

Phytoconstituents of Androstachys johnsonii Prain Prevent Reactive Oxygen Species Production and Regulate the Expression of Inflammatory Mediators in LPS-Stimulated RAW 264.7 Macrophages

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Extraction Process
2.2. Antioxidant Assays
2.2.1. ABTS•+ Radical Scavenging Assay
2.2.2. DPPH• Radical Scavenging Assay
2.2.3. Ferric Reducing Antioxidant Power (FRAP) Assay
2.3. Anti-Inflammatory Assays
2.3.1. Cell Culture of Murine RAW 264.7 Macrophages and Cell Viability Assay
2.3.2. Nitric Oxide Production Inhibitory Assay
2.3.3. Quantification of Cyclo-Oxygenase (COX-2) and Cytokine Expression in LPS-Stimulated RAW 264.7 Cells
2.3.4. Measurement of Intracellular Reactive Oxygen Species (ROS) Levels
2.3.5. Soybean 15-Lipoxygenase (15-LOX) Enzyme Inhibitory Assay
2.4. Phytochemical Analysis
2.4.1. Total Phenolic Content
2.4.2. Total Flavonoid Content
2.4.3. Liquid Chromatography-Mass Spectrometric (LC-MS) Profiling
2.5. Statistical Analysis
3. Results and Discussion
3.1. Antioxidant Activity of A. johnsonii Hydroethanolic Leaf Extract
3.2. Anti-Inflammatory Activity of A. johnsonii Hydroethanolic Leaf Extract (AJHLE)
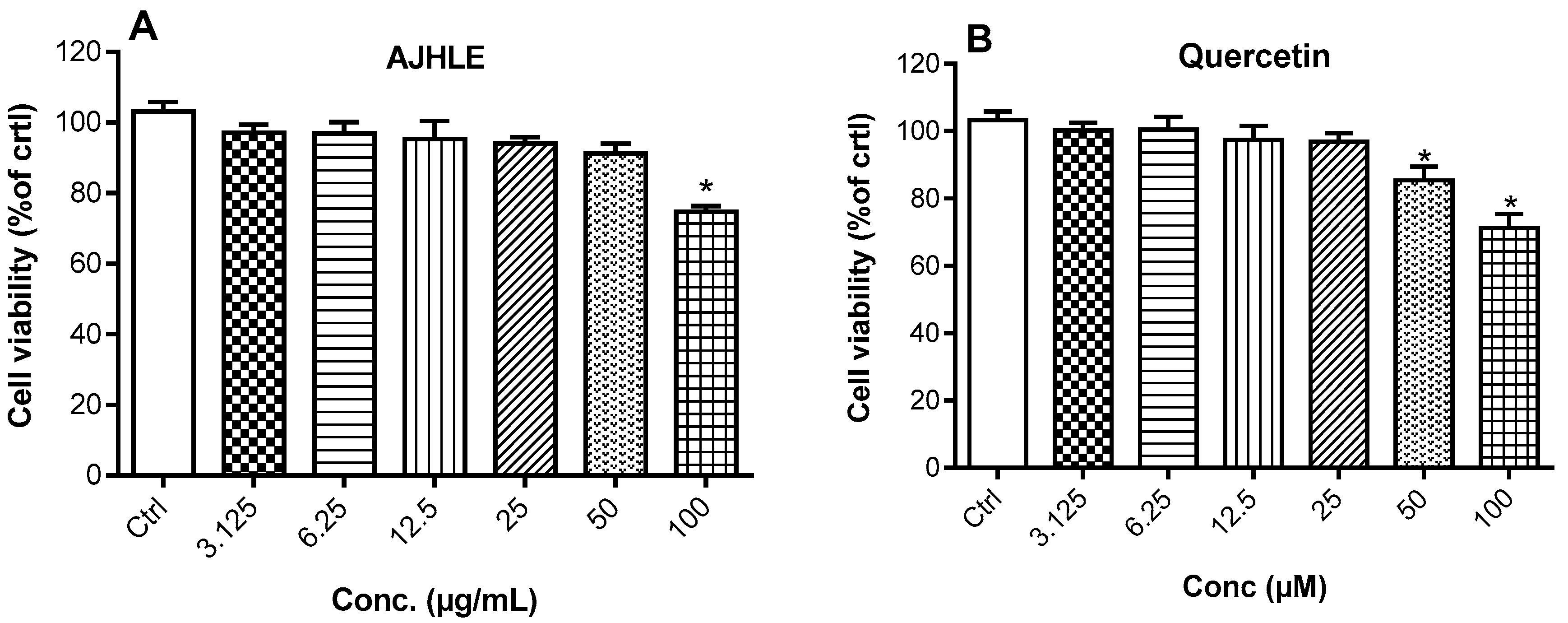
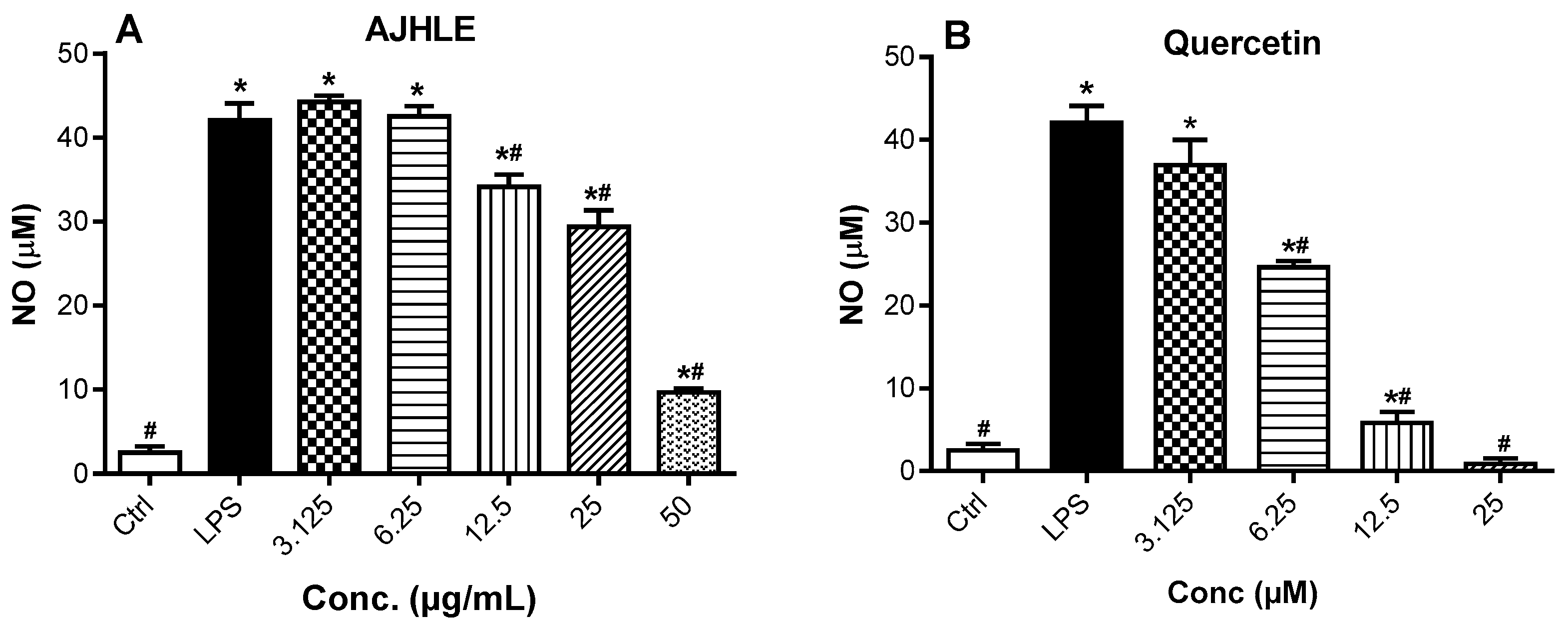
3.2.1. Cytotoxic and Nitric Oxide Inhibitory Effects of AJHLE on LPS-Stimulated RAW 264.7 Macrophages
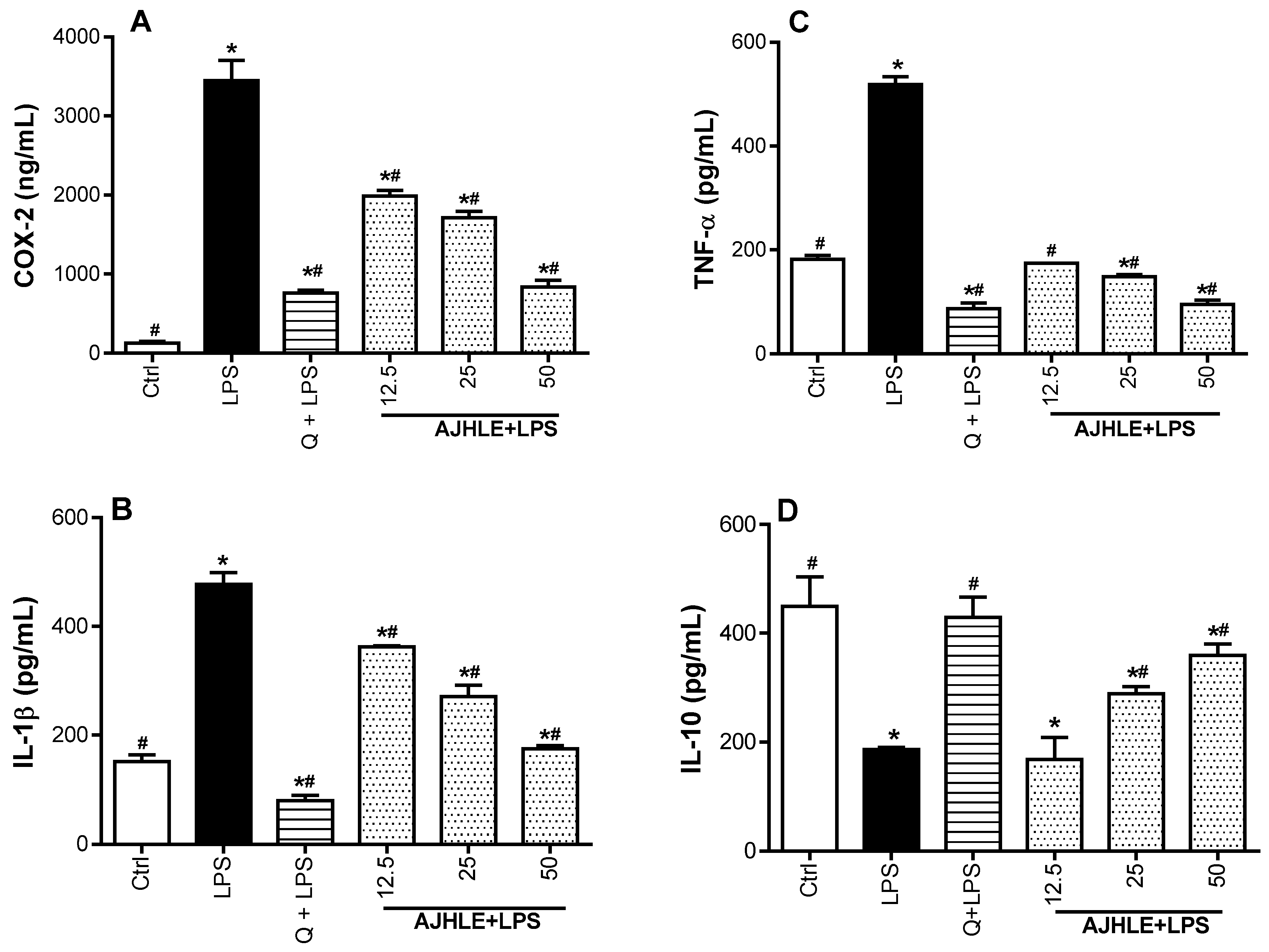
3.2.2. Regulatory Effect of AJHLE on the Expression of Pro-and Anti-Inflammatory Mediators
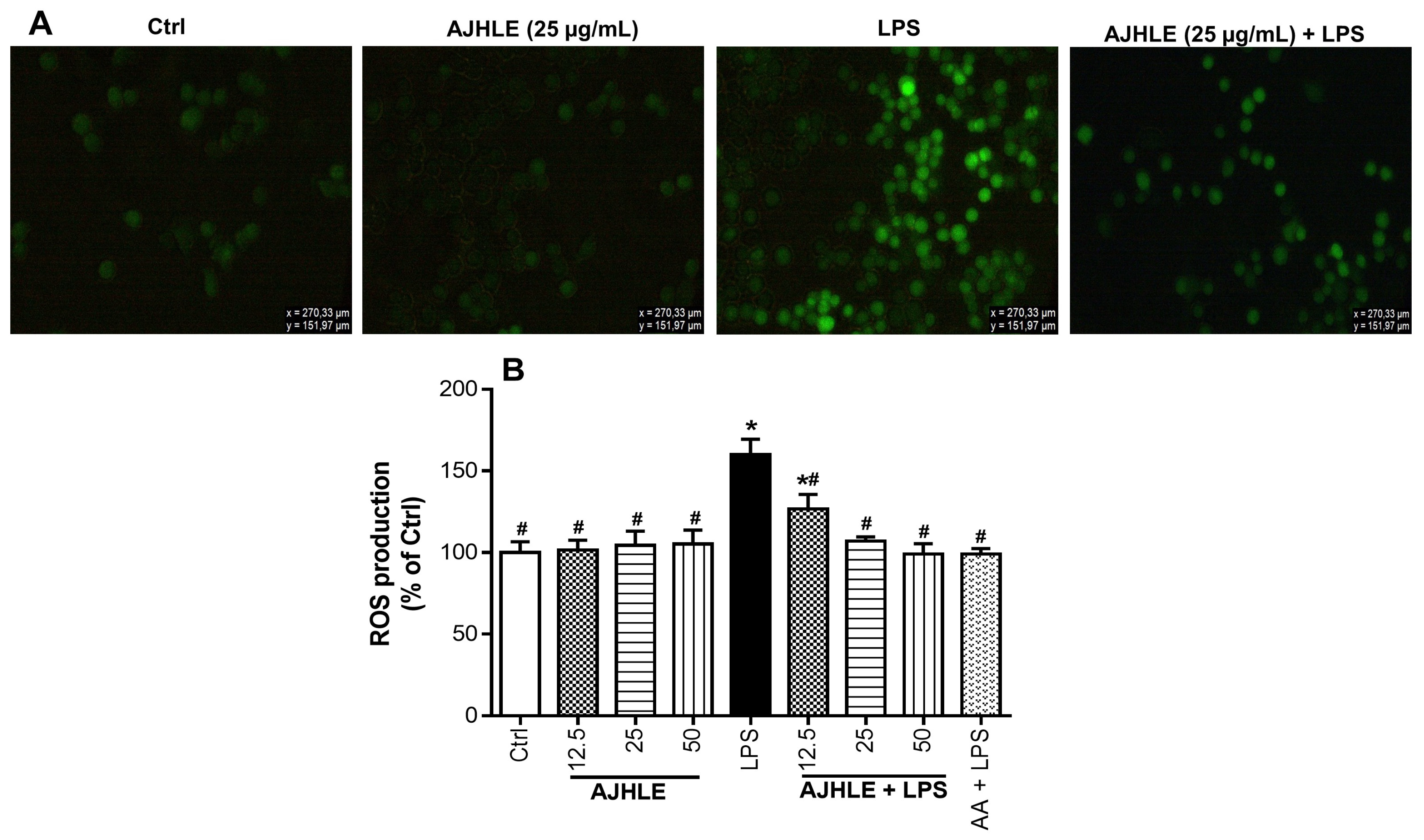
3.2.3. Protective Effect of AJHLE on Reactive Oxygen Species Production
3.2.4. 15-Lipoxygenase Inhibitory Activity of AJHLE
3.3. Phytochemical Composition of AJHLE
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Jouda, J.B.; Mfotie Njoya, E.; Mbazoa, C.D.; Zhou, Z.; Meli Lannang, A.; Wandji, J.; Shiono, Y.; Wang, F. Lambertellin from Pycnoporus sanguineus MUCL 51321 and its anti-inflammatory effect via modulation of MAPK and NF-kappaB signaling pathways. Bioorg. Chem. 2018, 80, 216–222. [Google Scholar] [CrossRef]
- Hunter, P. The inflammation theory of disease. The growing realization that chronic inflammation is crucial in many diseases opens new avenues for treatment. EMBO Rep. 2012, 13, 968–970. [Google Scholar] [CrossRef]
- Ackermann, J.A.; Hofheinz, K.; Zaiss, M.M.; Kronke, G. The double-edged role of 12/15-lipoxygenase during inflammation and immunity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 371–381. [Google Scholar] [CrossRef]
- Lu, X.J.; Ning, Y.J.; Liu, H.; Nie, L.; Chen, J. A Novel Lipopolysaccharide Recognition Mechanism Mediated by Internalization in Teleost Macrophages. Front. Immunol. 2018, 9, 2758. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef] [PubMed]
- Ghlichloo, I.; Gerriets, V. Nonsteroidal Anti-Inflammatory Drugs (NSAIDs). In StatPearls; Disclosure: Valerie Gerriets declares no relevant financial relationships with ineligible companies; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Street, R.A.; Prinsloo, G. Commercially Important Medicinal Plants of South Africa: A Review. J. Chem. 2013, 2013, 205048. [Google Scholar] [CrossRef]
- Asigbaase, M.; Adusu, D.; Anaba, L.; Abugre, S.; Kang-Milung, S.; Acheamfour, S.A.; Adamu, I.; Ackah, D.K. Conservation and economic benefits of medicinal plants: Insights from forest-fringe communities of Southwestern Ghana. Trees For. People 2023, 14, 100462. [Google Scholar] [CrossRef]
- Mamathaba, M.P.; Yessoufou, K.; Moteetee, A. What Does It Take to Further Our Knowledge of Plant Diversity in the Megadiverse South Africa? Diversity 2022, 2022, 748. [Google Scholar] [CrossRef]
- Williams, V.L.; Victor, J.E.; Crouch, N.R. Red Listed medicinal plants of South Africa: Status, trends, and assessment challenges. S. Afr. J. Bot. 2013, 86, 23–35. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, S.; Drakou, E.G.; Hickler, T.; Thines, M.; Nogues-Bravo, D. Evaluating natural medicinal resources and their exposure to global change. Lancet Planet. Health 2023, 7, e155–e163. [Google Scholar] [CrossRef] [PubMed]
- Bakali, M.; Ligavha-Mbelengwa, M.H.; Potgieter, M.J.; Tshisikhawe, M.P. Impact of Ethnobotanical Utilization on the Population Structure of Androstachys Johnsonii Prain in the Vhembe Area of the Limpopo Province, South Africa. Insight For. Res. 2017, 1, 50–56. [Google Scholar] [CrossRef]
- Van Wyk, B.; Van Wyk, P. Field Guide to Trees of Southern Africa; Struik Publishers: Cape Town, South Africa, 1997. [Google Scholar]
- Lukhele, M.S.; Van Ryssen, J.B.J. The chemical composition and potential nutritive value of foliage of four subtropical tree species in South Africa for ruminants. S. Afr. J. Anim. Sci. 2003, 33, 132–141. [Google Scholar] [CrossRef]
- Samie, A.; Obi, C.L.; Bessong, P.O.; Namirata, L. Activity profiles of fourteen selected medicinal plants from Rural Venda communities in South Africa against fifteen clinical bacterial species. Afr. J. Biotechnol. 2005, 4, 1443–1451. [Google Scholar]
- Ajao, A.A.; Sibiya, N.P.; Moteetee, A.N. Sexual prowess from nature: A systematic review of medicinal plants used as aphrodisiacs and sexual dysfunction in sub-Saharan Africa. S. Afr. J. Bot. 2019, 122, 342–359. [Google Scholar] [CrossRef]
- Maroyi, A. Traditional use of medicinal plants in south-central Zimbabwe: Review and perspectives. J. Ethnobiol. Ethnomed. 2013, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Molotja, G.M.; Ligavha-Mbelengwa, M.H.; Bhat, R.B. Antifungal activity of root, bark, leaf and soil extracts of Androstachys johnsonii Prain. Afr. J. Biotechnol. 2011, 10, 5725–5727. [Google Scholar]
- Makhafola, T.J.; Elgorashi, E.E.; McGaw, L.J.; Verschaeve, L.; Eloff, J.N. The correlation between antimutagenic activity and total phenolic content of extracts of 31 plant species with high antioxidant activity. BMC Complement. Altern. Med. 2016, 16, 490. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Mfotie NJoya, E. Medicinal plants, antioxidant potential, and cancer. In Cancer: Oxidative Stress and Dietary Antioxidants, 2nd ed.; Academic Press: San Diego, CA, USA, 2021; pp. 349–357. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Mfotie Njoya, E.; Ndemangou, B.; Akinyelu, J.; Munvera, A.M.; Chukwuma, C.I.; Mkounga, P.; Mashele, S.S.; Makhafola, T.J.; McGaw, L.J. In vitro antiproliferative, anti-inflammatory effects and molecular docking studies of natural compounds isolated from Sarcocephalus pobeguinii (Hua ex Pobég). Front. Pharmacol. 2023, 14, 1205414. [Google Scholar] [CrossRef]
- Marrazzo, P.; Angeloni, C.; Freschi, M.; Lorenzini, A.; Prata, C.; Maraldi, T.; Hrelia, S. Combination of Epigallocatechin Gallate and Sulforaphane Counteracts In Vitro Oxidative Stress and Delays Stemness Loss of Amniotic Fluid Stem Cells. Oxidative Med. Cell. Longev. 2018, 2018, 5263985. [Google Scholar] [CrossRef]
- Pinto, M.C.; Tejeda, A.; Duque, A.L.; Macias, P. Determination of lipoxygenase activity in plant extracts using a modified ferrous oxidation-xylenol orange assay. J. Agric. Food Chem. 2007, 55, 5956–5959. [Google Scholar] [CrossRef]
- Singleton, V.; Orthofer, R.; Lamuela-Raventós, R. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.; Barrow, C. A simple 96-well microplate method for estimation of total polyphenol content in seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Papi, S.; Ahmadizar, F.; Hasanvand, A. The role of nitric oxide in inflammation and oxidative stress. Immunopathol. Persa 2019, 5, e08. [Google Scholar] [CrossRef]
- Desai, S.J.; Prickril, B.; Rasooly, A. Mechanisms of Phytonutrient Modulation of Cyclooxygenase-2 (COX-2) and Inflammation Related to Cancer. Nutr. Cancer 2018, 70, 350–375. [Google Scholar] [CrossRef]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, S.; Berberi, A.; Fayyad-Kazan, M. Cytokines, Masticatory Muscle Inflammation, and Pain: An Update. J. Mol. Neurosci. 2020, 70, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Aghababaei, F.; Hadidi, M. Recent Advances in Potential Health Benefits of Quercetin. Pharmaceuticals 2023, 16, 1020. [Google Scholar] [CrossRef]
- Anik, M.I.; Mahmud, N.; Masud, A.A.; Khan, M.I.; Islam, M.N.; Uddin, S.; Hossain, M.K. Role of Reactive Oxygen Species in Aging and Age-Related Diseases: A Review. ACS Appl. Bio Mater. 2022, 5, 4028–4054. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef]
- Gegotek, A.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants 2022, 11, 1993. [Google Scholar] [CrossRef]
- Singh, N.K.; Rao, G.N. Emerging role of 12/15-Lipoxygenase (ALOX15) in human pathologies. Prog. Lipid Res. 2019, 73, 28–45. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Diaz, P.; Jeong, S.C.; Lee, S.; Khoo, C.; Koyyalamudi, S.R. Antioxidant and anti-inflammatory activities of selected medicinal plants and fungi containing phenolic and flavonoid compounds. Chin. Med. 2012, 7, 26. [Google Scholar] [CrossRef]
- Boukhary, R.; Raafat, K.; Ghoneim, A.I.; Aboul-Ela, M.; El-Lakany, A. Anti-Inflammatory and Antioxidant Activities of Salvia fruticosa: An HPLC Determination of Phenolic Contents. Evid. Based Complement. Altern. Med. 2016, 2016, 7178105. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Muema, F.W.; Kimutai, F.; Chen, G.; Guo, M. Phenolic Compounds from Carissa spinarum Are Characterized by Their Antioxidant, Anti-Inflammatory and Hepatoprotective Activities. Antioxidants 2021, 10, 652. [Google Scholar] [CrossRef]
- Leyva-Soto, A.; Alejandra Chavez-Santoscoy, R.; Porras, O.; Hidalgo-Ledesma, M.; Serrano-Medina, A.; Alejandra Ramirez-Rodriguez, A.; Alejandra Castillo-Martinez, N. Epicatechin and quercetin exhibit in vitro antioxidant effect, improve biochemical parameters related to metabolic syndrome, and decrease cellular genotoxicity in humans. Food Res. Int. 2021, 142, 110101. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, M.; Lu, G.; Wang, R.; Wei, Y.; Guo, Y.; Yu, Y.; Jiang, C. Anti-inflammation of epicatechin mediated by TMEM35A and TMPO in bovine mammary epithelial cell line cells and mouse mammary gland. J. Dairy Sci. 2021, 104, 12925–12938. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Li, F.; Zhang, X.; Xu, W.; Wang, Y.; Yao, Y.; Han, Z.; Xia, D. (-)-Epicatechin Ameliorates Monosodium Urate-Induced Acute Gouty Arthritis Through Inhibiting NLRP3 Inflammasome and the NF-kappaB Signaling Pathway. Front. Pharmacol. 2022, 13, 799552. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Cremonini, E.; Kang, H.; Kang, J.; Haj, F.G.; Oteiza, P.I. Anti-inflammatory actions of (-)-epicatechin in the adipose tissue of obese mice. Int. J. Biochem. Cell Biol. 2016, 81, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; You, Q.; Hu, L.; Gao, J.; Meng, Q.; Liu, W.; Wu, X.; Xu, Q. The Antioxidant Procyanidin Reduces Reactive Oxygen Species Signaling in Macrophages and Ameliorates Experimental Colitis in Mice. Front. Immunol. 2017, 8, 1910. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, L.; Yuan, Y.; Luo, X.; Jiang, M.; Ni, J.; Wang, N. Procyanidin B2 inhibits NLRP3 inflammasome activation in human vascular endothelial cells. Biochem. Pharmacol. 2014, 92, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, Y.; Huang, J.; Li, Y.; Lin, Z.; Zhang, B. Rutin ameliorates gout via reducing XOD activity, inhibiting ROS production and NLRP3 inflammasome activation in quail. Biomed. Pharmacother. 2023, 158, 114175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Gao, H.; Yuan, R.; Han, S.; Li, X.X.; Tang, M.; Dong, B.; Li, J.X.; Zhao, L.C.; Feng, J.; et al. Procyanidin A2, a polyphenolic compound, exerts anti-inflammatory and anti-oxidative activity in lipopolysaccharide-stimulated RAW264.7 cells. PLoS ONE 2020, 15, e0237017. [Google Scholar] [CrossRef]
- Bak, M.J.; Truong, V.L.; Kang, H.S.; Jun, M.; Jeong, W.S. Anti-inflammatory effect of procyanidins from wild grape (Vitis amurensis) seeds in LPS-induced RAW 264.7 cells. Oxid. Med. Cell. Longev. 2013, 2013, 409321. [Google Scholar] [CrossRef]
- Xu, H.-Y.; Feng, X.-H.; Zhao, P.-F.; Damirin, A.; Ma, C.-M. Procyanidin A2 penetrates L-02 cells and protects against tert-butyl hydroperoxide-induced oxidative stress by activating Nrf2 through JNK and p38 phosphorylation. J. Funct. Foods 2019, 62, 103562. [Google Scholar] [CrossRef]
- Muvhulawa, N.; Dludla, P.V.; Ziqubu, K.; Mthembu, S.X.H.; Mthiyane, F.; Nkambule, B.B.; Mazibuko-Mbeje, S.E. Rutin ameliorates inflammation and improves metabolic function: A comprehensive analysis of scientific literature. Pharmacol. Res. 2022, 178, 106163. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Yang, M.; Liu, M. Rutin prevents inflammation induced by lipopolysaccharide in RAW 264.7 cells via conquering the TLR4-MyD88-TRAF6-NF-kappaB signalling pathway. J. Pharm. Pharmacol. 2021, 73, 110–117. [Google Scholar] [CrossRef]
- Vo, Q.V.; Bay, M.V.; Nam, P.C.; Quang, D.T.; Flavel, M.; Hoa, N.T.; Mechler, A. Theoretical and Experimental Studies of the Antioxidant and Antinitrosant Activity of Syringic Acid. J. Org. Chem. 2020, 85, 15514–15520. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, D.M.; Sanches, M.P.; Poffo, C.M.; Parize, A.L.; Darelli, G.J.S.; de Lima, V.R. Syringic and cinnamic acids antiradical/antioxidant activities as R. ferruginea extract components and membrane physico-chemical influence. J. Mol. Struct. 2020, 1220, 128749. [Google Scholar] [CrossRef]
- Xiang, S.; Xiao, J. Protective effects of syringic acid on inflammation, apoptosis and intestinal barrier function in Caco-2 cells following oxygen-glucose deprivation/reoxygenation-induced injury. Exp. Ther. Med. 2022, 23, 66. [Google Scholar] [CrossRef] [PubMed]
- Somade, O.T.; Oyinloye, B.E.; Ajiboye, B.O.; Osukoya, O.A. Syringic acid demonstrates an anti-inflammatory effect via modulation of the NF-kappaB-iNOS-COX-2 and JAK-STAT signaling pathways in methyl cellosolve-induced hepato-testicular inflammation in rats. Biochem. Biophys. Rep. 2023, 34, 101484. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield of Extraction (g Extract/100 g Dry Material) | Phenolic Content (mgGAE/g Extract) | Flavonoid Content (mgQE/g Extract) | |
|---|---|---|---|
| AJHLE | 16.23 | 161.27 ± 0.90 | 33.57 ± 1.40 |
| IC50 (µg/mL) | |||
|---|---|---|---|
| Sample | ABTS | DPPH | FRAP |
| AJHLE | 9.07 ± 1.35 | 8.53 ± 1.16 | 79.09 ± 1.16 |
| Ascorbic acid | 4.99 ± 0.20 * | 4.64 ± 0.85 * | 26.68 ± 1.34 * |
| Quercetin | 2.94 ± 0.46 * | 3.21 ± 0.19 * | 21.62 ± 1.15 * |
| IC50 (µg/mL) | |||
|---|---|---|---|
| Sample | NO | 15-LOX | ROS |
| AJHLE | 24.06 ± 1.34 | 37.71 ± 1.03 | 10.49 ± 1.19 |
| Gallic acid | - | 22.08 ± 1.96 * | - |
| Quercetin (µM) | 7.08 ± 1.01 | - | - |
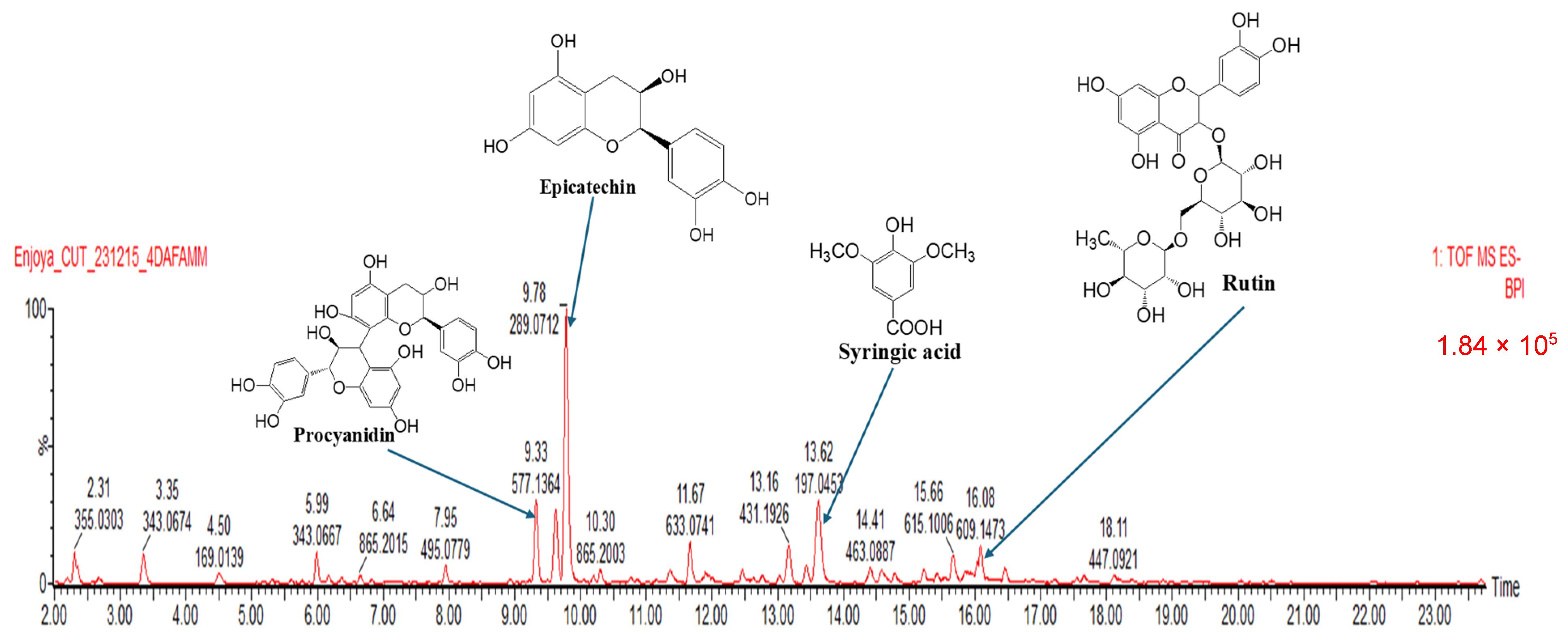
| Peak No | RT (min) | [M-H]-(m/z) | Tentative Assignment (Compound Name) | Ontology | Molecular Formula | Total Score | Peak Height Intensity | Conc. in Extract vs. EGCG (mg/L) |
|---|---|---|---|---|---|---|---|---|
| 1 | 5.99 | 343.06 | Theogallin | Quinic acids and derivatives | C14H16O10 | 7.27 | 16,943 | 626 |
| 2 | 6.64 | 865.19 | Procyanidin C1 | Biflavonoids and polyflavonoids | C45H38O18 | 7.49 | 6920 | 256 |
| 3 | 7.95 | 495.07 | Hibiscitrin | Flavonoid-3-O-glycosides | C21H20O14 | 7.10 | 10,731 | 397 |
| 4 | 9.33 | 577.13 | Procyanidin | Biflavonoids and polyflavonoids | C30H26O12 | 6.12 | 50,504 | 1867 |
| 5 | 9.73 | 311.04 | Caftaric acid | Coumaric acids and derivatives | C13H12O9 | 4.57 | 3,979 | 147 |
| 6 | 9.78 | 289.07 | Epicatechin | Catechins | C15H14O6 | 6.79 | 167,515 | 6193 |
| 7 | 11.36 | 541.05 | Putranjivain A | Hydrolyzable tannins | C46H36O31 | 4.38 | 8535 | 316 |
| 8 | 11.67 | 633.07 | Corilagin | Hydrolyzable tannins | C27H22O18 | 7.19 | 24,505 | 906 |
| 9 | 12.46 | 729.14 | ent-epicatechin 3-gallate | Biflavonoids and polyflavonoids | C37H30O16 | 5.06 | 7375 | 273 |
| 10 | 12.77 | 631.09 | 8-C-Ascorbylepigallocatechin 3-gallate | Catechin gallates | C28H24O17 | 4.54 | 4531 | 168 |
| 11 | 13.03 | 479.08 | Myricetin 3-galactoside | Flavonoid-3-O-glycosides | C21H20O13 | 6.84 | 4306 | 159 |
| 12 | 13.43 | 577.13 | Procyanidin B5 | Biflavonoids and polyflavonoids | C30H26O12 | 7.19 | 11,537 | 427 |
| 13 | 13.62 | 197.04 | Syringic acid | Gallic acid and derivatives | C9H10O5 | 6.75 | 53,310 | 1971 |
| 14 | 14.41 | 463.08 | Quercetin 3-galactoside | Flavonoid-3-O-glycosides | C21H20O12 | 8.11 | 9361 | 346 |
| 15 | 14.51 | 625.14 | Quercetin 3-glucosyl-2-galactoside | Flavonoid-3-O-glycosides | C27H29O17 | 7.22 | 1743 | 64 |
| 16 | 14.58 | 953.09 | Chebulagic acid | Hydrolyzable tannins | C41H30O27 | 4.89 | 6614 | 245 |
| 17 | 14.78 | 481.09 | 3′-(2″-Galloylglucosyl)-phloroacetophenone | Phenolic glycosides | C21H22O13 | 5.46 | 4214 | 156 |
| 18 | 15.23 | 595.13 | Quercetin 3-lathyroside | Flavonoid-3-O-glycosides | C26H28O16 | 6.68 | 8662 | 320 |
| 19 | 15.47 | 581.22 | (7′R)-(+)-Lyoniresinol 9′-glucoside | Lignan glycosides | C28H38O13 | 6.25 | 755 | 28 |
| 20 | 15.66 | 615.10 | Quercetin 3-(2-galloylglucoside) | Flavonoid-3-O-glycosides | C28H24O16 | 5.09 | 6334 | 234 |
| 21 | 16.08 | 609.14 | Rutin | Flavonoid-3-O-glycosides | C27H30O16 | 8.09 | 31,331 | 1158 |
| 22 | 16.14 | 463.08 | Kaempferol 3-alpha-l-arabinopyranoside | Flavonoid-3-O-glycosides | C20H18O10 | 6.13 | 1380 | 51 |
| 23 | 16.88 | 447.09 | Quercitrin | Flavonoid-3-O-glycosides | C21H20O11 | 8.09 | 2532 | 94 |
| 24 | 17.55 | 505.20 | Icariside E4 | 2-arylbenzofuran flavonoids | C26H34O10 | 5.89 | 3333 | 123 |
| 25 | 17.59 | 515.12 | 3,5-Dicaffeoylquinic acid | Quinic acids and derivatives | C25H24O12 | 6.11 | 467 | 17 |
| 26 | 17.65 | 599.10 | Kaempferol 7-(6″-galloylglucoside) | Flavonoid-7-O-glycosides | C28H24O15 | 5.43 | 4681 | 173 |
| 27 | 17.72 | 593.15 | Astragalin 7-rhamnoside | Flavonoid-7-O-glycosides | C27H30O15 | 8.12 | 2021 | 75 |
| 28 | 18.11 | 447.09 | Astragalin | Flavonoid-7-O-glycosides | C21H20O11 | 8.09 | 4786 | 177 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mfotie Njoya, E.; Tabakam, G.T.; Chukwuma, C.I.; Mashele, S.S.; Makhafola, T.J. Phytoconstituents of Androstachys johnsonii Prain Prevent Reactive Oxygen Species Production and Regulate the Expression of Inflammatory Mediators in LPS-Stimulated RAW 264.7 Macrophages. Antioxidants 2024, 13, 401. https://doi.org/10.3390/antiox13040401
Mfotie Njoya E, Tabakam GT, Chukwuma CI, Mashele SS, Makhafola TJ. Phytoconstituents of Androstachys johnsonii Prain Prevent Reactive Oxygen Species Production and Regulate the Expression of Inflammatory Mediators in LPS-Stimulated RAW 264.7 Macrophages. Antioxidants. 2024; 13(4):401. https://doi.org/10.3390/antiox13040401
Chicago/Turabian StyleMfotie Njoya, Emmanuel, Gaetan T. Tabakam, Chika I. Chukwuma, Samson S. Mashele, and Tshepiso J. Makhafola. 2024. "Phytoconstituents of Androstachys johnsonii Prain Prevent Reactive Oxygen Species Production and Regulate the Expression of Inflammatory Mediators in LPS-Stimulated RAW 264.7 Macrophages" Antioxidants 13, no. 4: 401. https://doi.org/10.3390/antiox13040401