
Cysteine Is the Only Universally Affected and Disfavored Proteomic Amino Acid under Oxidative Conditions in Animals

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inclusion Criteria for the Analyzed Species and Proteins
2.2. Data Analysis
2.3. Transmembrane Domain Content Correction
2.4. Body Mass and Phylogenetic Corrections
2.5. Statistical Analysis
3. Results and Discussion
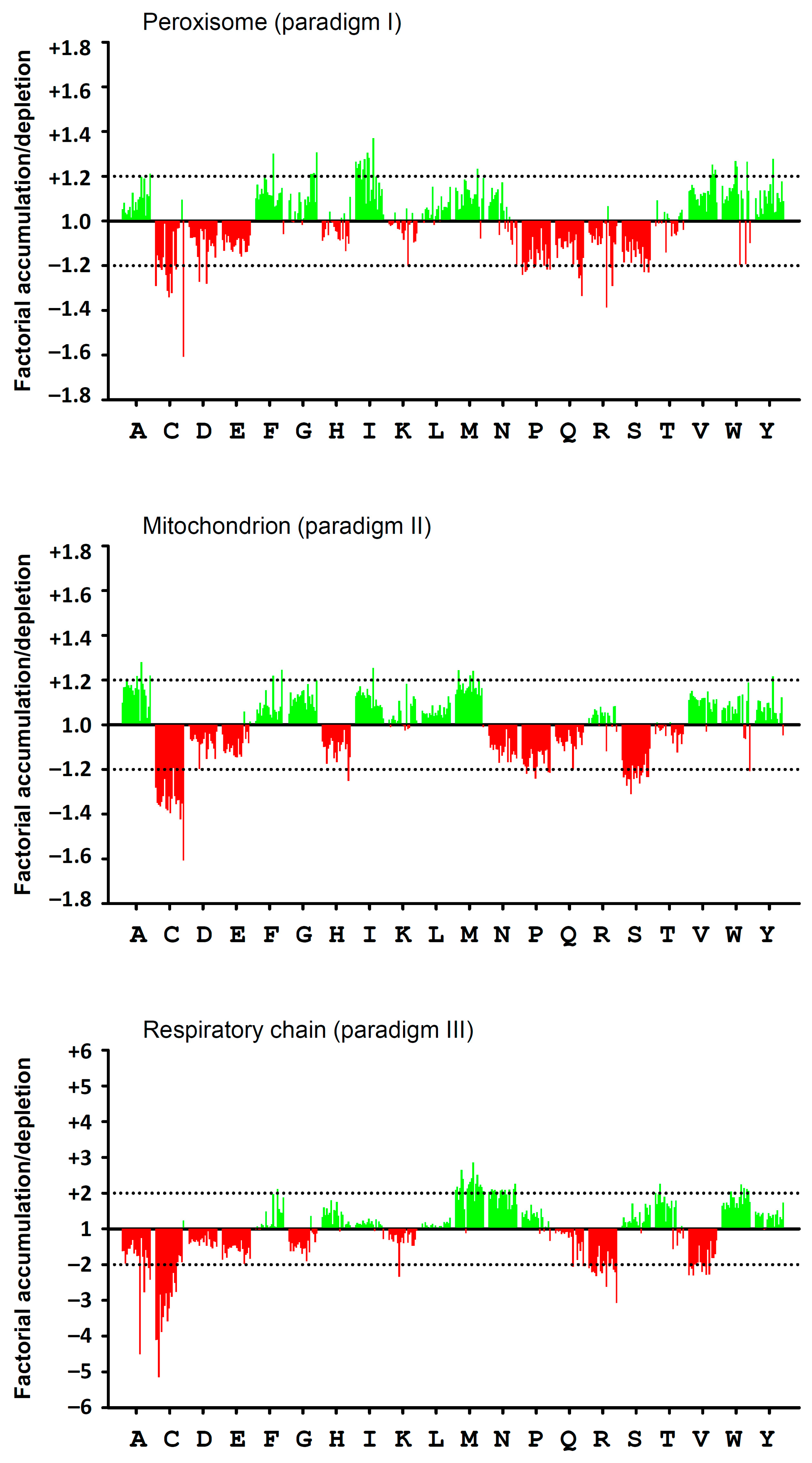
3.1. Amino Acid Usage Differences between Cellular Compartments
3.2. Amino Acid Usage Alterations in Relation to Longevity
3.3. Amino Acid Usage Adaptations Linked to an Aerobic Lifestyle
3.4. Relation of the Observed Amino Acid Usage Differences to Amino Acid Redox Reactivity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.; Barja, G. Highly resistant macromolecular components and low rate of generation of endogenous damage: Two key traits of longevity. Ageing Res. Rev. 2007, 6, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Golubev, A.; Hanson, A.D.; Gladyshev, V.N. A tale of two concepts: Harmonizing the free radical and antagonistic pleiotropy theories of aging. Antioxid. Redox Signal. 2018, 29, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed]
- Kunath, S.; Schindeldecker, M.; De Giacomo, A.; Meyer, T.; Sohre, S.; Hajieva, P.; von Schacky, C.; Urban, J.; Moosmann, B. Prooxidative chain transfer activity by thiol groups in biological systems. Redox Biol. 2020, 36, 101628. [Google Scholar] [CrossRef] [PubMed]
- Moosmann, B.; Hajieva, P. Probing the role of cysteine thiyl radicals in biology: Eminently dangerous, difficult to scavenge. Antioxidants 2022, 11, 885. [Google Scholar] [CrossRef] [PubMed]
- Moosmann, B.; Behl, C. Mitochondrially encoded cysteine predicts animal lifespan. Aging Cell 2008, 7, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Schindeldecker, M.; Stark, M.; Behl, C.; Moosmann, B. Differential cysteine depletion in respiratory chain complexes enables the distinction of longevity from aerobicity. Mech. Ageing Dev. 2011, 132, 171–179. [Google Scholar] [CrossRef]
- Kunath, S.; Moosmann, B. What is the rate-limiting step towards aging? Chemical reaction kinetics might reconcile contradictory observations in experimental aging research. Geroscience 2020, 42, 857–866. [Google Scholar] [CrossRef]
- Nauser, T.; Pelling, J.; Schöneich, C. Thiyl radical reaction with amino acid side chains: Rate constants for hydrogen transfer and relevance for posttranslational protein modification. Chem. Res. Toxicol. 2004, 17, 1323–1328. [Google Scholar] [CrossRef]
- Nauser, T.; Koppenol, W.H.; Schöneich, C. Protein thiyl radical reactions and product formation: A kinetic simulation. Free Radic. Biol. Med. 2015, 80, 158–163. [Google Scholar] [CrossRef]
- Hajieva, P.; Abrosimov, R.; Kunath, S.; Moosmann, B. Antioxidant and prooxidant modulation of lipid peroxidation by integral membrane proteins. Free Radic. Res. 2023, 57, 105–114. [Google Scholar] [CrossRef]
- Marino, S.M.; Gladyshev, V.N. Cysteine function governs its conservation and degeneration and restricts its utilization on protein surfaces. J. Mol. Biol. 2010, 404, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.M.; Gladyshev, V.N. Analysis and functional prediction of reactive cysteine residues. J. Biol. Chem. 2012, 287, 4419–4425. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Superoxide as an intracellular radical sink. Free Radic. Biol. Med. 1993, 14, 85–90. [Google Scholar] [CrossRef]
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef]
- Weissbach, H.; Etienne, F.; Hoshi, T.; Heinemann, S.H.; Lowther, W.T.; Matthews, B.; St John, G.; Nathan, C.; Brot, N. Peptide methionine sulfoxide reductase: Structure, mechanism of action, and biological function. Arch. Biochem. Biophys. 2002, 397, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Hajieva, P.; Moosmann, B. Adaptive antioxidant methionine accumulation in respiratory chain complexes explains the use of a deviant genetic code in mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 16496–16501. [Google Scholar] [CrossRef]
- Aledo, J.C. Methionine in proteins: The Cinderella of the proteinogenic amino acids. Protein Sci. 2019, 28, 1785–1796. [Google Scholar] [CrossRef]
- Aledo, J.C.; Li, Y.; de Magalhães, J.P.; Ruíz-Camacho, M.; Pérez-Claros, J.A. Mitochondrially encoded methionine is inversely related to longevity in mammals. Aging Cell 2011, 10, 198–207. [Google Scholar] [CrossRef]
- Schindeldecker, M.; Moosmann, B. Protein-borne methionine residues as structural antioxidants in mitochondria. Amino Acids 2015, 47, 1421–1432. [Google Scholar] [CrossRef]
- Moosmann, B. Redox biochemistry of the genetic code. Trends Biochem. Sci. 2021, 46, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.K.; Lalwani, N.D.; Reddy, J.K. Peroxisome proliferation and lipid peroxidation in rat liver. Cancer Res. 1986, 46, 1324–1330. [Google Scholar] [PubMed]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O. Role of peroxisomes in ROS/RNS-metabolism: Implications for human disease. Biochim. Biophys. Acta 2012, 1822, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Tahara, E.B.; Navarete, F.D.; Kowaltowski, A.J. Tissue-, substrate-, and site-specific characteristics of mitochondrial reactive oxygen species generation. Free Radic. Biol. Med. 2009, 46, 1283–1297. [Google Scholar] [CrossRef]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Goncalves, R.L.S.; Watson, M.A.; Wong, H.S.; Orr, A.L.; Brand, M.D. The use of site-specific suppressors to measure the relative contributions of different mitochondrial sites to skeletal muscle superoxide and hydrogen peroxide production. Redox Biol. 2020, 28, 101341. [Google Scholar] [CrossRef]
- Suthammarak, W.; Somerlot, B.H.; Opheim, E.; Sedensky, M.; Morgan, P.G. Novel interactions between mitochondrial superoxide dismutases and the electron transport chain. Aging Cell 2013, 12, 1132–1140. [Google Scholar] [CrossRef]
- Pamplona, R.; Costantini, D. Molecular and structural antioxidant defenses against oxidative stress in animals. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R843–R863. [Google Scholar] [CrossRef]
- Gomez, J.; Mota-Martorell, N.; Jove, M.; Pamplona, R.; Barja, G. Mitochondrial ROS production, oxidative stress and aging within and between species: Evidences and recent advances on this aging effector. Exp. Gerontol. 2023, 174, 112134. [Google Scholar] [CrossRef]
- Kitazoe, Y.; Kishino, H.; Hasegawa, M.; Matsui, A.; Lane, N.; Tanaka, M. Stability of mitochondrial membrane proteins in terrestrial vertebrates predicts aerobic capacity and longevity. Genome Biol. Evol. 2011, 3, 1233–1244. [Google Scholar] [CrossRef]
- Kitazoe, Y.; Hasegawa, M.; Tanaka, M.; Futami, M.; Futami, J. Mitochondrial determinants of mammalian longevity. Open Biol. 2017, 7, 170083. [Google Scholar] [CrossRef] [PubMed]
- Jove, M.; Mota-Martorell, N.; Fernàndez-Bernal, A.; Portero-Otin, M.; Barja, G.; Pamplona, R. Phenotypic molecular features of long-lived animal species. Free Radic. Biol. Med. 2023, 208, 728–747. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Mih, N.; Anand, A.; Park, J.H.; Tan, J.; Yurkovich, J.T.; Monk, J.M.; Lloyd, C.J.; Sandberg, T.E.; Seo, S.W.; et al. Cellular responses to reactive oxygen species are predicted from molecular mechanisms. Proc. Natl. Acad. Sci. USA 2019, 116, 14368–14373. [Google Scholar] [CrossRef] [PubMed]
- Saz, H.J. Energy metabolisms of parasitic helminths: Adaptations to parasitism. Annu. Rev. Physiol. 1981, 43, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J. The anaerobic end-products of helminths. Parasitology 1984, 88, 179–198. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.R.; Judge, D.S. Life Spans of Mammals, Birds, Amphibians, Reptiles, and Fish; Monographs on Population Aging No. 8; Odense University Press: Odense, Denmark, 2000. [Google Scholar]
- Bowler, J.K. Longevity of Reptiles and Amphibians in North American Collections as of 1 November 1975; Herpetological Circular No. 6; Society for the Study of Amphibians and Reptiles: Milwaukee, WI, USA, 1977. [Google Scholar]
- Oger, P.M.; Cario, A. Adaptation of the membrane in Archaea. Biophys. Chem. 2013, 183, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L. Anaerobic Gram-Positive Bacilli. In Clinical Microbiology Procedures Handbook, 3rd ed.; ASM Press: Washington, DC, USA, 2010; pp. 770–781. [Google Scholar]
- Stajich, J.E.; Block, D.; Boulez, K.; Brenner, S.E.; Chervitz, S.A.; Dagdigian, C.; Fuellen, G.; Gilbert, J.G.; Korf, I.; Lapp, H.; et al. The Bioperl toolkit: Perl modules for the life sciences. Genome Res. 2002, 12, 1611–1618. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.; von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Kitazoe, Y.; Kishino, H.; Hasegawa, M.; Nakajima, N.; Thorne, J.L.; Tanaka, M. Adaptive threonine increase in transmembrane regions of mitochondrial proteins in higher primates. PLoS ONE 2008, 3, e3343. [Google Scholar] [CrossRef] [PubMed]
- Tacutu, R.; Thornton, D.; Johnson, E.; Budovsky, A.; Barardo, D.; Craig, T.; Diana, E.; Lehmann, G.; Toren, D.; Wang, J.; et al. Human Ageing Genomic Resources: New and updated databases. Nucleic Acids Res. 2018, 46, D1083–D1090. [Google Scholar] [CrossRef]
- Fang, J.; Wong, H.S.; Brand, M.D. Production of superoxide and hydrogen peroxide in the mitochondrial matrix is dominated by site IQ of complex I in diverse cell lines. Redox Biol. 2020, 37, 101722. [Google Scholar] [CrossRef]
- Ong, G.; Logue, S.E. Unfolding the interactions between endoplasmic reticulum stress and oxidative stress. Antioxidants 2023, 12, 981. [Google Scholar] [CrossRef]
- Laddha, A.P.; Kulkarni, Y.A. NADPH oxidase: A membrane-bound enzyme and its inhibitors in diabetic complications. Eur. J. Pharmacol. 2020, 881, 173206. [Google Scholar] [CrossRef] [PubMed]
- Osawa, S.; Jukes, T.H.; Watanabe, K.; Muto, A. Recent evidence for evolution of the genetic code. Microbiol. Rev. 1992, 56, 229–264. [Google Scholar] [CrossRef]
- Sengupta, S.; Yang, X.; Higgs, P.G. The mechanisms of codon reassignments in mitochondrial genetic codes. J. Mol. Evol. 2007, 64, 662–688. [Google Scholar] [CrossRef]
- Swire, J.; Judson, O.P.; Burt, A. Mitochondrial genetic codes evolve to match amino acid requirements of proteins. J. Mol. Evol. 2005, 60, 128–139. [Google Scholar] [CrossRef]
- de Magalhães, J.P.; Costa, J.; Church, G.M. An analysis of the relationship between metabolism, developmental schedules, and longevity using phylogenetic independent contrasts. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2007, 62, 149–160. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Barja, G. Correlations with longevity and body size: To correct or not correct? J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2014, 69, 1096–1098. [Google Scholar] [CrossRef]
- Nishimura, A.; Yoshikawa, Y.; Ichikawa, K.; Takemoto, T.; Tanahashi, R.; Takagi, H. Longevity regulation by proline oxidation in yeast. Microorganisms 2021, 9, 1650. [Google Scholar] [CrossRef]
- Canfield, C.A.; Bradshaw, P. Amino acids in the regulation of aging and aging-related diseases. Transl. Med. Aging 2019, 3, 70–89. [Google Scholar] [CrossRef]
- Mota-Martorell, N.; Jové, M.; Berdún, R.; Òbis, È.; Barja, G.; Pamplona, R. Methionine metabolism is down-regulated in heart of long-lived mammals. Biology 2022, 11, 1821. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Suravajhala, P.; Rathnagiri, P.; Sreenivasulu, N. Intriguing role of proline in redox potential conferring high temperature stress tolerance. Front. Plant Sci. 2022, 13, 867531. [Google Scholar] [CrossRef] [PubMed]
- Mehlotra, R.K. Antioxidant defense mechanisms in parasitic protozoa. Crit. Rev. Microbiol. 1996, 22, 295–314. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Flores, A.; Martínez-González, J.J.; Rendón, J.L.; Del Arenal, I.P. The architecture of thiol antioxidant systems among invertebrate parasites. Molecules 2017, 22, 259. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Mo, W.; Shen, D.; Sun, L.; Wang, J.; Lu, S.; Gitschier, J.M.; Zhou, B. Yeast model uncovers dual roles of mitochondria in the action of artemisinin. PLoS Genet. 2005, 1, e36. [Google Scholar] [CrossRef] [PubMed]
- Löfmark, S.; Edlund, C.; Nord, C.E. Metronidazole is still the drug of choice for treatment of anaerobic infections. Clin. Infect. Dis. 2010, 50 (Suppl. S1), S16–S23. [Google Scholar] [CrossRef] [PubMed]
- Kavishe, R.A.; Koenderink, J.B.; Alifrangis, M. Oxidative stress in malaria and artemisinin combination therapy: Pros and Cons. FEBS J. 2017, 284, 2579–2591. [Google Scholar] [CrossRef] [PubMed]
- Mih, N.; Monk, J.M.; Fang, X.; Catoiu, E.; Heckmann, D.; Yang, L.; Palsson, B.O. Adaptations of Escherichia coli strains to oxidative stress are reflected in properties of their structural proteomes. BMC Bioinform. 2020, 21, 162. [Google Scholar] [CrossRef]
- Vieira-Silva, S.; Rocha, E.P. An assessment of the impacts of molecular oxygen on the evolution of proteomes. Mol. Biol. Evol. 2008, 25, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.L.; Stanley, J.A.; Robinson, M.C.; Sher, J.W.; Li, Z.; Chan, Y.A.; Omdahl, A.R.; Wattiez, R.; Godzik, A.; Matallana-Surget, S. Protein structure, amino acid composition and sequence determine proteome vulnerability to oxidation-induced damage. EMBO J. 2020, 39, e104523. [Google Scholar] [CrossRef]
- Copley, S.D.; Dhillon, J.K. Lateral gene transfer and parallel evolution in the history of glutathione biosynthesis genes. Genome Biol. 2002, 3, 0025. [Google Scholar] [CrossRef]
- Moosmann, B.; Schindeldecker, M.; Hajieva, P. Cysteine, glutathione and a new genetic code: Biochemical adaptations of the primordial cells that spread into open water and survived biospheric oxygenation. Biol. Chem. 2020, 401, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.N.; Rubinstein, N.D.; Buffenstein, R. A window into extreme longevity; the circulating metabolomic signature of the naked mole-rat, a mammal that shows negligible senescence. Geroscience 2018, 40, 105–121. [Google Scholar] [CrossRef]
- Mariadassou, M.; Pellay, F.X. Identification of amino acids in mitochondrially encoded proteins that correlate with lifespan. Exp. Gerontol. 2014, 56, 53–58. [Google Scholar] [CrossRef]
- Stadtman, E.R. Protein oxidation and aging. Free Radic. Res. 2006, 40, 1250–1258. [Google Scholar] [CrossRef]
- Geiger, T.; Clarke, S. Deamidation, isomerization, and racemization at asparaginyl and aspartyl residues in peptides. Succinimide-linked reactions that contribute to protein degradation. J. Biol. Chem. 1987, 262, 785–794. [Google Scholar] [CrossRef]
- Fujii, N.; Takata, T.; Fujii, N.; Aki, K.; Sakaue, H. D-Amino acids in protein: The mirror of life as a molecular index of aging. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 840–847. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Phylogenetic Group | Lifespan (Years) | Body Mass (kg) | Analyzed Sequences | ||
|---|---|---|---|---|---|---|
| Full proteome | Mitochondrion | Peroxisome | ||||
| Homo sapiens | Mammalia | 100 | 70 | 15,072 | 978 | 238 |
| Equus caballus | Mammalia | 62 | 1000 | 16,148 | 283 | 100 |
| Pan troglodytes | Mammalia | 60 | 45 | 14,349 | 376 | 168 |
| Macaca mulatta | Mammalia | 35 | 8 | 15,939 | 314 | 145 |
| Bos taurus | Mammalia | 30 | 750 | 28,590 | 749 | 110 |
| Canis familiaris | Mammalia | 30 | 40 | 14,903 | 429 | 227 |
| Gallus gallus | Aves | 30 | 2.6 | 15,525 | 301 | 101 |
| Strongylocentrotus purpuratus | Echinoidea | 20 | 0.1 | 25,696 | 309 | 29 |
| Oryctolagus cuniculus | Mammalia | 18 | 1.8 | 14,808 | 269 | 90 |
| Cavia porcellus | Mammalia | 15 | 0.73 | 16,419 | 204 | 59 |
| Monodelphis domestica | Mammalia | 6 | 0.11 | 16,593 | 273 | 46 |
| Mus musculus | Mammalia | 6 | 0.021 | 21,551 | 1186 | 188 |
| Apis mellifera | Insecta | 5 | 9 × 10−5 | 8529 | 204 | 19 |
| Danio rerio | Pisces | 5 | 5 × 10−4 | 25,049 | 588 | 62 |
| Rattus norvegicus | Mammalia | 4 | 0.3 | 26,060 | 799 | 158 |
| Ciona intestinalis | Ascidiae | 2 | 1 × 10−4 | 11,653 | 203 | 25 |
| Tribolium castaneum | Insecta | 2 | 2.5 × 10−6 | 9340 | 193 | 55 |
| Drosophila melanogaster | Insecta | 0.15 | 1 × 10−6 | 9748 | 162 | 45 |
| Anopheles gambiae | Insecta | 0.07 | 2.5 × 10−7 | 11,200 | 126 | 36 |
| Caenorhabditis elegans | Nematoda | 0.05 | 2 × 10−9 | 23,865 | 219 | 39 |
| Amino Acid | Peroxisome (Paradigm I) | Mitochondrion (Paradigm II) | Respiratory Chain (Paradigm III) | ||||
|---|---|---|---|---|---|---|---|
| Ratio | p | Ratio | p | Ratio | p | Decoding * | |
| A | 1.09 | 3 × 10−4 | 1.15 | 3 × 10−5 | 0.57 | 1 × 10−7 | |
| C | 0.86 | 3 × 10−5 | 0.74 | 1 × 10−7 | 0.41 | 1 × 10−7 | |
| D | 0.91 | 8 × 10−5 | 0.93 | 4 × 10−4 | 0.74 | 2 × 10−7 | |
| E | 0.91 | 3 × 10−7 | 0.92 | 2 × 10−6 | 0.63 | 6 × 10−8 | |
| F | 1.12 | 2 × 10−6 | 1.08 | 5 × 10−5 | 1.28 | 1 × 10−2 | |
| G | 1.09 | 9 × 10−5 | 1.12 | 4 × 10−4 | 0.73 | 1 × 10−6 | +AGA, +AGG (1/20) |
| H | 0.97 | 1 × 10−2 | 0.90 | 3 × 10−6 | 1.36 | 3 × 10−5 | |
| I | 1.19 | 1 × 10−4 | 1.12 | 2 × 10−3 | 1.12 | 4 × 10−3 | −AUA (19/20) |
| K | 0.97 | 7 × 10−2 | 1.04 | 2 × 10−1 | 0.79 | 4 × 10−5 | −AAA (1/20) |
| L | 1.05 | 1 × 10−2 | 1.06 | 6 × 10−3 | 1.11 | 2 × 10−7 | |
| M | 1.11 | 2 × 10−4 | 1.16 | 1 × 10−6 | 2.14 | 2 × 10−7 | +AUA (19/20) |
| N | 1.03 | 2 × 10−1 | 0.90 | 8 × 10−3 | 1.93 | 9 × 10−8 | +AAA (1/20) |
| P | 0.86 | 3 × 10−4 | 0.86 | 3 × 10−4 | 1.28 | 3 × 10−3 | |
| Q | 0.88 | 9 × 10−6 | 0.93 | 4 × 10−4 | 0.81 | 2 × 10−5 | |
| R | 0.92 | 2 × 10−3 | 1.03 | 2 × 10−2 | 0.49 | 6 × 10−8 | −AGA, −AGG (13/20) |
| S | 0.87 | 2 × 10−7 | 0.82 | 6 × 10−8 | 1.29 | 3 × 10−6 | +AGA, +AGG (6/20) |
| T | 0.99 | 6 × 10−1 | 0.97 | 1 × 10−1 | 1.48 | 7 × 10−3 | |
| V | 1.14 | 1 × 10−6 | 1.10 | 1 × 10−6 | 0.53 | 6 × 10−8 | |
| W | 1.10 | 2 × 10−3 | 1.05 | 3 × 10−2 | 1.81 | 6 × 10−8 | +UGA (20/20) |
| Y | 1.10 | 4 × 10−3 | 1.06 | 1 × 10−2 | 1.33 | 6 × 10−6 | |
| Amino Acid | Longevity within Paradigm I (Paradigm IVa) | Longevity within Paradigm II (Paradigm IVb) | Longevity within Paradigm III (Paradigm IVc) | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| A | −0.598 | 5 × 10−3 | 0.220 | 4 × 10−1 | 0.370 | 1 × 10−1 |
| C | −0.166 | 5 × 10−1 | 0.523 | 2 × 10−2 | −0.888 | 2 × 10−7 |
| D | 0.159 | 5 × 10−1 | 0.220 | 4 × 10−1 | 0.190 | 4 × 10−1 |
| E | 0.064 | 8 × 10−1 | −0.505 | 2 × 10−2 | 0.215 | 4 × 10−1 |
| F | 0.365 | 1 × 10−1 | −0.175 | 5 × 10−1 | −0.651 | 2 × 10−3 |
| G | −0.655 | 2 × 10−3 | −0.082 | 7 × 10−1 | −0.436 | 5 × 10−2 |
| H | −0.096 | 7 × 10−1 | 0.350 | 1 × 10−1 | 0.535 | 2 × 10−2 |
| I | 0.593 | 6 × 10−3 | 0.616 | 4 × 10−3 | 0.502 | 2 × 10−2 |
| K | 0.349 | 1 × 10−1 | −0.375 | 1 × 10−1 | −0.131 | 6 × 10−1 |
| L | −0.493 | 3 × 10−2 | −0.636 | 3 × 10−3 | −0.554 | 1 × 10−2 |
| M | 0.156 | 5 × 10−1 | 0.485 | 3 × 10−2 | −0.163 | 5 × 10−1 |
| N | 0.712 | 4 × 10−4 | 0.353 | 1 × 10−1 | 0.072 | 8 × 10−1 |
| P | −0.018 | 9 × 10−1 | 0.020 | 9 × 10−1 | 0.669 | 1 × 10−3 |
| Q | 0.671 | 2 × 10−3 | −0.154 | 5 × 10−1 | 0.686 | 8 × 10−4 |
| R | 0.099 | 7 × 10−1 | 0.053 | 8 × 10−1 | 0.321 | 2 × 10−1 |
| S | 0.510 | 2 × 10−2 | −0.369 | 1 × 10−1 | −0.484 | 3 × 10−2 |
| T | −0.012 | 1 × 10−0 | 0.490 | 3 × 10−2 | 0.697 | 6 × 10−4 |
| V | −0.352 | 1 × 10−1 | 0.219 | 4 × 10−1 | −0.562 | 1 × 10−2 |
| W | 0.236 | 3 × 10−1 | 0.241 | 3 × 10−1 | −0.474 | 3 × 10−2 |
| Y | −0.257 | 3 × 10−1 | 0.223 | 3 × 10−1 | −0.161 | 5 × 10−1 |
| Amino Acid | Raw | Body Mass-Corrected | Phylogeny-Corrected | Effect Size | |||||
|---|---|---|---|---|---|---|---|---|---|
| r | p | q [%] | r | p | r | p | Ratio | p | |
| A | 0.370 | 1 × 10−1 | 1 | −0.099 | 7 × 10−1 | 0.063 | 8 × 10−1 | 1.22 | 2 × 10−2 |
| C | −0.888 | 2 × 10−7 | 43 | −0.162 | 5 × 10−1 | −0.791 | 6 × 10−5 | 0.53 | 3 × 10−4 |
| D | 0.190 | 4 × 10−1 | 1 | −0.150 | 5 × 10−1 | −0.161 | 5 × 10−1 | 1.00 | 5 × 10−1 |
| E | 0.215 | 4 × 10−1 | 9 | −0.131 | 6 × 10−1 | 0.281 | 2 × 10−1 | 1.04 | 2 × 10−1 |
| F | −0.651 | 2 × 10−3 | 3 | 0.234 | 3 × 10−1 | −0.170 | 5 × 10−1 | 0.74 | 2 × 10−2 |
| G | −0.436 | 5 × 10−2 | 5 | 0.084 | 7 × 10−1 | −0.434 | 6 × 10−2 | 0.84 | 3 × 10−1 |
| H | 0.535 | 2 × 10−2 | 4 | −0.286 | 2 × 10−1 | −0.011 | 1 × 10−0 | 1.17 | 3 × 10−2 |
| I | 0.502 | 2 × 10−2 | 4 | −0.020 | 9 × 10−1 | 0.619 | 5 × 10−3 | 1.06 | 3 × 10−1 |
| K | −0.131 | 6 × 10−1 | 4 | −0.221 | 4 × 10−1 | −0.375 | 1 × 10−1 | 0.92 | 5 × 10−1 |
| L | −0.554 | 1 × 10−2 | 0 | 0.104 | 7 × 10−1 | −0.519 | 2 × 10−2 | 0.97 | 5 × 10−1 |
| M | −0.163 | 5 × 10−1 | 3 | −0.285 | 2 × 10−1 | −0.379 | 1 × 10−1 | 0.88 | 2 × 10−1 |
| N | 0.072 | 8 × 10−1 | 12 | −0.520 | 2 × 10−2 | −0.141 | 6 × 10−1 | 1.04 | 6 × 10−1 |
| P | 0.669 | 1 × 10−3 | 1 | 0.059 | 8 × 10−1 | 0.412 | 8 × 10−2 | 1.22 | 3 × 10−2 |
| Q | 0.686 | 8 × 10−4 | 2 | −0.081 | 7 × 10−1 | 0.292 | 2 × 10−1 | 1.27 | 1 × 10−2 |
| R | 0.321 | 2 × 10−1 | 1 | 0.325 | 2 × 10−1 | 0.473 | 4 × 10−2 | 1.02 | 6 × 10−1 |
| S | −0.484 | 3 × 10−2 | 1 | −0.007 | 1 × 10−0 | 0.177 | 5 × 10−1 | 0.93 | 4 × 10−1 |
| T | 0.697 | 6 × 10−4 | 0 | −0.247 | 3 × 10−1 | 0.145 | 6 × 10−1 | 1.52 | 7 × 10−3 |
| V | −0.562 | 1 × 10−2 | 4 | 0.019 | 9 × 10−1 | −0.577 | 1 × 10−2 | 0.88 | 1 × 10−1 |
| W | −0.474 | 3 × 10−2 | 2 | 0.133 | 6 × 10−1 | −0.028 | 9 × 10−1 | 0.91 | 7 × 10−2 |
| Y | −0.161 | 5 × 10−1 | 0 | −0.369 | 1 × 10−1 | −0.564 | 1 × 10−2 | 0.98 | 8 × 10−1 |
| Body mass | 0.933 | 2 × 10−9 | − | − | − | 0.886 | 5 × 10−7 | − | − |
| Amino Acid | Raw | Body Mass-Corrected | Phylogeny-Corrected | ||||
|---|---|---|---|---|---|---|---|
| r | p | q [%] | r | p | r | p | |
| A | 0.569 | 5 × 10−20 | 0 | 0.130 | 6 × 10−2 | −0.022 | 8 × 10−1 |
| C | −0.737 | 2 × 10−38 | 39 | −0.322 | 2 × 10−6 | −0.325 | 1 × 10−6 |
| D | 0.039 | 6 × 10−1 | 1 | −0.074 | 3 × 10−1 | 0.088 | 2 × 10−1 |
| E | 0.516 | 4× 10−16 | 5 | 0.387 | 4 × 10−9 | 0.119 | 8 × 10−2 |
| F | −0.740 | 5 × 10−39 | 1 | −0.226 | 1 × 10−3 | −0.133 | 5 × 10−2 |
| G | −0.036 | 6 × 10−1 | 1 | 0.005 | 9 × 10−1 | 0.004 | 1 × 10−0 |
| H | 0.706 | 4× 10−34 | 8 | 0.250 | 2 × 10−4 | 0.201 | 3 × 10−3 |
| I | −0.347 | 2 × 10−7 | 0 | −0.129 | 6 × 10−2 | 0.057 | 4 × 10−1 |
| K | −0.216 | 1 × 10−3 | 7 | 0.149 | 3 × 10−2 | 0.059 | 4 × 10−1 |
| L | 0.416 | 2 × 10−10 | 0 | 0.039 | 6 × 10−1 | 0.033 | 6 × 10−1 |
| M | −0.255 | 2 × 10−4 | 1 | −0.098 | 2 × 10−1 | −0.007 | 9 × 10−1 |
| N | −0.351 | 2 × 10−7 | 7 | −0.309 | 4 × 10−6 | −0.104 | 1 × 10−1 |
| P | 0.768 | 1 × 10−43 | 17 | 0.350 | 2 × 10−7 | 0.163 | 2 × 10−2 |
| Q | 0.610 | 2 × 10−23 | 0 | 0.096 | 2 × 10−1 | 0.005 | 9 × 10−1 |
| R | 0.475 | 2 × 10−13 | 4 | 0.077 | 3 × 10−1 | −0.052 | 4 × 10−1 |
| S | −0.620 | 2 × 10−24 | 0 | −0.105 | 1 × 10−1 | −0.036 | 6 × 10−1 |
| T | 0.747 | 4 × 10−40 | 1 | 0.236 | 5 × 10−4 | 0.167 | 1 × 10−2 |
| V | −0.183 | 7 × 10−3 | 1 | −0.022 | 7 × 10−1 | 0.024 | 7 × 10−1 |
| W | 0.329 | 7 × 10−7 | 0 | 0.111 | 1 × 10−1 | 0.172 | 1 × 10−2 |
| Y | −0.518 | 3 × 10−16 | 9 | −0.393 | 3 × 10−9 | −0.064 | 3 × 10−1 |
| Body mass | 0.868 | 2 × 10−67 | - | - | - | 0.555 | 6 × 10−19 |
| Amino Acid | Aerobicity (Paradigm V) | |
|---|---|---|
| Ratio | p | |
| A | 1.77 | 6 × 10−5 |
| C | 0.43 | 9 × 10−10 |
| D | 0.84 | 1 × 10−5 |
| E | 0.99 | 6 × 10−1 |
| F | 0.82 | 8 × 10−5 |
| G | 0.83 | 8 × 10−6 |
| H | 1.35 | 4 × 10−11 |
| I | 1.16 | 1 × 10−3 |
| K | 1.39 | 5 × 10−5 |
| L | 1.01 | 6 × 10−1 |
| M | 1.51 | 2 × 10−6 |
| N | 1.16 | 1 × 10−2 |
| P | 1.63 | 5 × 10−9 |
| Q | 1.77 | 9 × 10−12 |
| R | 1.04 | 2 × 10−1 |
| S | 0.93 | 5 × 10−5 |
| T | 1.68 | 1 × 10−9 |
| V | 0.58 | 3 × 10−12 |
| W | 1.03 | 6 × 10−1 |
| Y | 0.68 | 2 × 10−11 |
| Amino Acid | Archaea (Paradigm VI) | Gram-Positive Bacteria (Paradigm VII) | Gram-Negative Bacteria (Paradigm VIII) | |||
|---|---|---|---|---|---|---|
| Ratio | p | Ratio | p | Ratio | p | |
| A | 1.31 | 5 × 10−2 | 1.06 | 6 × 10−1 | 1.11 | 2 × 10−1 |
| C | 0.73 | 2 × 10−2 | 0.53 | 1 × 10−6 | 0.94 | 7 × 10−1 |
| D | 1.23 | 4 × 10−1 | 0.96 | 2 × 10−1 | 1.01 | 9 × 10−1 |
| E | 0.94 | 2 × 10−1 | 0.99 | 1 × 10+0 | 0.87 | 5 × 10−3 |
| F | 0.91 | 2 × 10−1 | 0.99 | 9 × 10−1 | 0.95 | 1 × 10−1 |
| G | 1.10 | 2 × 10−2 | 1.04 | 6 × 10−1 | 0.99 | 7 × 10−1 |
| H | 1.07 | 3 × 10−1 | 1.06 | 1 × 10−1 | 1.11 | 8 × 10−3 |
| I | 0.77 | 2 × 10−2 | 0.92 | 6 × 10−1 | 0.90 | 1 × 10−1 |
| K | 0.60 | 3 × 10−3 | 0.89 | 5 × 10−1 | 0.85 | 3 × 10−1 |
| L | 0.97 | 3 × 10−1 | 1.06 | 4 × 10−3 | 1.03 | 1 × 10−1 |
| M | 0.90 | 7 × 10−2 | 0.87 | 2 × 10−2 | 0.96 | 4 × 10−1 |
| N | 0.77 | 3 × 10−2 | 0.94 | 6 × 10−1 | 1.02 | 8 × 10−1 |
| P | 1.09 | 8 × 10−2 | 1.10 | 6 × 10−1 | 1.04 | 6 × 10−1 |
| Q | 1.07 | 2 × 10−1 | 1.05 | 5 × 10−1 | 1.33 | 8 × 10−5 |
| R | 1.24 | 3 × 10−2 | 1.06 | 7 × 10−1 | 0.99 | 8 × 10−1 |
| S | 1.03 | 8 × 10−1 | 0.94 | 1 × 10−1 | 1.00 | 1 × 10+0 |
| T | 1.16 | 5 × 10−2 | 1.07 | 1 × 10−2 | 1.02 | 4 × 10−1 |
| V | 1.12 | 2 × 10−2 | 1.04 | 3 × 10−1 | 1.01 | 8 × 10−1 |
| W | 1.06 | 6 × 10−1 | 1.11 | 3 × 10−1 | 1.18 | 2 × 10−3 |
| Y | 0.92 | 5 × 10−1 | 0.91 | 4 × 10−1 | 0.91 | 4 × 10−1 |
| Amino Acid | Peroxisome (I) | Mitochondrion (II) | Respiratory Chain (III) | Longevity (IV) | Aerobicity (V) |
|---|---|---|---|---|---|
| A | 14 | 5 | 7 | 5 | 2 * |
| C # | 2 * | 1 * | 1 * | 1 * | 1 * |
| D | 10 | 13 | 12 | 20 | 13 |
| E | 9 | 11 | 8 | 16 | 19 |
| F | 7 | 12 | 15 | 3 * | 11 |
| G | 13 | 7 | 10 | 7 | 12 |
| H | 17 | 9 | 11 | 8 | 10 |
| I | 1 * | 6 | 19 | 14 | 15 |
| K | 19 | 18 | 17 | 12 | 9 |
| L | 16 | 16 | 20 | 17 | 20 |
| M # | 8 | 3 * | 2 * | 10 | 7 |
| N | 18 | 8 | 4 * | 15 | 14 |
| P # | 3 * | 4 * | 16 | 6 | 6 |
| Q # | 6 | 14 | 18 | 4 * | 3 * |
| R | 15 | 20 | 3 * | 18 | 17 |
| S # | 4 * | 2 * | 14 | 13 | 16 |
| T | 20 | 19 | 9 | 2 * | 5 |
| V | 5 | 10 | 5 | 9 | 4 * |
| W | 12 | 17 | 6 | 11 | 18 |
| Y | 11 | 15 | 13 | 19 | 8 |
| Amino Acid | Archaea (VI) | Gram-Positive Bacteria (VII) | Gram-Negative Bacteria (VIII) |
|---|---|---|---|
| A | 3 * | 11 | 7 |
| C # | 2 * | 1 * | 9 |
| D | 7 | 17 | 16 |
| E | 17 | 20 | 4 * |
| F | 12 | 19 | 10 |
| G | 11 | 18 | 18 |
| H | 15 | 9 | 5 |
| I | 4 * | 7 | 6 |
| K # | 1 * | 3 * | 3 * |
| L | 19 | 14 | 13 |
| M | 10 | 2 * | 12 |
| N | 5 | 13 | 14 |
| P | 13 | 5 | 11 |
| Q | 16 | 15 | 1 * |
| R | 6 | 10 | 17 |
| S | 20 | 12 | 20 |
| T | 8 | 8 | 15 |
| V | 9 | 16 | 19 |
| W # | 18 | 4 * | 2 * |
| Y | 14 | 6 | 8 |
| Amino Acid | Perox (I) | Mito (II) | RC (III) | Long (IV) | Aero (V) | Arch (VI) | GPB (VII) | GNB (VIII) |
|---|---|---|---|---|---|---|---|---|
| C | −14 | −26 | −59 | −47 | −57 | −27 | −47 | −6 |
| M | +11 | +16 | +114 | −12 | +51 | −10 | −13 | −4 |
| W | +10 | +5 | +81 | −9 | +3 | +6 | +11 | +18 |
| Y | +10 | +6 | +33 | −2 | −32 | −8 | −9 | −9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schindeldecker, M.; Moosmann, B. Cysteine Is the Only Universally Affected and Disfavored Proteomic Amino Acid under Oxidative Conditions in Animals. Antioxidants 2024, 13, 267. https://doi.org/10.3390/antiox13030267
Schindeldecker M, Moosmann B. Cysteine Is the Only Universally Affected and Disfavored Proteomic Amino Acid under Oxidative Conditions in Animals. Antioxidants. 2024; 13(3):267. https://doi.org/10.3390/antiox13030267
Chicago/Turabian StyleSchindeldecker, Mario, and Bernd Moosmann. 2024. "Cysteine Is the Only Universally Affected and Disfavored Proteomic Amino Acid under Oxidative Conditions in Animals" Antioxidants 13, no. 3: 267. https://doi.org/10.3390/antiox13030267