
Oxidative Stress in Parasitic Diseases—Reactive Oxygen Species as Mediators of Interactions between the Host and the Parasites



Abstract
:1. Introduction
2. Methods
3. Oxidant–Antioxidant Balance
3.1. Reactive Oxygen Species Formation
3.2. Antioxidant Defense Mechanisms
3.3. Oxidative Stress
4. Oxidative Stress in the Host
4.1. Protozoa Infections

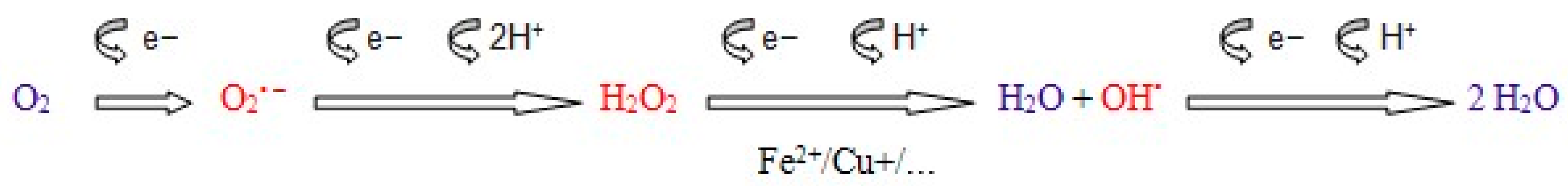{kind=link}
| Study Group | Parasite | Results | Ref. |
|---|---|---|---|
| 72 patients with malaria, 40 healthy subjects (control) | Plasmodium falciparum | decreased vit. A, vit. C and vit. E levels, decreased GSH activity, increased MDA level in malaria patients | [83] |
| 63 patients with malaria, 67 healthy subjects (control) | Plasmodium spp. | decreased SOD and GSH activities and increased MDA level in malaria patients | [76] |
| 17 patients with complicated malaria, 51 patients with uncomplicated malaria, 15 healthy subjects (control) | Plasmodium spp. | decreased vit. C level, increased MDA level in malaria patients | [84] |
| 551 patients with malaria, 221 healthy subjects (control) | Plasmodium spp. | decreased GST, SOD, and CAT activities, increased MDA level in malaria patients | [85] |
| 60 patients with malaria, 30 healthy subjects (control) | Plasmodium falciparum | decreased vit. C level, decreased SOD activity in malaria patients | [86] |
| 80 patients with malaria, 80 healthy subjects (control) | Plasmodium falciparum | decreased vit. A, vit. C, vit. E levels, decreased GSH activity, increased MDA level in malaria patients | [87] |
| 20 infected patients, 20 patients with diarrhea, 30 healthy subjects (control) | Entamoeba histolytica | decreased GSH, CAT, and SOD activities, increased MDA level in infected patients | [88] |
| 50 infected patients (giardiasis), 32 infected patients (toxoplasmosis), 40 healthy subjects (control) | Giardia duodenalis, Toxoplasma gondii | decreased GSH activity, increased MDA and NO levels in infected patients | [89] |
| 44 seropositive pregnant women, 40 healthy pregnant women (control) | Toxoplasma gondii | decreased vit. C and vit. E levels, decreased GSH, SOD, GPx, and CAT activities, increased MDA level in seropositive women | [90] |
| 50 seropositive patients, 30 healthy subjects (control) | Toxoplasma gondii | decreased GSH activity, increased MDA level in seropositive patients | [91] |
| 50 seropositive patients, 50 healthy subjects (control) | Toxoplasma gondii | increased MDA level in seropositive patients | [92] |
| 65 infected patients, 50 healthy subjects (control) | Leishmania donovani | decreased CAT activity, increased MDA level in infected patients | [93] |
| 28 infected patients, 10 healthy subjects (control) | Leishmania braziliensis | increased NO production in monocytes of infected patients | [94] |
4.2. Helminth Infections
| Study Group | Parasite | Results | Ref. |
|---|---|---|---|
| 40 infected patients (enterobiasis), 46 infected patients (echinococcosis), 40 healthy subjects (control) | Enterobius vermicularis, Echinococcus granulosus | decreased GSH activity, increased MDA and NO levels in infected patients | [89] |
| 20 infected patients, 10 healthy subjects (control) | Fasciola hepatica | decreased GPx and SOD activities, increased CAT activity, increased MDA level in infected patients | [105] |
| 140 infected patients, 140 healthy subjects (control) | Fasciola hepatica | increased GPx, CAT, and SOD activities, increased MDA level in infected patients | [106] |
| 12 infected rats, 12 uninfected rats (control) | Fasciola hepatica | increased NOX and TBARS levels in infected rats | [107] |
| 29 infected dogs, 16 uninfected dogs (control) | Ancylostoma spp. | increased TAC level in infected dogs | [108] |
| 22 infected cattle (fasciolosis), 30 infected cattle (cysticercosis), 40 healthy cattle (control) | Fasciola hepatica, Cysticersus bovis | decreased GSH, GST, SOD, and CAT activities, increased MDA levels in infected cattle | [109] |
5. Oxidative Stress in the Parasite
5.1. Protozoa Infections
5.2. Helminth Infections
6. Conclusions
Author Contributions
Funding
Institutional Review Board
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cable, J.; Barber, I.; Boag, B.; Ellison, A.R.; Morgan, E.R.; Murray, K.; Pascoe, E.L.; Sait, S.M.; Wilson, A.J.; Booth, M. Global Change, Parasite Transmission and Disease Control: Lessons from Ecology. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160088. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. The Rise of Ecological Parasitology: Twelve Landmark Advances That Changed Its History. Int. J. Parasitol. 2021, 51, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Alves, F.; Bucheton, B.; Burrows, L.; Büscher, P.; Carrillo, E.; Felger, I.; Hübner, M.P.; Moreno, J.; Pinazo, M.J.; et al. Implications of Asymptomatic Infection for the Natural History of Selected Parasitic Tropical Diseases. Semin. Immunopathol. 2020, 42, 231–246. [Google Scholar] [CrossRef]
- Çenesiz, S. The Role of Oxidant and Antioxidant Parameters in the Infectious Diseases: A Systematic Literature Review. Kafkas Univ. Vet. Fak. Derg. 2020, 26, 849–858. [Google Scholar] [CrossRef]
- Maldonado, E.; Rojas, D.A.; Morales, S.; Miralles, V.; Solari, A. Dual and Opposite Roles of Reactive Oxygen Species (ROS) in Chagas Disease: Beneficial on the Pathogen and Harmful on the Host. Oxid. Med. Cell. Longev. 2020, 2020, 8867701. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk-Golec, K.; Pawłowska, M.; Wesołowski, R.; Wróblewski, M.; Mila-Kierzenkowska, C. Oxidative Stress as a Possible Target in the Treatment of Toxoplasmosis: Perspectives and Ambiguities. Int. J. Mol. Sci. 2021, 22, 5705. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.K.; Zhou, J. Role of Reactive Oxygen Species and Iron in Host Defense against Infection. Front. Biosci. 2020, 1, 50–56. [Google Scholar]
- Sorci, G.; Faivre, B. Inflammation and Oxidative Stress in Vertebrate Host-Parasite Systems. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 71–83. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Shaw, G.H. Earth’s Atmosphere—Hadean to Early Proterozoic. Chem. Erde 2008, 68, 235–264. [Google Scholar] [CrossRef]
- Piera, J.; Bäckvall, J.E. Catalytic Oxidation of Organic Substrates by Molecular Oxygen and Hydrogen Peroxide by Multistep Electron Transfer—A Biomimetic Approach. Angew. Chemie Int. Ed. 2008, 47, 3506–3523. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Venditti, P. Evolution of the Knowledge of Free Radicals and Other Oxidants. Oxid. Med. Cell. Longev. 2020, 2020, 9829176. [Google Scholar] [CrossRef] [PubMed]
- Saibi, W.; Brini, F. Superoxide Dismutase (SOD) And Abiotic Stress Tolerance in Plants: An Overview. In Superoxide Dismutase; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2004; pp. 3–5. ISBN 9780470962916. [Google Scholar]
- Kremer, M.L. The Fenton Reaction. Dependence of the Rate on PH. J. Phys. Chem. A 2003, 107, 1734–1741. [Google Scholar] [CrossRef]
- Gligorovski, S.; Strekowski, R.; Barbati, S.; Vione, D. Environmental Implications of Hydroxyl Radicals (•OH). Chem. Rev. 2015, 115, 13051–13092. [Google Scholar] [CrossRef] [PubMed]
- Ifeanyi, O.E. A Review on Free Radicals and Antioxidants. Infect. Disord. Drug Targets 2022, 20, 16–26. [Google Scholar] [CrossRef]
- Żukowski, P.; Maciejczyk, M.; Waszkiel, D. Sources of Free Radicals and Oxidative Stress in the Oral Cavity. Arch. Oral Biol. 2018, 92, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Larosa, V.; Remacle, C. Insights into the Respiratory Chain and Oxidative Stress. Biosci. Rep. 2018, 38, BSR20171492. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Venditti, P. Mitochondrial Management of Reactive Oxygen Species. Antioxidants 2021, 10, 1824. [Google Scholar] [CrossRef]
- Del Río, L.A.; López-Huertas, E. ROS Generation in Peroxisomes and Its Role in Cell Signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Schenkman, J.B.; Jansson, I. The Many Roles of Cytochrome B5. Pharmacol. Ther. 2003, 97, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Ozougwu, J. The Role of Reactive Oxygen Species and Antioxidants in Oxidative Stress. Int. J. Res. Pharm. Biosci. 2016, 3, 1–8. [Google Scholar]
- Dixon, S.J.; Stockwell, B.R. The Role of Iron and Reactive Oxygen Species in Cell Death. Nat. Chem. Biol. 2014, 10, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; de la Lastra, J.M.P.; Plou, F.J.; Pérez-Lebeña, E.; Reinbothe, S. Molecular Sciences the Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Gerschman, R.; Gilbert, D.L.; Nye, S.W.; Dwyer, P.; Fenn, W.O. Oxygen Poisoning and X-Irradiation: A Mechanism in Common. Science 1954, 119, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Fereidoon, S.; Ying, Z. Lipid Oxidation and Improving the Oxidative Stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, Y. Erratum: What Is Responsible for the Initiating Chemistry of Iron-Mediated Lipid Peroxidation: An Update (Chemical Reviews (2007) Vol. 107 (749–766)). Chem. Rev. 2007, 107, 2165. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of Lipid Peroxidation by Measuring Malondialdehyde (MDA) and Relatives in Biological Samples: Analytical and Biological Challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Jiang, S.; Carroll, L.; Rasmussen, L.M.; Davies, M.J. Oxidation of Protein Disulfide Bonds by Singlet Oxygen Gives Rise to Glutathionylated Proteins. Redox Biol. 2021, 38, 101822. [Google Scholar] [CrossRef]
- Di Mascio, P.; Martinez, G.R.; Miyamoto, S.; Ronsein, G.E.; Medeiros, M.H.G.; Cadet, J. Singlet Molecular Oxygen Reactions with Nucleic Acids, Lipids, and Proteins. Chem. Rev. 2019, 119, 2043–2086. [Google Scholar] [CrossRef]
- Barnes, J.L.; Zubair, M.; John, K.; Poirier, M.C.; Martin, F.L. Carcinogens and DNA Damage. Biochem. Soc. Trans. 2018, 46, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I. Reactive Oxygen Species and the Free Radical Theory of Aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martín, V.; Reiter, R.J. Regulation of Antioxidant Enzymes: A Significant Role for Melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Fahimi, H.D. Mammalian Peroxisomes and Reactive Oxygen Species. Histochem. Cell Biol. 2004, 122, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Shariati, M.A.; Rebezov, M.; Rengasamy, K.R.R.; Mubarak, M.S. Superoxide Dismutase: An Updated Review on Its Health Benefits and Industrial Applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 7282–7300. [Google Scholar] [CrossRef]
- Eleutherio, E.C.A.; Magalhães, R.S.S.; de Araújo Brasil, A.; Neto, J.R.M.; de Holanda Paranhos, L. SOD1, More than Just an Antioxidant. Arch. Biochem. Biophys. 2021, 697, 108701. [Google Scholar] [CrossRef]
- Mruk, D.D.; Silvestrini, B.; Mo, M.Y.; Cheng, C.Y. Antioxidant Superoxide Dismutase—A Review: Its Function, Regulation in the Testis, and Role in Male Fertility. Contraception 2002, 65, 305–311. [Google Scholar] [CrossRef]
- Kim, Y.S.; Vallur, P.G.; Phaëton, R.; Mythreye, K.; Hempel, N. Insights into the Dichotomous Regulation of SOD2 in Cancer. Antioxidants 2017, 6, 86. [Google Scholar] [CrossRef]
- Kliment, C.R.; Tobolewski, J.M.; Manni, M.L.; Tan, R.J.; Enghild, J.; Oury, T.D. Extracellular Superoxide Dismutase Protects against Matrix Degradation of Heparan Sulfate in the Lung. Antioxid. Redox Signal. 2008, 10, 261–268. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- .Sepasi Tehrani, H.; Moosavi-Movahedi, A.A. Catalase and Its Mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione Peroxidase Family—An Evolutionary Overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.; Torramade-Moix, S.; Martin-Rodriguez, S.; Cases, A.; Cruzado, J.M.; Rivera, J.; Escolar, G.; Palomo, M.; Diaz-Ricart, M. Antioxidant and Anti-Inflammatory Strategies Based on the Potentiation of Glutathione Peroxidase Activity Prevent Endothelial Dysfunction in Chronic Kidney Disease. Cell. Physiol. Biochem. 2018, 51, 1287–1300. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Gomes, D.; Lima, M.; Bronze-Da-Rocha, E.; Santos-Silva, A. Peroxiredoxin 2, Glutathione Peroxidase, and Catalase in the Cytosol and Membrane of Erythrocytes under H2O2-Induced Oxidative Stress. Free Radic. Res. 2015, 49, 990–1003. [Google Scholar] [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive Oxygen Species, Toxicity, Oxidative Stress, and Antioxidants: Chronic Diseases and Aging; Springer: Berlin/Heidelberg, Germany, 2023; Volume 97, ISBN 0123456789. [Google Scholar]
- Husain, N.; Kumar, A. Reactive Oxygen Species and Natural Antioxidants: A Review. Adv. Biores. 2012, 3, 164–175. [Google Scholar]
- Parcheta, M.; Świsłocka, R.; Orzechowska, S.; Akimowicz, M.; Choińska, R.; Lewandowski, W. Recent Developments in Effective Antioxidants: The Structure and Antioxidant Properties. Materials 2021, 14, 1984. [Google Scholar] [CrossRef]
- Zingg, J.-M.; Azzi, A. Non-Antioxidant Activities of Vitamin E. Curr. Med. Chem. 2012, 11, 1113–1133. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Vitamin D Deficiency: Effects on Oxidative Stress, Epigenetics, Gene Regulation, and Aging. Biology 2019, 8, 30. [Google Scholar] [CrossRef]
- Davies, K.J.A. Oxidative Stress, Antioxidant Defenses, and Damage Removal, Repair, and Replacement Systems. IUBMB Life 2000, 50, 279–289. [Google Scholar] [CrossRef]
- Altieri, F.; Grillo, C.; Maceroni, M.; Chichiarelli, S. DNA Damage and Repair: From Molecular Mechanisms to Health Implications. Antioxid. Redox Signal. 2008, 10, 891–937. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative Stress and Autophagy: The Clash between Damage and Metabolic Needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef]
- Jeong, D.W.; Kim, T.S.; Cho, I.T.; Kim, I.Y. Modification of Glycolysis Affects Cell Sensitivity to Apoptosis Induced by Oxidative Stress and Mediated by Mitochondria. Biochem. Biophys. Res. Commun. 2004, 313, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Ermak, G.; Davies, K.J.A. Calcium and Oxidative Stress: From Cell Signaling to Cell Death. Mol. Immunol. 2002, 38, 713–721. [Google Scholar] [CrossRef]
- Gibson, G.E.; Zhang, H.; Xu, H.; Park, L.C.H.; Jeitner, T.M. Oxidative Stress Increases Internal Calcium Stores and Reduces a Key Mitochondrial Enzyme. Biochim. Biophys. Acta Mol. Basis Dis. 2002, 1586, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, Á.; Valério-Bolas, A.; Palma-Marques, J.; Mourata-Gonçalves, P.; Ruas, P.; Dias-Guerreiro, T.; Santos-Gomes, G. Cutaneous Leishmaniasis: The Complexity of Host’s Effective Immune Response against a Polymorphic Parasitic Disease. J. Immunol. Res. 2019, 2019, 2603730. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Role of Oxidative Stress in Infectious Diseases. A Review. Folia Microbiol. 2013, 58, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative Stress in Malaria Parasite-Infected Erythrocytes: Host-Parasite Interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef]
- Rossi, M.; Fasel, N. How to Master the Host Immune System? Leishmania Parasites Have the Solutions! Int. Immunol. 2018, 30, 103–111. [Google Scholar] [CrossRef]
- Sato, S. Correction to: Plasmodium—A Brief Introduction to the Parasites Causing Human Malaria and Their Basic Biology. J. Physiol. Anthropol. 2021, 40, 3. [Google Scholar] [CrossRef]
- Varo, R.; Chaccour, C.; Bassat, Q. Update on Malaria. Med. Clín. 2020, 155, 395–402. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Cebrián-Carmona, J.; González-Cerón, L.; Manzano-Agugliaro, F.; Mesa-Valle, C. Analysis of Global Research on Malaria and Plasmodium vivax. Int. J. Environ. Res. Public Health 2019, 16, 1928. [Google Scholar] [CrossRef] [PubMed]
- Ohiagu, F.O.; Chikezie, P.C.; Ahaneku, C.C.; Chikezie, C.M.; Law-Obi, F.C. Pathophysiology of Severe Malaria Infection. Asian J. Health Sci. 2021, 7, 22. [Google Scholar] [CrossRef]
- Atiku, S.M.; Louise, N.; Kasozi, D.M. Severe Oxidative Stress in Sickle Cell Disease Patients with Uncomplicated Plasmodium falciparum Malaria in Kampala, Uganda. BMC Infect. Dis. 2019, 19, 600. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, M.; Zuniga, M.; Rodriguez, A. Oxidative Stress and Pathogenesis in Malaria. Front. Cell. Infect. Microbiol. 2021, 11, 768182. [Google Scholar] [CrossRef]
- Sarkar, A.; Basak, S.; Ghosh, S.; Mahalanobish, S.; Sil, P.C. Insights into the Relation between Oxidative Stress and Malaria: A Mechanistic and Therapeutic Approach. React. Oxyg. Species 2021, 11, m26–m41. [Google Scholar] [CrossRef]
- Boulet, C.; Doerig, C.D.; Carvalho, T.G. Manipulating Eryptosis of Human Red Blood Cells: A Novel Antimalarial Strategy? Front. Cell. Infect. Microbiol. 2018, 8, 419. [Google Scholar] [CrossRef] [PubMed]
- White, N.J. Anaemia and Malaria. Malar. J. 2018, 17, 371. [Google Scholar] [CrossRef] [PubMed]
- Varela, E.L.P.; Gomes, A.R.Q.; Santos, A.d.S.B.d.; de Carvalho, E.P.; Vale, V.V.; Percário, S. Potential Benefits of Lycopene Consumption: Rationale for Using It as an Adjuvant Treatment for Malaria Patients and in Several Diseases. Nutrients 2022, 14, 5303. [Google Scholar] [CrossRef] [PubMed]
- Sovira, N.; Lubis, M.; Wahidiyat, P.A.; Suyatna, F.D.; Gatot, D.; Bardosono, S.; Sadikin, M. Effects of α-Tocopherol on Hemolysis and Oxidative Stress Markers on Red Blood Cells in β-Thalassemia Major. Clin. Exp. Pediatr. 2020, 63, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Aqeel, S.; Naheda, A.; Raza, A.; Khan, W. Biomarkers of Disease Severity in Vivax Malaria. Parasitol. Res. 2021, 120, 1437–1446. [Google Scholar] [CrossRef]
- Ebrahim, A.; Gnanasekaran, N.; Genet, S. Oxidative Stress and Diminished Total Antioxidant Capacity in Malaria Patients Correspond to Increased Parasitemia and Severity of the Disease. React. Oxyg. Species 2019, 8, 287e96. [Google Scholar] [CrossRef]
- Babalola, A.S.; Jonathan, J.; Michael, B.E. Oxidative Stress and Anti-Oxidants in Asymptomatic Malaria-Positive Patients: A Hospital-Based Cross-Sectional Nigerian Study. Egypt. J. Intern. Med. 2020, 32, 23. [Google Scholar] [CrossRef]
- Kapoor, G. Oxidative Stress in Plasmodium: Role of Glutathione Revisited. Bull. Pure Appl. Sci. Zool. 2022, 41, 149–157. [Google Scholar] [CrossRef]
- Joardar, N.; Sinha Babu, S.P. A Review on the Druggability of a Thiol-Based Enzymatic Antioxidant Thioredoxin Reductase for Treating Filariasis and Other Parasitic Infections. Int. J. Biol. Macromol. 2020, 142, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Charvat, R.A.; Arrizabalaga, G. Oxidative Stress Generated during Monensin Treatment Contributes to Altered Toxoplasma gondii Mitochondrial Function. Sci. Rep. 2016, 6, 22997. [Google Scholar] [CrossRef]
- Machado-Silva, A.; Cerqueira, P.G.; Grazielle-Silva, V.; Gadelha, F.R.; Peloso, E.d.F.; Teixeira, S.M.R.; Machado, C.R. How Trypanosoma cruzi Deals with Oxidative Stress: Antioxidant Defence and DNA Repair Pathways. Mutat. Res. Rev. Mutat. Res. 2016, 767, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Tanowitz, H.B.; Garg, N.J. Pathogenesis of Chronic Chagas Disease: Macrophages, Mitochondria, and Oxidative Stress. Curr. Clin. Microbiol. Rep. 2018, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Conceição-Silva, F.; Morgado, F.N. Leishmania Spp.-Host Interaction: There Is Always an Onset, but Is There an End? Front. Cell. Infect. Microbiol. 2019, 9, 330. [Google Scholar] [CrossRef] [PubMed]
- Abduljalil, M.M.; Abubakar, M.G.; Danjuma, M.A. Antioxidant Status of Children Infected with Plasmodium falciparum Malaria in Kebbi Metropolis, Northwestern Nigeria. Int. J. Trop. Dis. 2021, 4, 53. [Google Scholar] [CrossRef]
- Nsiah, K.; Bahaah, B.; Afranie, B.O.; Koffie, S.; Akowuah, E.; Donkor, S. Oxidative Stress and Hemoglobin Level of Complicated and Uncomplicated Malaria Cases among Children: A Cross-Sectional Study in Kumasi Metropolis, Ghana. J. Trop. Med. 2019, 2019, 8479076. [Google Scholar] [CrossRef]
- Tyagi, A.G.; Tyagi, R.A.; Choudhary, P.R.; Shekhawat, J.S. Study of Antioxidant Status in Malaria Patients. Int. J. Res. Med. Sci. 2017, 5, 1649. [Google Scholar] [CrossRef]
- Al Ezzi, A.A. Changes in Levels of Antioxidant Markers and Status of Some Enzyme Activities among Falciparum Malaria Patients in Yemen. J. Microbiol. Exp. 2017, 4, 00131. [Google Scholar] [CrossRef]
- Abubakar, M.G.; Usman, S.M.; Dandare, S.U. Oxidant Status of Children Infected with Plasmodium falciparum Malaria in Katsina Metropolis, Northwestern Nigeria. Afr. J. Infect. Dis. 2016, 10, 17–20. [Google Scholar] [CrossRef]
- Mohsin, L.A.; Hamad, S.S.; Rahim, S.M. The Effect of Infection with the Entamoeba histolytica on Oxidative Stress Status in Kirkuk Hospital Patients. J. Pharm. Negat. Results 2022, 13, 3191–3195. [Google Scholar] [CrossRef]
- Kiran, T.; Karaman, U.; Arici, Y.; Yildiz, S. Comparison of Malondialdehyde, Nitric Oxide, Adenosine Deaminase and Glutathione Levels in Patients with Entamoeba coli, Enterobius vermicularis, Giardia intestinalis, Demodex Spp. Positive, Hydatid Cyst and Toxoplasma Gondii Serum Positive. Ann. Med. Res. 2019, 26, 1420. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Rajab, K.I.; Al-rawi, K.F.; Al-darwesh, M.Y. Evaluation Some Antioxidants and Oxidative Stress Index in Seropositive Toxoplasmosis in Pregnant Women in Ramadi City of Iraq. Sys. Rev. Pharm. 2020, 11, 701–705. [Google Scholar]
- Al-Azzauy, A.A.M. Evaluation of Erythrocyte Malondialdehyde, Glutathione Concentration and Serum Nitric Oxide Levels in Patients with Toxoplasma gondii. Ibn AL Haitham J. Pure Appl. Sci. 2011, 24, 35–40. [Google Scholar]
- Yazar, S.; Kilic, E.; Saraymen, R.; Sahin, I. Serum Malondialdehyde Levels in Toxoplasma Seropositive Patients. Ann. Saudi Med. 2003, 23, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Ali, W.R. Role of Some Cytokines and Oxidative Stress Enzymes in Iraqi Children Infected with Visceral Leishmaniasis. J. Univ. Babylon 2016, 24, 1485–1492. [Google Scholar]
- Carneiro, P.P.; Conceição, J.; Macedo, M.; Magalhães, V.; Carvalho, E.M.; Bacellar, O. The Role of Nitric Oxide and Reactive Oxygen Species in the Killing of Leishmania braziliensis by Monocytes from Patients with Cutaneous Leishmaniasis. PLoS ONE 2016, 11, e014808. [Google Scholar] [CrossRef] [PubMed]
- Karamon, J.; Dabrowska, J.; Kochanowski, M.; Samorek-Pieróg, M.; Sroka, J.; Różycki, M.; Bilska-Zajac, E.; Zdybel, J.; Cencek, T. Prevalence of Intestinal Helminths of Red Foxes (Vulpes Vulpes) in Central Europe (Poland): A Significant Zoonotic Threat. Parasites Vectors 2018, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Meade, T.M.; Watson, J. Parasitic Diseases; Academic Press: Cambridge, MA, USA, 2019; ISBN 9780128143384. [Google Scholar]
- Baral, R.; Jha, P.; Amatya, R.; Khanal, B. Prevalence of Intestinal Parasitic Infections among Patients Attending in a Tertiary Care Hospital of Eastern Region of Nepal—A Retrospective, Laboratory Based Study. Asian J. Med. Sci. 2017, 8, 55–59. [Google Scholar] [CrossRef]
- Early, R. Foodborne Parasites; Springer: Berlin/Heidelberg, Germany, 2008; Volume 43, ISBN 9783319676623. [Google Scholar]
- Mendlovic, F.; Fleury, A.; Flisser, A. Zoonotic Taenia Infections with Focus on Cysticercosis Due to Taenia solium in Swine and Humans. Res. Vet. Sci. 2021, 134, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Chifunda, K.; Kelly, P. Parasitic Infections of the Gut in Children. Paediatr. Int. Child Health 2019, 39, 65–72. [Google Scholar] [CrossRef]
- Cancela, M.; Paes, J.A.; Moura, H.; Barr, J.R.; Zaha, A.; Ferreira, H.B. Unraveling Oxidative Stress Response in the Cestode Parasite Echinococcus granulosus. Sci. Rep. 2019, 9, 15876. [Google Scholar] [CrossRef]
- Brosschot, T.P.; Reynolds, L.A. The Impact of a Helminth-Modified Microbiome on Host Immunity Review-Article. Mucosal Immunol. 2018, 11, 1039–1046. [Google Scholar] [CrossRef]
- Motran, C.C.; Silvane, L.; Chiapello, L.S.; Theumer, M.G.; Ambrosio, L.F.; Volpini, X.; Celias, D.P.; Cervi, L. Helminth Infections: Recognition and Modulation of the Immune Response by Innate Immune Cells. Front. Immunol. 2018, 9, 664. [Google Scholar] [CrossRef]
- Jabbar, K.D. Biochemical Evaluation of Antioxidant Enzyme Activities and Lipid Peroxidation Level Associated with Liver Enzymes in Patients with Fascioliasis. Arch. Razi Inst. 2022, 77, 1067–1073. [Google Scholar] [CrossRef]
- Kamel, H.H.; Sarhan, R.M.; Saad, G.A. Biochemical Assessment of Oxidative Status versus Liver Enzymes in Patients with Chronic Fascioliasis. J. Parasit. Dis. 2015, 39, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, Z.T.; Yılmaz, H.; Beyhan, Y.E.; Ekici, A.; Çiçek, M.; Aydemir, S. The Importance of Antioxidant Enzymes and Oxidative Stress in Human Fascioliasis. Turk. Parazitoloji Derg. 2023, 47, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Bottari, N.B.; Mendes, R.E.; Lucca, N.J.; Schwertz, C.I.; Henker, L.C.; Olsson, D.C.; Piva, M.M.; Sangoi, M.; Campos, L.P.; Moresco, R.N.; et al. Oxidative Stress Associated with Pathological Lesions in the Liver of Rats Experimentally Infected by Fasciola hepatica. Exp. Parasitol. 2015, 159, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.M.S.; Tvarijonaviciute, A.; Martinez-Subiela, S.; Cerón, J.J.; Eckersall, P.D. Changes in Biochemical Analytes in Female Dogs with Subclinical Ancylostoma Spp. Infection. BMC Vet. Res. 2016, 12, 203. [Google Scholar] [CrossRef] [PubMed]
- Atteya, M.A.; Wahba, A.A.; Ghobashy, M.A.; Dessouky, A.A. Oxidative Stress and Histopathological Changes in Cattle Affected with Fascioliasis and Cysticercosis. Egypt. JM Sci. 2015, 36, 191–204. [Google Scholar]
- DeMichele, E.; Sosnowski, O.; Buret, A.G.; Allain, T. Regulatory Functions of Hypoxia in Host–Parasite Interactions: A Focus on Enteric, Tissue, and Blood Protozoa. Microorganisms 2023, 11, 1598. [Google Scholar] [CrossRef]
- Masamba, P.; Kappo, A.P. Immunological and Biochemical Interplay between Cytokines, Oxidative Stress and Schistosomiasis. Int. J. Mol. Sci. 2021, 22, 7216. [Google Scholar] [CrossRef]
- Tiwari, S.; Sharma, N.; Sharma, G.P.; Mishra, N. Redox Interactome in Malaria Parasite Plasmodium falciparum. Parasitol. Res. 2021, 120, 423–434. [Google Scholar] [CrossRef]
- Kotepui, M.; Mahittikorn, A.; Anabire, N.G.; Kotepui, K.U. Impact of Malaria on Glutathione Peroxidase Levels: A Systematic Review and Meta-Analysis. Sci. Rep. 2023, 13, 13928. [Google Scholar] [CrossRef]
- Lesanavičius, M.; Aliverti, A.; Šarlauskas, J.; Čėnas, N. Reactions of Plasmodium falciparum Ferredoxin:Nadp+ Oxidoreductase with Redox Cycling Xenobiotics: A Mechanistic Study. Int. J. Mol. Sci. 2020, 21, 3234. [Google Scholar] [CrossRef]
- Maroziene, A.; Lesanavičius, M.; Davioud-Charvet, E.; Aliverti, A.; Grellier, P.; Šarlauskas, J.; Čenas, N. Antiplasmodial Activity of Nitroaromatic Compounds: Correlation with Their Reduction Potential and Inhibitory Action on Plasmodium falciparum Glutathione Reductase. Molecules 2019, 24, 4509. [Google Scholar] [CrossRef]
- Raza, A.; Manzoor, B.; Akhtar, A.; Kaushal, N.; Varshney, S.K.; Khan, H.M.; Malik, M.A. Superoxide Dismutase Activity in Erythrocytes Infected With Plasmodium falciparum and Plasmodium vivax, A Comparison. East Afr. Sch. J. Med. Sci. 2019, 4421, 2–5. [Google Scholar]
- Rodríguez-Saavedra, C.; Morgado-Martínez, L.E.; Burgos-Palacios, A.; King-Díaz, B.; López-Coria, M.; Sánchez-Nieto, S. Moonlighting Proteins: The Case of the Hexokinases. Front. Mol. Biosci. 2021, 8, 701975. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, A.I.; Panova, A.S.; Kokova, A.D.; Kotova, D.A.; Maltsev, D.I.; Podgorny, O.V.; Belousov, V.V.; Bilan, D.S. In Vivo Imaging with Genetically Encoded Redox Biosensors. Int. J. Mol. Sci. 2020, 21, 8164. [Google Scholar] [CrossRef]
- Wang, B.; Pakpour, N.; Napoli, E.; Drexler, A.; Glennon, E.K.K.; Surachetpong, W.; Cheung, K.; Aguirre, A.; Klyver, J.M.; Lewis, E.E.; et al. Anopheles Stephensi P38 MAPK Signaling Regulates Innate Immunity and Bioenergetics during Plasmodium falciparum Infection. Parasites Vectors 2015, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, A. Thioredoxin and Redox Signaling: Roles of the Thioredoxin System in Control of Cell Fate. Arch. Biochem. Biophys. 2017, 617, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Balsera, M.; Buchanan, B.B. Evolution of the Thioredoxin System as a Step Enabling Adaptation to Oxidative Stress. Free Radic. Biol. Med. 2019, 140, 28–35. [Google Scholar] [CrossRef] [PubMed]
- McCarty, S.E.; Schellenberger, A.; Goodwin, D.C.; Fuanta, N.R.; Tekwani, B.L.; Calderón, A.I. Plasmodium falciparum Thioredoxin Reductase (PfTrxR) and Its Role as a Target for New Antimalarial Discovery. Molecules 2015, 20, 11459. [Google Scholar] [CrossRef]
- Fata, F.; Gencheva, R.; Cheng, Q.; Lullo, R.; Ardini, M.; Silvestri, I.; Gabriele, F.; Ippoliti, R.; Bulman, C.A.; Sakanari, J.A.; et al. Biochemical and Structural Characterizations of Thioredoxin Reductase Selenoproteins of the Parasitic Filarial Nematodes Brugia malayi and Onchocerca volvulus. Redox Biol. 2022, 51, 102278. [Google Scholar] [CrossRef]
- Wang, W.; Huang, P.; Jiang, N.; Lu, H.; Zhang, D.; Wang, D.; Zhang, K.; Wahlgren, M.; Chen, Q. A Thioredoxin Homologous Protein of Plasmodium falciparum Participates in Erythrocyte Invasion. Infect. Immun. 2018, 86, e00289-18. [Google Scholar] [CrossRef]
- Andrade, R.M.; Reed, S.L. New Drug Target in Protozoan Parasites: The Role of Thioredoxin Reductase. Front. Microbiol. 2015, 6, 975. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.P.; Chitnis, C.E. Lipid Peroxidation and Its Repair in Malaria Parasites. Trends Parasitol. 2023, 39, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Zou, L.; Wang, J.; Chasapis, C.T.; Peana, M. Thioredoxin Reductase as a Pharmacological Target. Pharmacol. Res. 2021, 174, 105854. [Google Scholar] [CrossRef]
- Vašková, J.; Kočan, L.; Vaško, L.; Perjési, P. Glutathione-Related Enzymes and Proteins: A Review. Molecules 2023, 28, 1447. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, E.; Novichkova, M. Glutathione in Protein Redox Modulation through S-Glutathionylation and s-Nitrosylation. Molecules 2021, 26, 435. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, R.; Sharma, S.; Patankar, S. Glutathione and Thioredoxin Systems of the Malaria Parasite Plasmodium falciparum: Partners in Crime? Biochem. Biophys. Res. Commun. 2017, 488, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; Hartley, R.C. Mitochondria as a Therapeutic Target for Common Pathologies. Nat. Rev. Drug Discov. 2018, 17, 865–886. [Google Scholar] [CrossRef] [PubMed]
- Haeussler, K.; Fritz-Wolf, K.; Reichmann, M.; Rahlfs, S.; Becker, K. Characterization of Plasmodium falciparum 6-Phosphogluconate Dehydrogenase as an Antimalarial Drug Target. J. Mol. Biol. 2018, 430, 4049–4067. [Google Scholar] [CrossRef] [PubMed]
- Kori, L.D.; Valecha, N.; Anvikar, A.R. Glutamate Dehydrogenase: A Novel Candidate to Diagnose Plasmodium falciparum through Rapid Diagnostic Test in Blood Specimen from Fever Patients. Sci. Rep. 2020, 10, 6307. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-Transferase: A Versatile Protein Family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Von Mengden, L.; Klamt, F.; Smitz, J. Redox Biology of Human Cumulus Cells: Basic Concepts, Impact on Oocyte Quality, and Potential Clinical Use. Antioxid. Redox Signal. 2020, 32, 522–535. [Google Scholar] [CrossRef]
- Tafreshi, A.S.; Sadeghi, S.; Hekmatipour, M.; Shams, S.; Mohammadi, M.; Vahabi, F.; Arjmand, M.; Zamani, Z. The Effect of Some Antimalarial Drugs on Plasmodium berghei Glutathione-S-Transferase Enzyme. J. Microb. World 2019, 12, 84–90. [Google Scholar]
- Egwu, C.O.; Pério, P.; Augereau, J.M.; Tsamesidis, I.; Benoit-Vical, F.; Reybier, K. Resistance to Artemisinin in Falciparum Malaria Parasites: A Redox-Mediated Phenomenon. Free Radic. Biol. Med. 2022, 179, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.M.S.; Carvalho, A.R., Jr.; Lacerda, E.M.d.C.B.; da Silva, L.C.N.; Marinho, C.R.F.; André, E.; Fernandes, E.S. Oxidative and Nitrosative Stresses in Cerebral Malaria: Can We Target Them to Avoid a Bad Prognosis? J. Antimicrob. Chemother. 2020, 75, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Chulanetra, M.; Chaicumpa, W. Revisiting the Mechanisms of Immune Evasion Employed by Human Parasites. Front. Cell. Infect. Microbiol. 2021, 11, 702125. [Google Scholar] [CrossRef] [PubMed]
- Bosch, S.S.; Kronenberger, T.; Meissner, K.A.; Zimbres, F.M.; Stegehake, D.; Izui, N.M.; Schettert, I.; Liebau, E.; Wrenger, C. Oxidative Stress Control by Apicomplexan Parasites. Biomed Res. Int. 2015, 2015, 351289. [Google Scholar] [CrossRef] [PubMed]
- Goes, G.R.; Rocha, P.S.; Diniz, A.R.S.; Aguiar, P.H.N.; Machado, C.R.; Vieira, L.Q. Trypanosoma cruzi Needs a Signal Provided by Reactive Oxygen Species to Infect Macrophages. PLoS Negl. Trop. Dis. 2016, 10, e0004555. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, J.; Xu, M.; Shan, D.; Wu, Z.; Yuan, D. Speciation and Adaptive Evolution Reshape Antioxidant Enzymatic System Diversity across the Phylum Nematoda. BMC Biol. 2020, 18, 181. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Zhou, T.; Pannell, B.K.; Ziegler, A.C.; Best, T.M. Biological and Physiological Role of Reactive Oxygen Species—The Good, the Bad and the Ugly. Acta Physiol. 2015, 214, 329–348. [Google Scholar] [CrossRef]
- Selkirk, M.E.; Smith, V.P.; Thomas, G.R.; Gounaris, K. Resistance of Filarial Nematode Parasites to Oxidative Stress. Int. J. Parasitol. 1998, 28, 1315–1332. [Google Scholar] [CrossRef]
- Sureshan, M.; Prabhu, D.; Aruldoss, I.; Saraboji, K. Potential Inhibitors for Peroxiredoxin 6 of W. bancrofti: A Combined Study of Modelling, Structure-Based Drug Design and MD Simulation. J. Mol. Graph. Model. 2022, 112, 108115. [Google Scholar] [CrossRef]
- de Oliveira, M.A.; Tairum, C.A.; Netto, L.E.S.; de Oliveira, A.L.P.; Aleixo-Silva, R.L.; Cabrera, V.I.M.; Breyer, C.A.; dos Santos, M.C. Relevance of Peroxiredoxins in Pathogenic Microorganisms. Appl. Microbiol. Biotechnol. 2021, 105, 5701–5717. [Google Scholar] [CrossRef] [PubMed]
- Dorey, A.; Cwiklinski, K.; Rooney, J.; De Marco Verissimo, C.; López Corrales, J.; Jewhurst, H.; Fazekas, B.; Calvani, N.E.D.; Hamon, S.; Gaughan, S.; et al. Autonomous Non Antioxidant Roles for Fasciola hepatica Secreted Thioredoxin-1 and Peroxiredoxin-1. Front. Cell. Infect. Microbiol. 2021, 11, 667272. [Google Scholar] [CrossRef] [PubMed]
- Miazek, K.; Beton, K.; Śliwińska, A.; Brożek-Płuska, B. The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used. Biomolecules 2022, 12, 1087. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawłowska, M.; Mila-Kierzenkowska, C.; Szczegielniak, J.; Woźniak, A. Oxidative Stress in Parasitic Diseases—Reactive Oxygen Species as Mediators of Interactions between the Host and the Parasites. Antioxidants 2024, 13, 38. https://doi.org/10.3390/antiox13010038
Pawłowska M, Mila-Kierzenkowska C, Szczegielniak J, Woźniak A. Oxidative Stress in Parasitic Diseases—Reactive Oxygen Species as Mediators of Interactions between the Host and the Parasites. Antioxidants. 2024; 13(1):38. https://doi.org/10.3390/antiox13010038
Chicago/Turabian StylePawłowska, Marta, Celestyna Mila-Kierzenkowska, Jan Szczegielniak, and Alina Woźniak. 2024. "Oxidative Stress in Parasitic Diseases—Reactive Oxygen Species as Mediators of Interactions between the Host and the Parasites" Antioxidants 13, no. 1: 38. https://doi.org/10.3390/antiox13010038