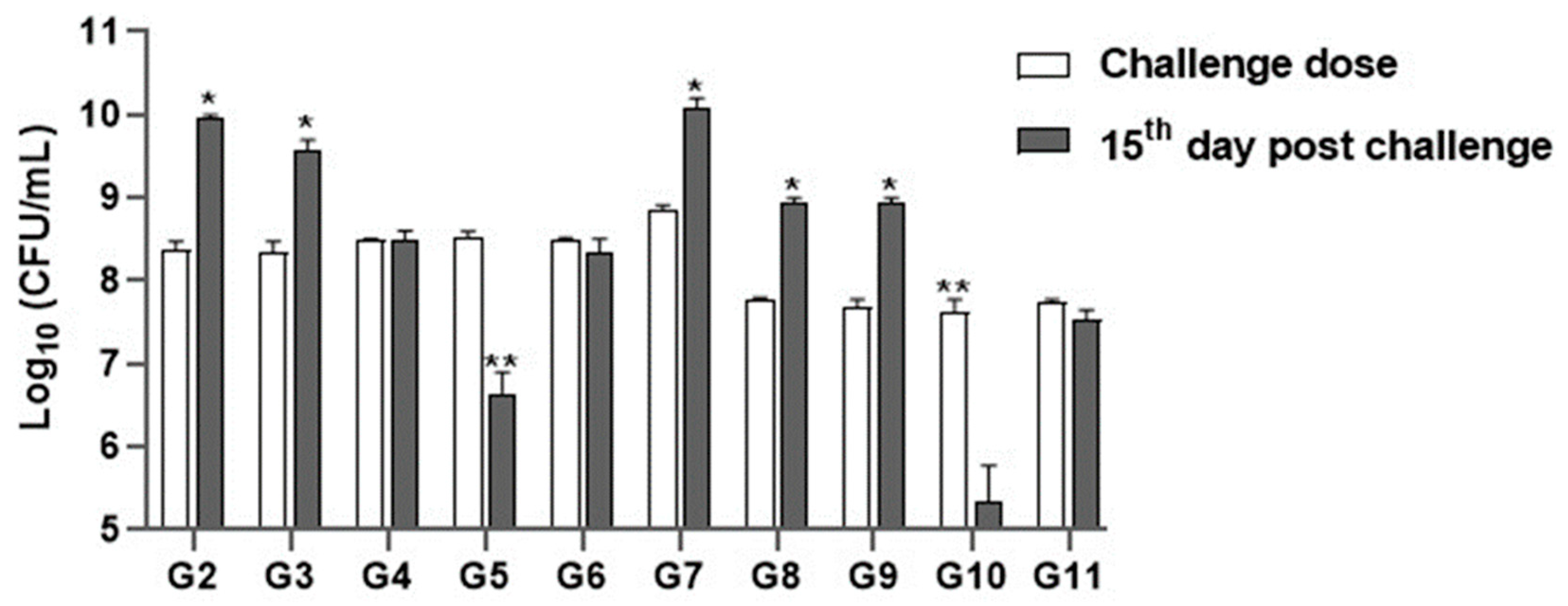
3.5.3. Microbial Count Analysis
3.5.4. Histopathological Findings
3.5.5. Transcriptional Modulation of IL10 and IFNG Cytokine Genes in the Experimental Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World health organization (WHO). Newsroom, Antimicrobial Resistance. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance, (accessed on 22 April 2023).
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials—A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe; ECDC: Solna Municipality, Sweden, 2017. [Google Scholar]
- Shrestha, P.; Cooper, B.S.; Coast, J.; Oppong, R.; Thuy, N.D.T.; Phodha, T.; Celhay, O.; Guerin, P.J.; Wertheim, H.; Lubell, Y. Enumerating the economic cost of antimicrobial resistance per antibiotic consumed to inform the evaluation of interventions affecting their use. Antimicrob. Resist. Infect. Control 2018, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Butucel, E.; Balta, I.; Ahmadi, M.; Dumitrescu, G.; Morariu, F.; Pet, I.; Stef, L.; Corcionivoschi, N. Biocides as Biomedicines against Foodborne Pathogenic Bacteria. Biomedicines 2022, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Bundurus, I.A.; Balta, I.; Stef, L.; Ahmadi, M.; Pet, I.; McCleery, D.; Corcionivoschi, N. Overview of virulence and antibiotic resistance in Campylobacter spp. livestock isolates. Antibiotics 2023, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Eugene, S.S.; Lam, J.S. Pseudomonas aeruginosa antigens as potential vaccines FEMS. Microbiol. Rev. 1997, 21, 243–277. [Google Scholar]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Sievert, D.M.; Hageman, J.C. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N. Engl. J. Med. 2003, 348, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Weigel, L.M.; Clewell, D.B.; Gill, S.R. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef] [PubMed]
- Rota, M.C.; Herrera, A.; Martínez, R.M.; Sotomayor, J.A.; Jordán, M.J. Antimicrobial activity and chemical composition of Thymus vulgaris, Thymus zygis and Thymus hyemalis essential oils. Food Control 2008, 19, 681–687. [Google Scholar] [CrossRef]
- Said, P.P.; Pradhan, R.C.; Rai, B.N. A green separation of Lagenaria siceraria seed oil. Ind. Crops Prod. 2014, 52, 796–800. [Google Scholar] [CrossRef]
- Shah, B.N.; Seth, A.K.; Desai, R.V. Phytopharmacological profile of Lagenaria siceraria: A review. Asian J. Plant Sci. 2010, 9, 152–157. [Google Scholar] [CrossRef]
- Al-Zahrani, S.S.; Bora, R.S.; Al-Garni, S.M. Antimicrobial activity of chitosan nanoparticles. Biotechnol. Biotechnol. Equip. 2021, 35, 1874–1880. [Google Scholar] [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial properties of chitosan nanoparticles: Mode of action and factors affecting activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Syeda, J.T.M.; Wasan, K.M.; Wasan, E.K. An overview of chitosan nanoparticles and its application in non-parenteral drug delivery. Pharmaceutics 2017, 9, 53. [Google Scholar] [CrossRef]
- Quinn, P.J.; Carter, M.E.; Markey, B.; Carter, G.R. Clinical Veterinary Microbiology, 1st ed.; London Wolfe Publishing: Cheltenham, UK, 1994; pp. 21–66. [Google Scholar]
- Gad, G.F.; El-Domany, R.A.; Zaki, S.; Ashour, H.M. Characterization of Pseudomonas aeruginosa isolated from clinical and environmental samples in Minia, Egypt: Prevalence, antibiogram and resistance mechanisms. J. Antimicrob. Chemother. 2007, 60, 1010–1017. [Google Scholar] [CrossRef]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of S. aureus by polymerase chain reaction amplification of the nuc gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Truck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, E.; Lassa, H.; Smulski, S.; Kłossowska, A.; Kaczmarowski, M. Antimicrobial susceptibility of bacteria isolated from cows with mastitis in 2006–2007. Bull. Vet. Inst. Puławy 2008, 4, 52. [Google Scholar]
- Rankin, D.I. MIC Testing. In Manual of Antimicrobial Susceptibility Testing; American Society for Microbiology: Seattle, WA, USA, 2005; pp. 53–62. [Google Scholar]
- CLSI. Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Doc. M100; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Essien, E.E.; Antia, B.S.; Udoh, B.I. Phytochemical screening and antimicrobial activity of Lagenaria siceraria seeds extracts. Int. J. Pharm. Phytochem. Res. 2015, 7, 554–558. [Google Scholar]
- Castro, D.P.; Cardoso, M.G.; Moraes, J.C.; Santos, N.M.; Baliza, D.P. Não-preferência de Spodoptera frugiperda (Lepidoptera: Noctuidae) por óleos essenciais de Achillea millefolium L. e Thymus vulgaris L. Rev. Bras. Plantas Med. 2006, 8, 27–32. [Google Scholar]
- Millezi, A.F.; Caixeta, D.S.; Rossoni, F.; Cardoso, M.d.G.; Piccoli, R.H. In vitro antimicrobial properties of plant essential oils Thymus vulgaris, Cymbopogon citratus and Laurus nobilis against five important foodborne pathogens Original Papers. Food Sci. Technol. 2012, 32, 167–172. [Google Scholar] [CrossRef]
- Hadidia, M.; Pouraminb, S.; Adinepourc, F.; Haghanib, S.; Jafaric, S.M. Chitosan nanoparticles loaded with clove essential oil: Characterization, antioxidant and antibacterial activities. Carbohydr. Polym. 2020, 236, 116075. [Google Scholar] [CrossRef] [PubMed]
- Keawchaoon, L.; Yoksan, R. Preparation characterization and in vitro release study of carvacrol-loaded chitosan nanoparticles. Colloids Surf. B Biointerfaces 2011, 84, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, X.; Xiao, Z.; Bi, W. Effect of chitosan nanoparticles loaded with cinnamon essential oil on the quality of chilled pork. LWT-Food Sci. Technol. 2015, 63, 519–526. [Google Scholar] [CrossRef]
- Ghaderi-Ghahfarokhi, M.; Barzegar, M.; Sahari, M.A.; Azizi, M.H. Nanoencapsulation approach to improve antimicrobial and antioxidant activity of thyme essential oil in beef burgers during refrigerated storage. Food Bioprocess Technol. 2016, 9, 1187–1201. [Google Scholar] [CrossRef]
- Mohd Yusof, H.; Abdul Rahman, N.A.; Mohamad, R.; Zaidan, U.H. Microbial mediated synthesis of silver nanoparticles by Lactobacillus Plantarum TA4 and its antibacterial and antioxidant activity. Appl. Sci. 2020, 10, 6973. [Google Scholar] [CrossRef]
- Hamilton-Miller, J.M.T. Calculating MIC50. J. Antimicrob. Chemother. 1991, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Laboratory Animals; National Academies Press (US): Washington, DC, USA, 1995. [Google Scholar]
- Jankie, S.; Jenelle, J.; Suepaul, R.; Pereira, L.P.; Akpaka, P.; Sarafadeen, A.; Adebayo, A.S.; Pillai, G. Determination of the infective dose of Staphylococcus aureus (ATCC 29213) and Pseudomonas aeruginosa (ATCC 27853) when injected intraperitoneally in Sprague Dawley rats. Br. J. Pharm. Res. 2016, 14, 1–11. [Google Scholar] [CrossRef]
- Hassanen, I.E.; Ragab, E. In vivo and in vitro assessments of the antibacterial potential of chitosan-silver nanocompositeagainst methicillin-resistant Staphylococcus aureus–induced infection in rats. Biol. Trace Elem. Res. 2021, 199, 244–257. [Google Scholar] [CrossRef]
- Channabasappa, S.; Durgaiah, M.; Chikkamadaiah, R.; Kumar, S.; Joshi, A.; Sriram, B. Efficacy of novel antistaphylococcal ectolysin P128 in a rat model of methicillin-resistant Staphylococcus aureus bacteremia. Antimicrob. Agents Chemother. 2018, 62, e01358-17. [Google Scholar] [CrossRef]
- Weatherby, D.; Ferguson, S. Blood Chemistry and CBC Analysis: Clinical Laboratory Testing from a Functional Perspective, 1st ed.; Vis Medicatrix Press: Jacksonville, OR, USA, 2002. [Google Scholar]
- Gross, R.T.; Bracci, R.; Rudolph, N.; Schroeder, E.; Kochen, J.A. Hydrogen peroxide toxicity and detoxification in the erythro-cytes. Blood 1967, 29, 481–493. [Google Scholar] [CrossRef]
- Nishikimi, M.; Roa, N.A.; Yogi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulphate andmolecular oxygen. Biochem. Bioph. Res. Common. 1972, 46, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Dong, B.; Wang, K.; Cai, S.; Liu, T.; Cheng, X.; Lei, D.; Chen, Y.; Li, Y.; Kong, J.; et al. Baicalin inhibits biofilm formation, attenuates the quorum sensing-controlled virulence and enhances Pseudomonas aeruginosa clearance in a mouse peritoneal implant infection model. PLoS ONE 2017, 12, e0176883. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.; Floyd, A.; Suvarna, S. Bancroft’s Theory and Practice of Histological Techniques, 7th ed.; Churchill Livingstone: London, UK, 2013. [Google Scholar]
- Banni, M.; Messaoudi, I.; Said, L.; El Heni, J.; Kerkeni, A.; Said, K. Metallothionein gene expression in liver of rats exposed to cadmium and supplemented with zinc and selenium. Arch. Environ. Contam. Toxicol. 2010, 59, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Shynlova, O.; Dorogin, A.; Li, Y.; Lye, S. Inhibition of infection-mediated preterm birth by administration of broad-spectrum chemokine inhibitor in mice. J. Cell. Mol. Med. 2014, 18, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- Sisto, F.; Miluzio, A.; Leopardi, Q.; Mirra, M.; Boelaert, J.R.; Taramelli, D. Differential cytokine pattern in the spleens and livers of BALB/c mice infected with Penicillium marneffei: Protective role of gamma interferon. Infect. Immun. 2003, 71, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]
- Razali, N.M.; Wah, Y.B. Power comparisons of shapiro-wilk, kolmogorov-smirnov, lilliefors and anderson-darling tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]
- SAS Institute Inc. SAS/STAT Statistics User’s Guide. Statistical Analytical System, 5th rev. ed.; SAS Institute Inc: Cary, NC, USA, 2012. [Google Scholar]
- Abdelatti, M.A.I.; Abd El-Aziz, N.K.; El-Naenaeey, E.-s.Y.M.; Ammar, A.M.; Alharbi, N.K.; Alharthi, A.; Zakai, S.A.; Abdelkhalek, A. Antibacterial and anti-efflux activities of cinnamon essential oil against pan and extensive drug-resistant Pseudomonas aeruginosa isolated from human and animal sources. Antibiotics 2023, 12, 1514. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aureus recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55 (Suppl. S20), 221–230. [Google Scholar]
- Standing Medical Advisory Committee, Sub-Group in Antimicrobial Resistance, Department of Health, Edinburgh University Library Catalogue: Wetherby, UK, 1998. Available online: https://discovered.ed.ac.uk/permalink/44UOE_INST/iatqhp/alma998008053502466 (accessed on 11 May 2023).
- Naik, P.; Pandey, S.; Gagan, S.; Biswas, S.; Joseph, J. Virulence factors in multidrug (MDR) and pan-drug resistant (XDR) Pseudomonas aeruginosa: A cross-sectional study of isolates recovered from ocular infections in a high-incidence setting in southern India. J. Ophthalmic Inflamm. Infect. 2021, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Almanaa, T.N.; Alyahya, S.A.; Khaled, J.M.; Shehu, M.R.; Alharbi, N.S.; Kadaikunnan, S.; Alzahrani, A.K. The extreme drug resistance (XDR) Staphylococcus aureus strains among patients: A retrospective study. Saudi J. Biol. Sci. 2020, 27, 1985–1992. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, D.; Ashiq, N. Lagenaria siceraria fruit pedicle extracts as a remedy against microbial infections. Int. J. Veg. Sci. 2018, 24, 539–549. [Google Scholar] [CrossRef]
- Nagaraja, Y.P.; Geetha, K.N.; Vinay, M.S. Antimicrobial effect of Lagenaria siceraria (Mol.) Standley, against certain bacterial and fungal strains. J. Appl. Nat. Sci. 2011, 3, 124–127. [Google Scholar]
- Diniz, A.F.; Santos, B.; Nóbrega, L.M.M.O.; Santos, V.R.L.; Mariz, W.S.; Cruz, P.S.C.; Oliveira-Filho, A.A. Antibacterial activity of Thymus vulgaris (thyme) essential oil against strains of Pseudomonas aeruginosa, Klebsiella pneumoniae and Staphylococcus saprophyticus isolated from meat product. Braz. J. Biol. 2023, 83, e275306. [Google Scholar] [CrossRef] [PubMed]
- Dar, P.; Dar, A.; Dar, A.M.; Waqas, U. Synthesis of silver nanoparticles of mesocarp of Lagenaria siceraria fruit by using oleylamine and evaluation of its antimicrobial potential against clinically used pathogens. Int. J. Mod. Biol. Res. 2017, 5, 42–55. [Google Scholar]
- Kim, N.N.; Stankovic, M.; Cushman, T.T. Streptozotocin-induced diabetes in the rat is associated with changes in vaginal hemodynamics, morphology and biochemical markers. BMC Physiol. 2006, 6, 4. [Google Scholar] [CrossRef]
- Udayakumar, R.; Kasthurirengan, S.; Mariashibu, T.S.; Rajesh, M.; Anbazhagan, V.R. Hypoglycaemic and hypolipidaemic effects of Withania somnifera root and leaf extracts on alloxan-induced diabetic rats. Int. J. Mol. Sci. 2009, 10, 2367–2382. [Google Scholar] [CrossRef]
- Rahman, A.N.A.; Abdelwarith, A.A.; Younis, E.M.; Rhouma, N.R.; Zaki, H.T.; Khalil, S.S.; Ibrahim, R.E. The alleviative effects of green synthesized copper oxide nanoparticles against oxidative stress, hepato-renal alterations, and immune suppression induced by Staphylococcus aureus infection in Clarias gariepinus. Aquac. Rep. 2023, 32, 101746. [Google Scholar] [CrossRef]
- Awad, M.; Yosri, M.; Abdel-Aziz, M.M.; Younis, A.M.; Sidkey, N.M. Assessment of the antibacterial potential of biosynthesized silver nanoparticles combined with vancomycin against methicillin-resistant Staphylococcus aureus–induced infection in rats. Biol. Trace Elem. Res. 2021, 199, 4225–4236. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.M.; El-Kabany, H. Evaluation of protective and treatment of Thyme (Thymus vulgaris) oil on Toxocara vitulorum infected rats. J. Rad. Res. Appl. Sci. 2013, 6, 209–232. [Google Scholar]
- Mauer, S.M.; Steffes, M.W.; Brown, D.M. The kidney in diabetes. Am. J. Med. 1981, 70, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Baraaj, A.H. Histopathological and functional changes in kidney of male rabbits infected with Pseudomonas aeruginosa DNA. Iraqi J. Biotechnol. 2015, 14, 404–413. [Google Scholar]
- Cheng, A.G.; Kim, H.K.; Burts, M.L.; Krausz, T.; Schneewind, O.; Missiakas, D.M. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 2009, 23, 3393. [Google Scholar] [CrossRef] [PubMed]
- Rahmawati, J.; Da’i, M.; Wahyuni, A.S.; Sujono, T.A. The effect of ethanol extract of thyme (Thymus vulgaris L.) on level of urea and creatinine of serum neprhropathy diabetic rat. EurAsian J. Biosci. 2020, 14, 5341. [Google Scholar]
- Aref, A.B.M.; Momenah, M.A.; Jad, M.M.; Semmler, M.; Mohamedaiin, H.S.; Ahmed, A.; Mohamedien, D. Tramadol Biological Effects: 4: Effective therapeutic efficacy of Lagenaria siceraria preparation (Gamal & Aref1) and melatonin on cell biological, histochemical, and histopathological changes in the kidney of tramadol-induced male mice. Microsc. Microanal. 2021, 27, 613–625. [Google Scholar]
- Nadia, Z.; Rachid, M. Antioxidant and antibacterial activities of Thymus vulgaris L. Med. Aromat. Plant Res. J. 2013, 1, 5–11. [Google Scholar]
- Attar, U.A.; Ghane, S.G. In vitro antioxidant, antidiabetic, antiacetylcholine esterase, anticancer activities and RP-HPLC analysis of phenolics from the wild bottle gourd (Lagenaria siceraria (Molina) Standl. S. Afr. J. Bot. 2019, 125, 360–370. [Google Scholar] [CrossRef]
- Rostami, R.; Eslamifar, Z.; Nazemi, S.; Hosseini, S.Z.; Jafaripour, L. The effect of thyme essential oil on liver injuries caused by renal ischemia-reperfusion in rats. Res. Int. 2022, 2022, 2988334. [Google Scholar] [CrossRef]
- Boxx, G.M.; Cheng, G. The roles of type I interferon in bacterial infection. Cell Host Microbe 2016, 19, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Shtrichman, R.; Samuel, C.E. The role of gamma interferon in antimicrobial immunity. Curr. Opin. Microbiol. 2001, 4, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Roderiquez, G.; Norcross, M.A. Control of Adaptive Immune Responses by Staphylococcus aureus through IL-10, PD-L1 and TLR2. Sci. Rep. 2012, 2, 606. [Google Scholar] [CrossRef] [PubMed]
- Van Belleghem, J.D.; Clement, F.; Merabishvili, M.; Lavigne, R.; Vaneechoutte, M. Pro-and anti-inflammatory responses of peripheral blood mononuclear cells induced by Staphylococcus aureus and Pseudomonas aeruginosa phages. Sci. Rep. 2017, 7, 8004. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.; Blount, D.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Leech, J.M.; Lacey, K.A.; Mulcahy, M.E.; Medina, E.; McLoughlin, R.M. IL-10 plays opposing roles during Staphylococcus aureus systemic and localized infections. J. Immunol. 2017, 198, 2352–2365. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Class | AMA | No. of Resistant Strains (%) | MAR Index | p-Value |
|---|---|---|---|---|
| P. aeruginosa strains (n = 10) | ||||
| Aminoglycosides | CN | 7 (70.00) | 0.04 | 0.0736 |
| AK | 5 (50.00) | 0.03 | 1.00 | |
| NET | 6 (60.00) | 0.35 | 0.3710 | |
| TOB | 7 (70.00) | 0.04 | 0.0736 | |
| Carbapenems | IMP | 0 (0.00) | 0 | ND |
| MRP | 1 (10.00) | 0.005 | 0.0003 | |
| Cephalosporins | FOX | 10 (100.0) | 0.06 | ND |
| CAZ | 10 (100.0) | 0.06 | ND | |
| FEB | 5 (50.00) | 0.03 | 1.00 | |
| Quinolones | NA | 8 (80.00) | 0.05 | 0.0072 |
| CIP | 9 (90.00) | 0.05 | 0.0003 | |
| LEV | 8 (80.00) | 0.05 | 0.0072 | |
| Penicillin | PTZ | 1 (10.00) | 0.005 | 0.0003 |
| Sulphonamide | SXT | 10 (100.0) | 0.06 | ND |
| Monobactam | ATM | 7 (70.00) | 0.04 | 0.0736 |
| Phosphonic | FF | 10 (100.0) | 0.06 | ND |
| polymyxin | PB | 7 (70.00) | 0.04 | 0.0736 |
| VRSA strains (n = 10) | ||||
| Aminoglycosides | CN | 3 (30.00) | 0.02 | 0.0736 |
| S | 3 (30.00) | 0.02 | 0.0736 | |
| Rifamycin | RA | 2 (20.00) | 0.01 | 0.0072 |
| Cephalosporins | FOX | 10 (100.0) | 0.06 | ND |
| Penicillin | AX | 10 (100.0) | 0.06 | ND |
| AMP | 10 (100.0) | 0.06 | ND | |
| Glycopeptides | VA | 8 (80.00) | 0.05 | 0.0072 |
| Fluoroquinolones | CIP | 3 (30.00) | 0.02 | 0.0736 |
| Tetracyclines | TC | 4 (40.00) | 0.03 | 0.3710 |
| DO | 3 (30.00) | 0.02 | 0.0736 | |
| Sulphonamides | SXT | 7 (70.00) | 0.04 | 0.0736 |
| Carbapenems | IMP | 0 (0.00) | 0 | ND |
| Macrolide | E | 10 (100.0) | 0.06 | ND |
| Chloramphenicol | C | 10 (100.0 | 0.06 | ND |
| Phosphonic | FF | 10 (100.0) | 0.06 | ND |
| Fusidane | FA | 9 (90.00) | 0.05 | 0.0003 |
| Peak | Compound | Formula | Retention Time (min) | Area under Peak | % |
|---|---|---|---|---|---|
| 1 | Hexadecanoic acid, methyl ester | C17H34O2 | 17.25 | 2,340,683.91 | 4.93 |
| 2 | 9,12-Octadecadienoic acid (Z,Z)-, methyl ester | C19H34O2 | 19.111 | 39,671,581.83 | 83.5 |
| 3 | 9-Octadecenoic acid (Z)-, methyl ester | C19H36O2 | 19.154 | 4,782,642.89 | 10.07 |
| 4 | Octadecanoic acid, methyl ester | C19H38O2 | 19.362 | 715,454.13 | 1.51 |
| Peak | Compound | Formula | Retention Time (min) | Area under Peak | % |
|---|---|---|---|---|---|
| 1 | 3-Carene | C10H16 | 4.43 | 714,943.46 | 7.45 |
| 2 | Benzene, 1-methyl-3-(1-methylethyl)- | C10H14 | 5.86 | 1,292,251.84 | 13.46 |
| 3 | D-Limonene | C10H16 | 5.917 | 2,559,940.08 | 26.67 |
| 4 | Eucalyptol | C10H18O | 6.038 | 437,115.8 | 4.55 |
| 5 | Gamma-Terpinene | C10H16 | 6.375 | 272,565.1 | 2.84 |
| 6 | Bicyclo[2.2.1]heptan-2-one, 1,7,7-trimethyl-, (1S)- | C10H16O | 7.817 | 594,544.71 | 6.19 |
| 7 | Terpinen-4-ol | C10H18O | 8.286 | 1,028,966.54 | 10.72 |
| 8 | Alpha-Terpineol | C10H18O | 8.509 | 287,248.53 | 2.99 |
| 9 | N-Methyl-1-adamantaneacetamide | C13H21NO | 9.242 | 160,158.78 | 1.67 |
| 10 | Isobornyl acetate | C12H20O2 | 9.785 | 1,156,405.17 | 12.05 |
| 11 | Alpha-Terpinyl acetate | C12H20O2 | 10.581 | 933,090.12 | 9.72 |
| 12 | Di-n-decylsulfone | C20H42O2S | 12.578 | 161,638.51 | 1.68 |
| Bacterial Strains | Code No. | Lagenaria siceraria | L. siceraria + Chitosan NPs | Thymus vulgaris | T. vulgaris + Chitosan NPs | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zone Diameters (mm) | Broth Microdilution (µg/mL) | Zone Diameters (mm) | Broth Microdilution (µg/mL) | Zone Diameters (mm) | Broth Microdilution (µg/mL) | Zone Diameters (mm) | Broth Microdilution (µg/mL) | ||||||||||||||
| P. aeruginosa | 20% | 60% | 100% | MIC | MBC | 20% | 60% | 100% | MIC | MBC | 20% | 60% | 100% | MIC | MBC | 20% | 60% | 100% | MIC | MBC | |
| 1P | 0 | 2 | 15 | 128 | 256 | 30 | 40 | 50 | 0.5 | 1 | 0 | 6 | 14 | 128 | 256 | 20 | 30 | 35 | 4 | 8 | |
| 2P | 0 | 0 | 10 | 128 | 256 | 28 | 38 | 45 | 0.5 | 1 | 0 | 4 | 14 | 128 | 256 | 25 | 30 | 40 | 4 | 8 | |
| 3P | 0 | 6 | 18 | 64 | 128 | 32 | 40 | 50 | 0.5 | 1 | 0 | 5 | 15 | 64 | 64 | 10 | 17 | 25 | 2 | 4 | |
| 4P | 0 | 4 | 16 | 64 | 128 | 30 | 42 | 50 | 1 | 2 | 0 | 5 | 16 | 64 | 128 | 20 | 35 | 42 | 8 | 16 | |
| 5P | 0 | 8 | 20 | 128 | 256 | 32 | 44 | 48 | 1 | 2 | 0 | 8 | 18 | 64 | 128 | 20 | 30 | 40 | 4 | 8 | |
| 6P | 0 | 2 | 12 | 64 | 128 | 24 | 32 | 44 | 0.5 | 1 | 0 | 6 | 16 | 32 | 64 | 15 | 22 | 30 | 4 | 8 | |
| 7P | 0 | 4 | 14 | 64 | 128 | 30 | 35 | 45 | 0.25 | 0.5 | 0 | 6 | 15 | 128 | 256 | 20 | 24 | 32 | 4 | 8 | |
| 8P | 0 | 4 | 14 | 128 | 256 | 28 | 34 | 44 | 1 | 2 | 0 | 5 | 12 | 64 | 128 | 24 | 32 | 40 | 4 | 8 | |
| 9P | 0 | 0 | 6 | 64 | 128 | 25 | 38 | 46 | 0.5 | 1 | 0 | 0 | 10 | 64 | 128 | 25 | 30 | 38 | 8 | 16 | |
| 10P | 0 | 2 | 6 | 128 | 256 | 32 | 40 | 50 | 1 | 2 | 0 | 4 | 16 | 128 | 256 | 25 | 32 | 40 | 8 | 16 | |
| VRSA | 1V | 0 | 2 | 15 | 128 | 256 | 30 | 40 | 50 | 0.25 | 0.5 | 0 | 4 | 12 | 32 | 64 | 20 | 32 | 42 | 2 | 4 |
| 2V | 0 | 0 | 10 | 128 | 256 | 28 | 38 | 45 | 0.25 | 0.5 | 0 | 0 | 10 | 64 | 128 | 18 | 30 | 40 | 2 | 4 | |
| 3V | 0 | 6 | 18 | 128 | 256 | 32 | 40 | 50 | 0.125 | 0.25 | 0 | 6 | 12 | 128 | 256 | 18 | 30 | 36 | 1 | 2 | |
| 4V | 0 | 4 | 16 | 32 | 64 | 30 | 42 | 50 | 0.25 | 0.5 | 0 | 6 | 14 | 16 | 32 | 22 | 32 | 38 | 2 | 4 | |
| 5V | 0 | 8 | 20 | 64 | 128 | 32 | 44 | 48 | 0.25 | 0.5 | 0 | 8 | 20 | 64 | 128 | 25 | 35 | 45 | 2 | 4 | |
| 6V | 0 | 2 | 12 | 128 | 256 | 24 | 32 | 44 | 0.5 | 1 | 0 | 6 | 16 | 128 | 256 | 20 | 28 | 32 | 2 | 4 | |
| 7V | 0 | 4 | 14 | 64 | 128 | 30 | 35 | 45 | 0.5 | 1 | 0 | 5 | 14 | 64 | 128 | 18 | 30 | 38 | 4 | 8 | |
| 8V | 0 | 4 | 14 | 64 | 128 | 28 | 34 | 44 | 0.125 | 0.25 | 0 | 4 | 12 | 64 | 128 | 22 | 30 | 40 | 2 | 4 | |
| 9V | 0 | 0 | 6 | 128 | 256 | 25 | 38 | 46 | 0.25 | 0.5 | 0 | 5 | 10 | 64 | 128 | 14 | 26 | 32 | 2 | 4 | |
| 10V | 0 | 2 | 6 | 64 | 128 | 32 | 40 | 50 | 0.5 | 1 | 0 | 6 | 12 | 64 | 128 | 16 | 28 | 38 | 4 | 8 | |
| Groups | ALT (IU/dL) | AST (IU/dL) | Alb (g/dL) | Creatinine (mg/dL) | Urea (mg/dL) | SOD (U/mL) | GPX (MU/mL) | MDA (mmol/mL) |
|---|---|---|---|---|---|---|---|---|
| G1 | 42.97 ± 1.36 b | 121.33 ± 2.96 b | 3.52 ± 0.03 a | 0.57 ± 0.01 b | 24.13 ± 5.52 a,b | 126.33 ± 0.88 a | 19.26 ± 0.57 a | 3.27 ± 0.23 f |
| G2 | 58.60 ± 0.56 a | 155.33 ± 2.60 a | 3.07 ± 0.04 c | 0.66 ± 0.02 a | 25.80 ± 6.31 a,b | 62.67 ± 1.45 e | 2.62 ± 0.33 e | 14.77 ± 0.30 a |
| G3 | 58.97 ± 1.03 a | 146.67 ± 2.03 a | 3.27 ± 0.04 b | 0.64 ± 0.03 a | 14.39 ± 1.86 b | 68.00 ± 2.31 d | 5.63 ± 0.75 d | 12.03 ± 0.43 b |
| G4 | 40.43 ± 2.78 b | 132.33 ± 5.36 b | 3.35 ± 0.03 b | 0.68 ± 0.01 a | 19.18 ± 2.21 a,b | 97.67 ± 1.20 c | 9.24 ± 0.61 c | 10.87 ± 0.30 c |
| G5 | 41.70 ± 3.65 b | 127.67 ± 4.06 b | 3.57 ± 0.04 a | 0.52 ± 0.02 b | 15.25 ± 1.93 b | 103.67 ± 0.84 b | 12.80 ± 0.64 b | 7.13 ± 0.48 e |
| G6 | 42.23 ± 1.63 b | 130.67 ± 3.28 b | 3.32 ± 0.06 b | 0.66 ± 1.01 a | 36.03 ± 4.72 a | 97.33 ± 1.20 c | 9.42 ± 0.49 c | 9.63 ± 0.23 d |
| G7 | 60.40 ± 1.72 A | 148.67 ± 3.76 A | 3.31 ± 0.05 B,C | 0.65 ± 0.01 A | 18.99 ± 0.73 | 69.33 ± 2.33 F | 4.77 ± 0.24 E | 15.87 ± 0.38 A |
| G8 | 54.53 ± 0.88 A | 148.67 ± 4.10 A | 3.48 ± 0.06 A,B | 0.65 ± 0.01 A | 24.43 ± 2.69 | 78.00 ± 1.15 E | 7.97 ± 0.41 D | 13.13 ± 0.48 B |
| G9 | 56.73 ± 2.70 A | 143.33 ± 5.24 A | 3.23 ± 0.10 C | 0.61 ± 0.03 A,B | 31.80 ± 7.68 | 93.67 ± 0.88 D | 10.40 ± 0.29 C | 11.20 ± 0.46 C |
| G10 | 40.00 ± 2.08 B | 116.00 ± 3.79 B | 3.48 ± 0.02 A,B | 0.63 ± 0.01 A,B | 24.78 ± 1.56 | 108.00 ± 2.08 B | 12.83 ± 0.91 B | 7.37 ± 0.61 E |
| G11 | 41.30 ± 1.42 B | 123.67 ± 2.73 B | 3.35 ± 0.03 A,B,C | 0.63 ± 0.04 A,B | 20.35 ± 2.32 | 98.33 ± 0.81 C | 12.10 ± 0.97 B,C | 9.20 ± 0.35 D |
| p-value | 0.0005 | <0.0001 | 0.0189 | 0.0004 | 0.0500 | <0.0001 | <0.0001 | <0.0001 |
| Groups | Hb (g/dL) | RBCs (106/mm3) | WBCs (109/L) | NEUT (%) | LYM (%) |
|---|---|---|---|---|---|
| G1 | 11.77 ± 0.02 a | 6.87 ± 0.02 a | 9.67 ± 0.09 e | 17.33 ± 1.20 e | 83.33 ± 1.20 a |
| G2 | 9.35 ± 0.09 c | 5.37 ± 0.08 c | 23.17 ± 0.59 a | 39.33 ± 1.19 a | 62.33 ± 1.20 d |
| G3 | 9.58 ± 0.04 c | 5.84 ± 0.02 b | 18.23 ± 0.19 b | 33.00 ± 1.15 b | 72.67 ± 2.33 bc |
| G4 | 10.18 ± 0.12 b | 5.89 ± 0.10 b | 15.37 ± 0.29 c | 28.33 ± 1.21 c | 72.67 ± 0.88 bc |
| G5 | 11.60 ± 0.14 a | 6.68 ± 0.08 a | 12.60 ± 0.74 d | 22.33 ± 0.87 d | 76.33 ± 1.45 b |
| G6 | 10.41 ± 0.41 b | 6.08 ± 0.11 b | 14.87 ± 0.18 c | 29.00 ± 0.12 c | 69.33 ± 0.87 c |
| G7 | 9.41 ± 0.06 C | 5.41 ± 0.02 E | 18.37 ± 0.50 A | 33.00 ± 0.58 A | 68.33 ± 0.84 D |
| G8 | 10.67 ± 0.22 B | 6.26 ± 0.06 C | 13.80 ± 0.38 BC | 25.67 ± 0.67 B | 73.33 ± 0.82 C |
| G9 | 9.63 ± 0.11 C | 5.61 ± 0.06 D | 14.83 ± 0.35 B | 28.33 ± 0.88 B | 71.00 ± 0.58 C |
| G10 | 11.64 ± 0.09 A | 6.47 ± 0.03 B | 9.87 ± 0.34 D | 21.67 ± 0.74 C | 76.67 ± 0.80 B |
| G11 | 10.57 ± 0.26 B | 6.14 ± 0.12 C | 12.87 ± 0.35 C | 26.00 ± 0.58 B | 73.00 ± 0.57 C |
| p-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taha, S.M.; Abd El-Aziz, N.K.; Abdelkhalek, A.; Pet, I.; Ahmadi, M.; El-Nabtity, S.M. Chitosan-Loaded Lagenaria siceraria and Thymus vulgaris Potentiate Antibacterial, Antioxidant, and Immunomodulatory Activities against Extensive Drug-Resistant Pseudomonas aeruginosa and Vancomycin-Resistant Staphylococcus aureus: In Vitro and In Vivo Approaches. Antioxidants 2024, 13, 428. https://doi.org/10.3390/antiox13040428
Taha SM, Abd El-Aziz NK, Abdelkhalek A, Pet I, Ahmadi M, El-Nabtity SM. Chitosan-Loaded Lagenaria siceraria and Thymus vulgaris Potentiate Antibacterial, Antioxidant, and Immunomodulatory Activities against Extensive Drug-Resistant Pseudomonas aeruginosa and Vancomycin-Resistant Staphylococcus aureus: In Vitro and In Vivo Approaches. Antioxidants. 2024; 13(4):428. https://doi.org/10.3390/antiox13040428
Chicago/Turabian StyleTaha, Selwan M., Norhan K. Abd El-Aziz, Adel Abdelkhalek, Ioan Pet, Mirela Ahmadi, and Sameh M. El-Nabtity. 2024. "Chitosan-Loaded Lagenaria siceraria and Thymus vulgaris Potentiate Antibacterial, Antioxidant, and Immunomodulatory Activities against Extensive Drug-Resistant Pseudomonas aeruginosa and Vancomycin-Resistant Staphylococcus aureus: In Vitro and In Vivo Approaches" Antioxidants 13, no. 4: 428. https://doi.org/10.3390/antiox13040428