
Deciphering the Crucial Roles of the Quorum-Sensing Transcription Factor SdiA in NADPH Metabolism and (S)-Equol Production in Escherichia coli Nissle 1917


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Chemicals
2.2. Plasmids and DNA Manipulation
2.3. Culture Conditions
2.4. SDS-PAGE Gel Electrophoresis
2.5. Determination of Intracellular NADPH Levels
2.6. Transcriptome Analysis by RNA-seq
2.7. Real-Time PCR Measurements for Transcriptional Analysis
2.8. Molecular Docking
2.9. MD Simulations
2.10. Analytical Methods
2.11. Statistical Analysis
3. Results and Discussion
3.1. Engineering E. coli Nissle 1917 for (S)-Equol Production from Daidzein and Daidzin through Chromosomal Integration
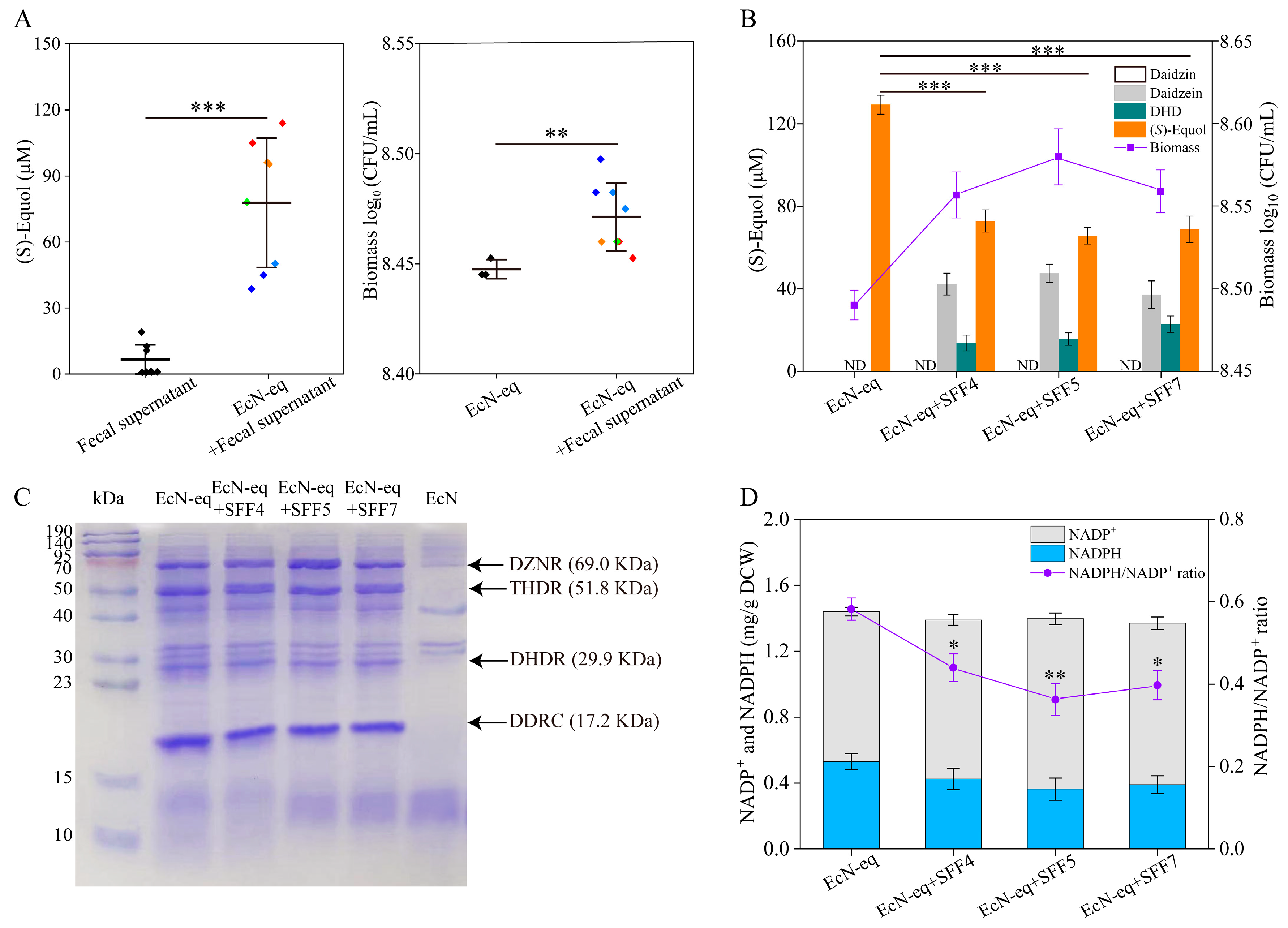
3.2. (S)-Equol Production by the EcN-eq Strain in Media with Fecal Supernatants or Sterile Fecal Filtrates
3.3. Transcriptome Analysis of the EcN-eq Strain in Media with or without the Addition of Sterile Fecal Filtrates
3.4. Quorum-Sensing Transcription Factor SdiA Modulates the Intracellular NADPH/NADP+ Ratio
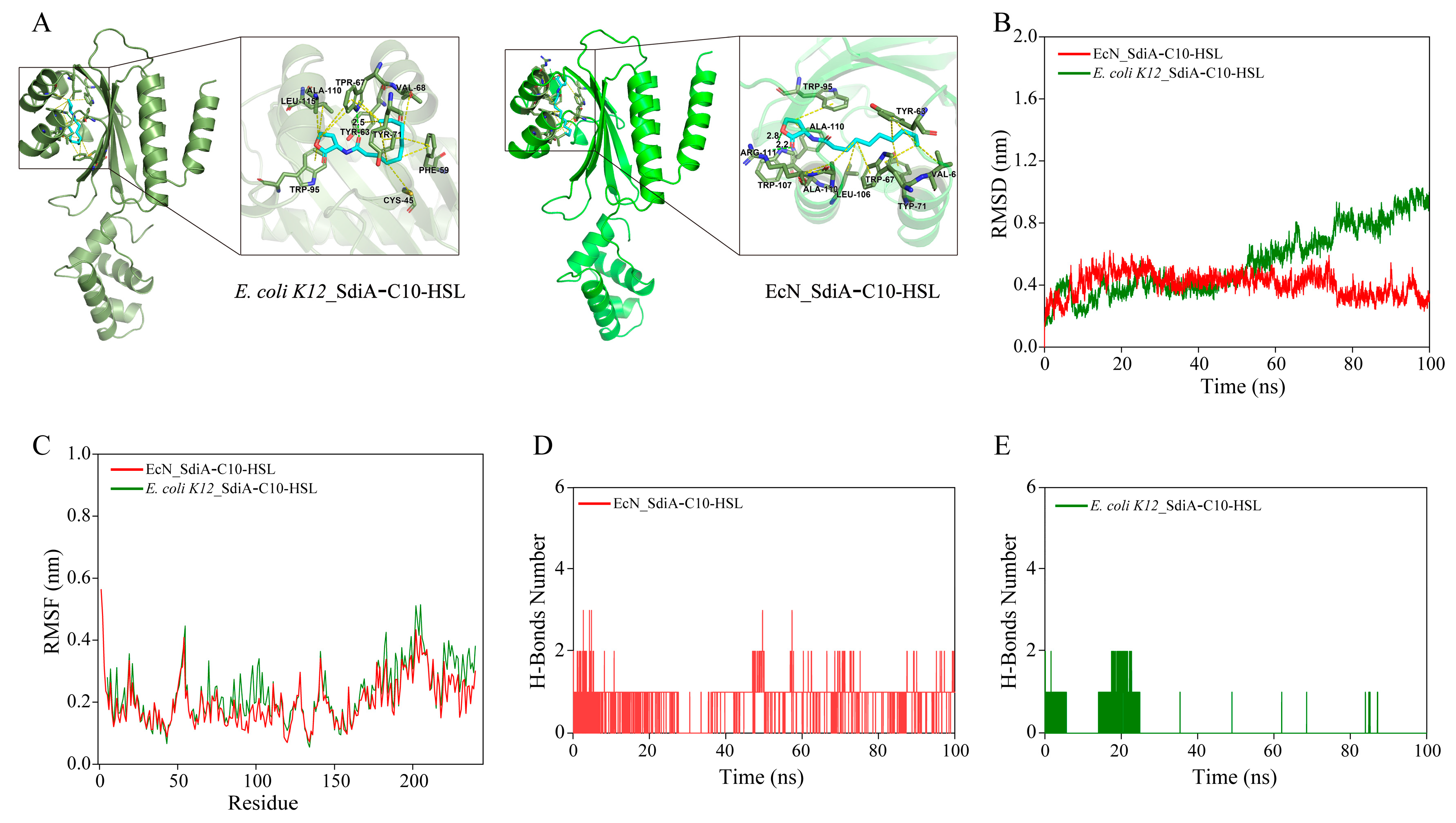
3.5. Interaction Analysis of N-Decanoyl-l-Homoserine Lactone (C10-HSL) with SdiA_EcN
3.6. Replacing sdiA_EcN with sdiA_K12 Improved (S)-Equol Production and Avoided Biomass Reduction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bešlo, D.; Došlić, G.; Agić, D.; Rastija, V.; Šperanda, M.; Gantner, V.; Lučić, B. Polyphenols in ruminant nutrition and their effects on reproduction. Antioxidants 2022, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-S. Current perspectives on the beneficial effects of soybean isoflavones and their metabolites for humans. Antioxidants 2021, 10, 1064. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, X.; Azi, F.; Dai, Y.; Xu, Z.; Yu, L.; Zhou, J.; Dong, M.; Xia, X. Biosynthesis of (S)-equol from soy whey by metabolically engineered Escherichia coli. J. Agric. Food Chem. 2023, 71, 6110–6119. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, B.; Fang, X.; Zhong, Q.; Liao, Z.; Wang, J.; Wu, X.; Ma, Y.; Li, P.; Feng, X. Soy isoflavone-specific biotransformation product S-equol in the colon: Physiological functions, transformation mechanisms, and metabolic regulatory pathways. Crit. Rev. Food Sci. 2022, 62, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Cole, S.J. Method of defining equol-producer status and its frequency among vegetarians. J. Nutr. 2006, 136, 2188–2193. [Google Scholar] [CrossRef] [PubMed]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and characterisation of an equol-producing mixed microbial culture from a human faecal sample and its activity under gastrointestinal conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Zhao, L.; Zhang, J.; Fang, X.; Zhong, Q.; Liao, Z.; Wang, J.; Guo, Y.; Liang, H.; Wang, L. Colonization potential to reconstitute a microbe community in pseudo germ-free mice after fecal microbe transplant from equol producer. Front. Microbiol. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Deng, H.; Gao, S.; Zhang, W.; Zhang, T.; Li, N.; Zhou, J. High titer of (S)-equol synthesis from daidzein in Escherichia coli. ACS Synth. Biol. 2022, 11, 4043–4053. [Google Scholar] [CrossRef]
- Effendi, S.S.W.; Ng, I.S. Prospective and challenges of live bacterial therapeutics from a superhero Escherichia coli Nissle 1917. Crit. Rev. Microbiol. 2023, 49, 611–627. [Google Scholar] [CrossRef]
- Yu, Y.; Gong, B.; Wang, H.; Yang, G.; Zhou, X. Chromosome evolution of Escherichia coli Nissle 1917 for high-level production of heparosan. Biotechnol. Bioeng. 2023, 120, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Kydd, L.; Shiveshwarkar, P.; Jaworski, J. Engineering Escherichia coli for conversion of dietary isoflavones in the gut. ACS Synth. Biol. 2022, 11, 3575–3582. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Yasuda, S.; Takahashi, M.; Hayashi, T.; Miyazawa, N.; Sato, I.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Cloning and expression of a novel NADP(H)-dependent daidzein reductase, an enzyme involved in the metabolism of daidzein, from equol-producing Lactococcus strain 20-92. Appl. Environ. Microb. 2010, 76, 5892–5901. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Tian, P.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Quorum sensing of lactic acid bacteria: Progress and insights. Food Rev. Int. 2023, 39, 4781–4792. [Google Scholar] [CrossRef]
- Spangler, J.R.; Dean, S.N.; Leary, D.H.; Walper, S.A. Response of Lactobacillus plantarum WCFS1 to the Gram-negative pathogen-associated quorum sensing molecule N-3-oxododecanoyl homoserine lactone. Front. Microbiol. 2019, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Chi, L.; Tu, P.; Lai, Y.; Liu, C.-W.; Ru, H.; Lu, K. Detection of gut microbiota and pathogen produced N-acyl homoserine in host circulation and tissues. NPJ Biofilms Microbiomes 2021, 7, 53. [Google Scholar] [CrossRef]
- Nguyen, Y.; Nguyen Nam, X.; Rogers Jamie, L.; Liao, J.; MacMillan John, B.; Jiang, Y.; Sperandio, V. Structural and mechanistic roles of novel chemical ligands on the SdiA quorum-sensing transcription regulator. mBio 2015, 6, 10–1128. [Google Scholar] [CrossRef]
- Shimada, T.; Shimada, K.; Matsui, M.; Kitai, Y.; Igarashi, J.; Suga, H.; Ishihama, A. Roles of cell division control factor SdiA: Recognition of quorum sensing signals and modulation of transcription regulation targets. Genes Cells 2014, 19, 405–418. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, X.; Zheng, J.; Zou, Y.; Qiu, S.; Dai, Y. AHL-mediated quorum sensing to regulate bacterial substance and energy metabolism: A review. Microbiol. Res. 2022, 30, 127102. [Google Scholar] [CrossRef]
- Carneiro, D.G.; Almeida, F.A.; Aguilar, A.P.; Vieira, N.M.; Pinto, U.M.; Mendes, T.A.; Vanetti, M.C.D. Salmonella enterica optimizes metabolism after addition of acyl-homoserine lactone under anaerobic conditions. Front. Microbiol. 2020, 11, 1459. [Google Scholar] [CrossRef]
- Yan, H.; Li, J.; Meng, J.; Li, J.; Jha, A.K.; Zhang, Y.; Wang, X.; Fan, Y. Insight into the effect of N-acyl-homoserine lactones-mediated quorum sensing on the microbial social behaviors in a UASB with the regulation of alkalinity. Sci. Total Environ. 2021, 800, 149413. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Li, X.; Zhang, G.; Zhu, Y.; Bi, J.; Hao, H.; Hou, H. Quorum sensing-mediated and growth phase-dependent regulation of metabolic pathways in Hafnia alvei H4. Front. Microbiol. 2021, 12, 567942. [Google Scholar] [CrossRef]
- Yan, X.; Liu, X.-Y.; Zhang, D.; Zhang, Y.-D.; Li, Z.-H.; Liu, X.; Wu, F.; Chen, G.-Q. Construction of a sustainable 3-hydroxybutyrate-producing probiotic Escherichia coli for treatment of colitis. Cell Mol. Immunol. 2021, 18, 2344–2357. [Google Scholar] [CrossRef] [PubMed]
- Leventhal, D.S.; Sokolovska, A.; Li, N.; Plescia, C.; Kolodziej, S.A.; Gallant, C.W.; Christmas, R.; Gao, J.-R.; James, M.J.; Abin-Fuentes, A.; et al. Immunotherapy with engineered bacteria by targeting the STING pathway for anti-tumor immunity. Nat. Commun. 2020, 11, 2739. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dai, Y.; Azi, F.; Wang, Z.; Xu, W.; Wang, D.; Dong, M.; Xia, X. Engineering Escherichia coli for cost-effective production of medium-chain fatty acids from soy whey using an optimized galactose-based autoinduction system. Bioresour. Technol. 2024, 393, 130145. [Google Scholar] [CrossRef]
- Wang, Z.; Li, X.; Dai, Y.; Yin, L.; Azi, F.; Zhou, J.; Dong, M.; Xia, X. Sustainable production of genistin from glycerol by constructing and optimizing Escherichia coli. Metab. Eng. 2022, 74, 206–219. [Google Scholar] [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef]
- Xu, P.; Vansiri, A.; Bhan, N.; Koffas, M.A.G. ePathBrick: A synthetic biology platform for engineering metabolic pathways in E. coli. ACS Synth. Biol. 2012, 1, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhang, C.; Zhang, X.; Zhao, X.; Chaeipeima Mahsa, G.; Ma, K.; Ji, F.; Azarpazhooh, E.; Ajami, M.; Rui, X.; et al. Effects of Lacticaseibacillus paracasei SNB-derived postbiotic components on intestinal barrier dysfunction and composition of gut microbiota. Food Res. Int. 2024, 175, 113773. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Ionescu, V.S.; Popa, A.; Alexandru, A.; Manole, E.; Neagu, M.; Pop, S. Dietary phytoestrogens and their metabolites as epigenetic modulators with impact on human health. Antioxidants 2021, 10, 1893. [Google Scholar] [CrossRef]
- Fiechter, G.; Raba, B.; Jungmayr, A.; Mayer, H.K. Characterization of isoflavone composition in soy-based nutritional supplements via ultra performance liquid chromatography. Anal. Chim. Acta 2010, 672, 72–78. [Google Scholar] [CrossRef]
- Wichmann, J.; Behrendt, G.; Boecker, S.; Klamt, S. Characterizing and utilizing oxygen-dependent promoters for efficient dynamic metabolic engineering. Metab. Eng. 2023, 77, 199–207. [Google Scholar] [CrossRef]
- Arcidiacono, S.; Spangler, J.R.; Litteral, V.; Doherty, L.A.; Stamps, B.; Walper, S.; Goodson, M.; Soares, J.W. In vitro fermentation evaluation of engineered sense and respond probiotics in polymicrobial communities. ACS Biomater. Sci. Eng. 2023, 9, 5176–5185. [Google Scholar] [CrossRef]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; Moltzau-Anderson, J.; Bharti, R.; Grasis, J.A.; Cassidy, L.; Tholey, A.; Fickenscher, H.; Seegert, D. Efficacy of sterile fecal filtrate transfer for treating patients with Clostridium difficile infection. Gastroenterology 2017, 152, 799–811.e797. [Google Scholar] [CrossRef]
- Tsuji, H.; Moriyama, K.; Nomoto, K.; Akaza, H. Identification of an enzyme system for daidzein-to-equol conversion in Slackia sp. strain NATTS. Appl. Environ. Microbiol. 2012, 78, 1228–1236. [Google Scholar] [CrossRef]
- Shimada, Y.; Takahashi, M.; Miyazawa, N.; Ohtani, T.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Identification of two novel reductases involved in equol biosynthesis in Lactococcus strain 20-92. J. Mol. Microbiol 2011, 21, 160–172. [Google Scholar] [CrossRef]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.-J. Antioxidant potential and capacity of microorganism-sourced C30 carotenoids—A review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef]
- Kim, M.; Jung, D.-H.; Hwang, C.Y.; Siziya, I.N.; Park, Y.-S.; Seo, M.-J. 4,4′-Diaponeurosporene production as C30 carotenoid with antioxidant activity in recombinant Escherichia coli. Appl. Biochem. 2023, 195, 135–151. [Google Scholar] [CrossRef]
- Lee, P.G.; Kim, J.; Kim, E.J.; Jung, E.; Pandey, B.P.; Kim, B.G. P212A Mutant of dihydrodaidzein reductase enhances (S)-equol production and enantioselectivity in a recombinant Escherichia coli whole-cell reaction system. Appl. Environ. Microbiol. 2016, 82, 1992–2002. [Google Scholar] [CrossRef]
- Wang, L.; Yu, H.; Xu, J.; Ruan, H.; Zhang, W. Deciphering the crucial roles of AraC-type transcriptional regulator Cgl2680 on NADPH metabolism and L-lysine production in Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2020, 36, 1–15. [Google Scholar] [CrossRef]
- Kim, T.; Duong, T.; Wu, C.-a.; Choi, J.; Lan, N.; Kang, S.W.; Lokanath, N.K.; Shin, D.; Hwang, H.-Y.; Kim, K.K. Structural insights into the molecular mechanism of Escherichia coli SdiA, a quorum-sensing receptor. Acta Crystallogr. D 2014, 70, 694–707. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, M.; Hardwidge, P.R.; Cui, H.; Zhu, G. Isolation and characterization of N-acyl homoserine lactone-producing bacteria from cattle rumen and swine intestines. Front. Cell. Infect. Microbiol. 2018, 8, 155. [Google Scholar] [CrossRef]
- Gu, J.; Xu, Y.; Nie, Y. Role of distal sites in enzyme engineering. Biotechnol. Adv. 2023, 63, 108094. [Google Scholar] [CrossRef]
- Xu, X.; Zeng, D.; Wu, D.; Lin, J. Single-point mutation near active center increases substrate affinity of alginate lyase AlgL-CD. Appl. Biochem. 2021, 193, 1513–1531. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, C.; Dong, W.; Lu, H.; Yang, Y.; Li, W.; Xu, Y.; Li, X. Simultaneously enhanced thermostability and catalytic activity of xylanase from Streptomyces rameus L2001 by rigidifying flexible regions in loop regions of the N-terminus. J. Agric. Food Chem. 2023, 71, 12785–12796. [Google Scholar] [CrossRef]
- Pedrolli, D.B.; Ribeiro, N.V.; Squizato, P.N.; de Jesus, V.N.; Cozetto, D.A. Engineering microbial living therapeutics: The synthetic biology toolbox. Trends Biotechnol. 2019, 37, 100–115. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Properties | Source |
|---|---|---|
| E. coli DH5α (collection no.: DSM 6897) | F−, φ80d lacZΔM15, Δ(lacZYA-argF)U169, recA1, endA1, hsdR17(rk−, mk+), phoA, supE44λ−, thi−, gyrA96, relA1 | Invitrogen (Invitrogen, Carlsbad, CA, USA) |
| EcN (collection no.: DSM 115365) | Wild-type E. coli Nissle 1917 | Lab stock |
| EcN-eq | EcN, malK::Pnar-dznr-Pnar-ddrc-Pnar-dhdr-Pnar-thdr, exo/cea::Pnar-bglF-Pnar-bglB, ΔptsG::KanR | This study |
| EcN-eq ΔdecR | EcN-eq, ΔdecR | This study |
| EcN-eq ΔHW372_01960 | EcN-eq, ΔHW372_01960 | This study |
| EcN-eq ΔyhjC | EcN-eq, ΔyhjC | This study |
| EcN-eq ΔHW372_03545 | EcN-eq, ΔHW372_03545 | This study |
| EcN-eq ΔsdiA | EcN-eq, ΔsdiA | This study |
| EcN-eq ΔyhaJ | EcN-eq, ΔyhaJ | This study |
| EcN-eq pETM6-Pnar-decR | EcN-eq carrying pETM6-Pnar-decR | This study |
| EcN-eq pETM6-Pnar-HW372_01960 | EcN-eq carrying pETM6-Pnar-HW372_01960 | This study |
| EcN-eq pETM6-Pnar-yhjC | EcN-eq carrying pETM6-Pnar-yhjC | This study |
| EcN-eq pETM6-Pnar-HW372_03545 | EcN-eq carrying pETM6-Pnar-HW372_03545 | This study |
| EcN-eq pETM6-Pnar-sdiA | EcN-eq carrying pETM6-Pnar-sdiA | This study |
| EcN-eq pETM6-Pnar-yhaJ | EcN-eq carrying pETM6-Pnar-yhaJ | This study |
| EcN-eq ΔsdiA::sdiA_K12 | EcN-eq, ΔsdiA::sdiA_K12 | This study |
| Gene | Annotation | log2 (Fold Change) |
|---|---|---|
| Transporters | ||
| yagG | sugar transporter | −2.9905 |
| ydcS | polyamine transporter | −2.2575 |
| fetB | iron-export ABC-transporter ATPase | 2.5437 |
| gatA | galactitol PTS | 1.5344 |
| Pentose phosphate pathway | ||
| zwf | glucose-6-phosphate 1-dehydrogenase | −1.8321 |
| gnd | 6-phosphogluconate dehydrogenase | −2.6247 |
| Acetate, anaplerotic, and other gluconeogenic pathways | ||
| poxB | 6-phosphofructokinase | 2.7182 |
| acs | acetyl-CoA synthetase | 1.5124 |
| pfkA | 6-phosphofructokinase I | 2.1273 |
| gpmM | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase | 1.5162 |
| Stress-response pathway | ||
| rsfA | ribosome-silencing factor | 1.7628 |
| yfcV | stress-response fimbriae | 2.2365 |
| ypjA | stress-response adhesin | 1.9634 |
| Cell to cell interaction | ||
| hlyE | hemolysin E | 2.6343 |
| ypfA | adhesion-like autotransporter | 1.7762 |
| fimA | major type-I fimbrin | 3.7689 |
| yqjH | siderophore interaction protein | 2.1547 |
| Transcriptional regulator | ||
| decR | AsnC-family transcriptional regulator | −1.9354 |
| ascG | transcriptional regulator | 3.2329 |
| yhjC | LysR-family transcriptional regulator | 2.7514 |
| frvR | transcriptional regulator | −2.1672 |
| sdiA | LuxI-family transcriptional regulator | 3.4855 |
| yhaJ | transcriptional regulator | 2.7841 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Dai, Y.; Azi, F.; Dong, M.; Xia, X. Deciphering the Crucial Roles of the Quorum-Sensing Transcription Factor SdiA in NADPH Metabolism and (S)-Equol Production in Escherichia coli Nissle 1917. Antioxidants 2024, 13, 259. https://doi.org/10.3390/antiox13030259
Wang Z, Dai Y, Azi F, Dong M, Xia X. Deciphering the Crucial Roles of the Quorum-Sensing Transcription Factor SdiA in NADPH Metabolism and (S)-Equol Production in Escherichia coli Nissle 1917. Antioxidants. 2024; 13(3):259. https://doi.org/10.3390/antiox13030259
Chicago/Turabian StyleWang, Zhe, Yiqiang Dai, Fidelis Azi, Mingsheng Dong, and Xiudong Xia. 2024. "Deciphering the Crucial Roles of the Quorum-Sensing Transcription Factor SdiA in NADPH Metabolism and (S)-Equol Production in Escherichia coli Nissle 1917" Antioxidants 13, no. 3: 259. https://doi.org/10.3390/antiox13030259