
Assessment of Bio-Compounds Content, Antioxidant Activity, and Neuroprotective Effect of Red Cabbage (Brassica oleracea var. Capitata rubra) Processed by Convective Drying at Different Temperatures

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solvents and Reagents
2.2. Raw Material and Drying Conditions
2.3. Determination of Bioactive Compounds
2.3.1. Total Polyphenolics, Total Flavonoids, Total Anthocyanins and Total Glucosinolates
2.3.2. Extraction, Identification and Quantification of Phenolic Compounds
2.3.3. Amino Acid and Fatty Acid Profiles
2.4. Antioxidant Activity-DPPH and ORAC Assays
2.5. Neuroprotective Potential
2.6. Statistical Analysis
3. Results
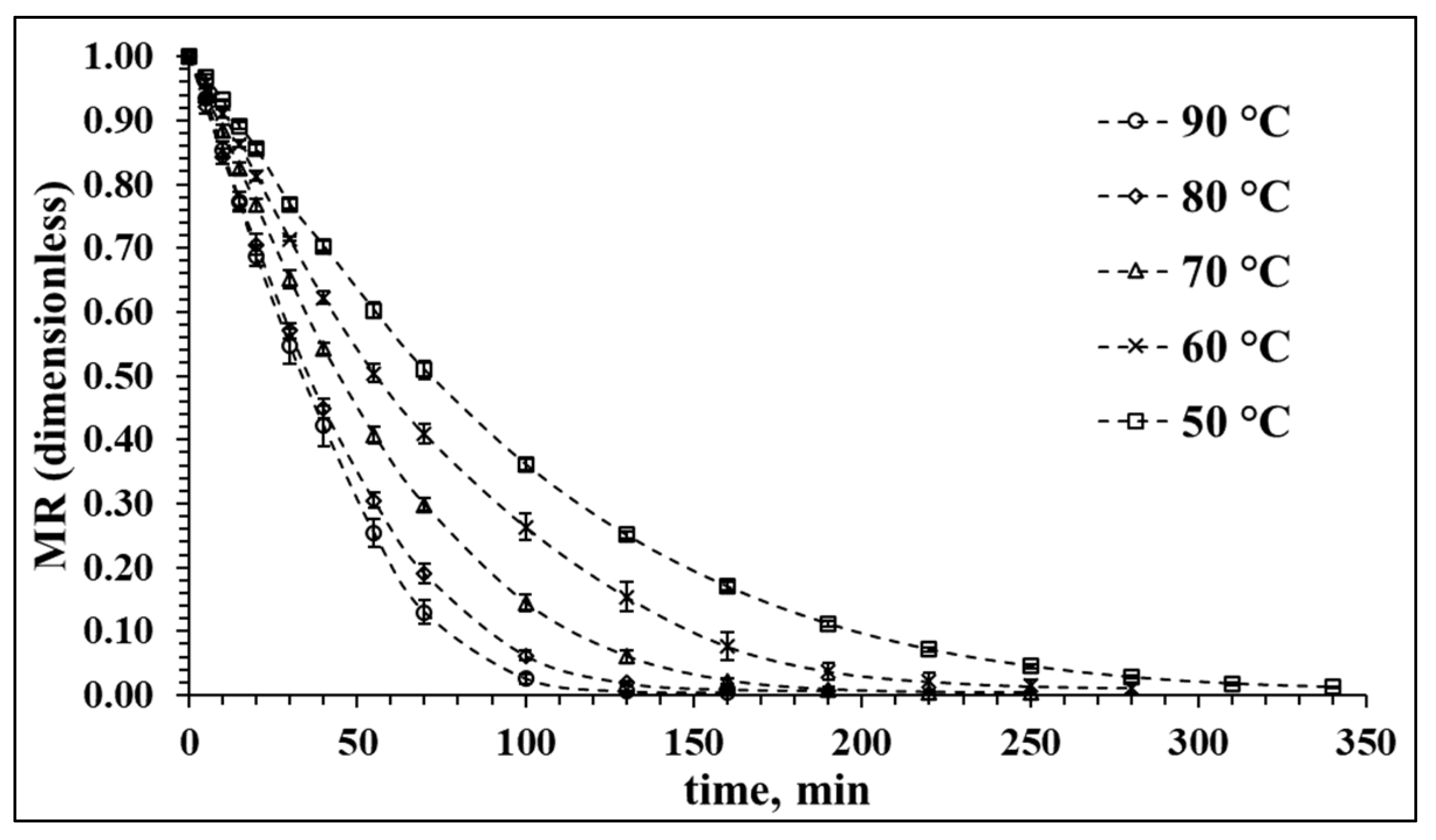
3.1. Drying Characteristics of Red Cabbage in Hot Air at Different Temperatures
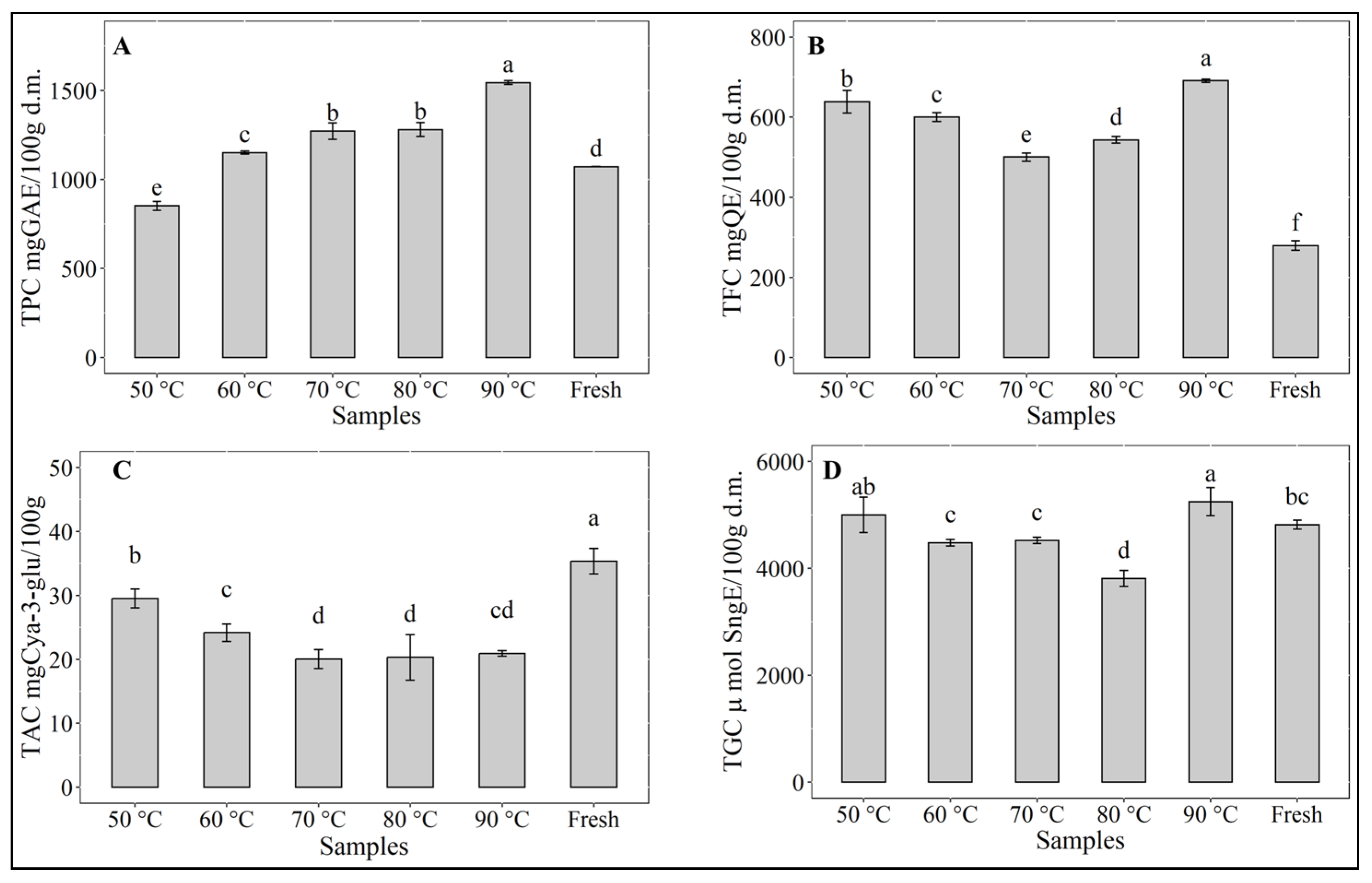
3.2. Bioactive Compounds
3.3. Identification and Quantification of Phenolic Compounds
3.4. Amino Acids and Fatty Acids Profiles
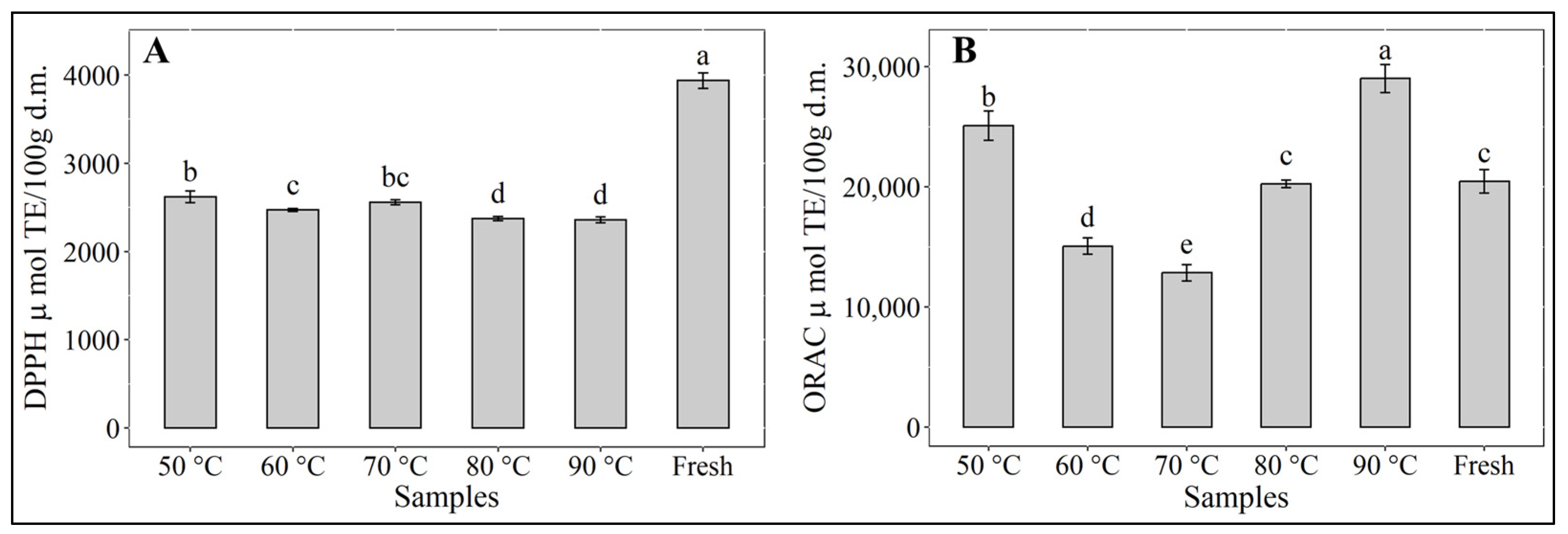
3.5. Antioxidant Activities
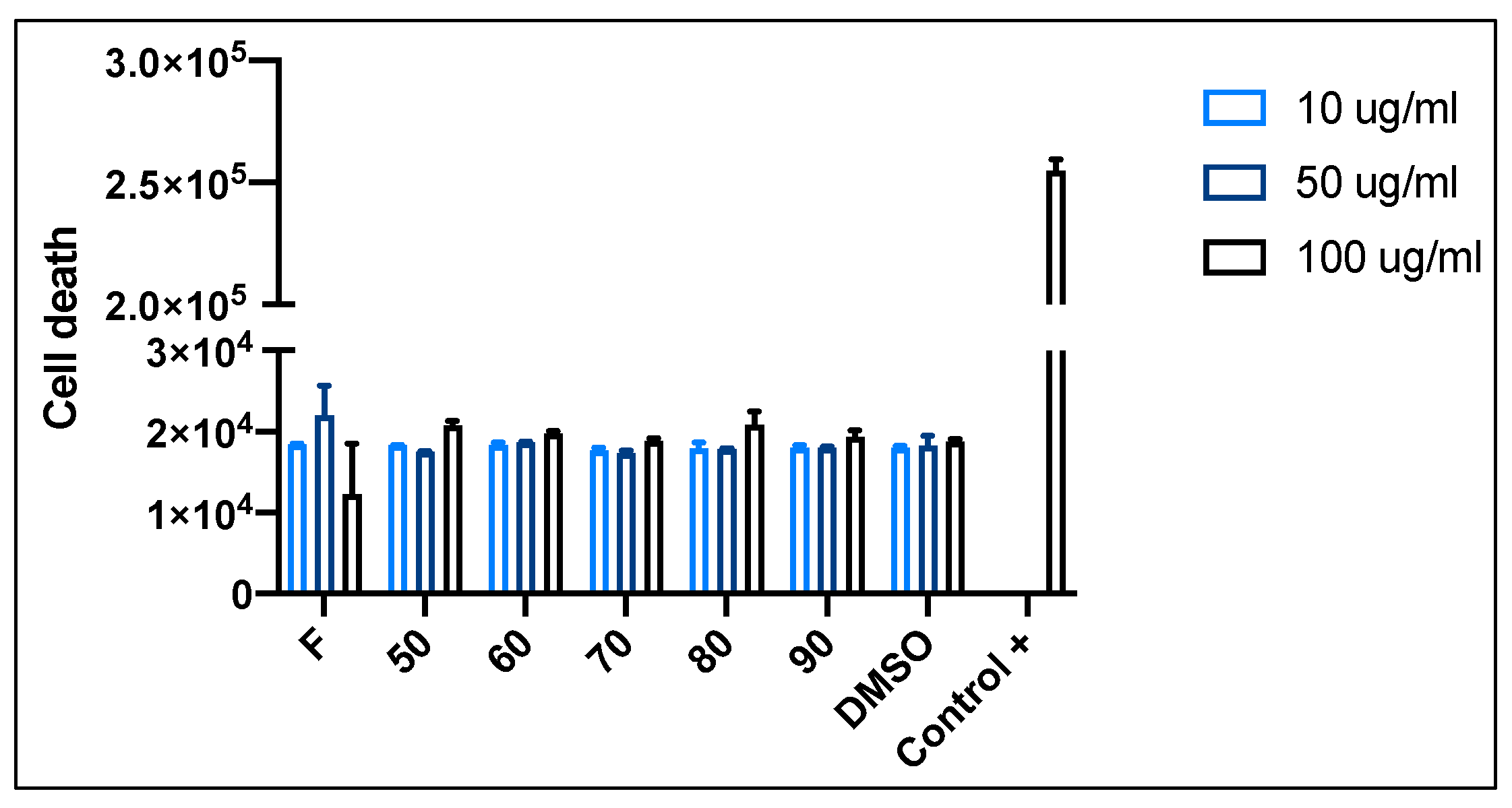
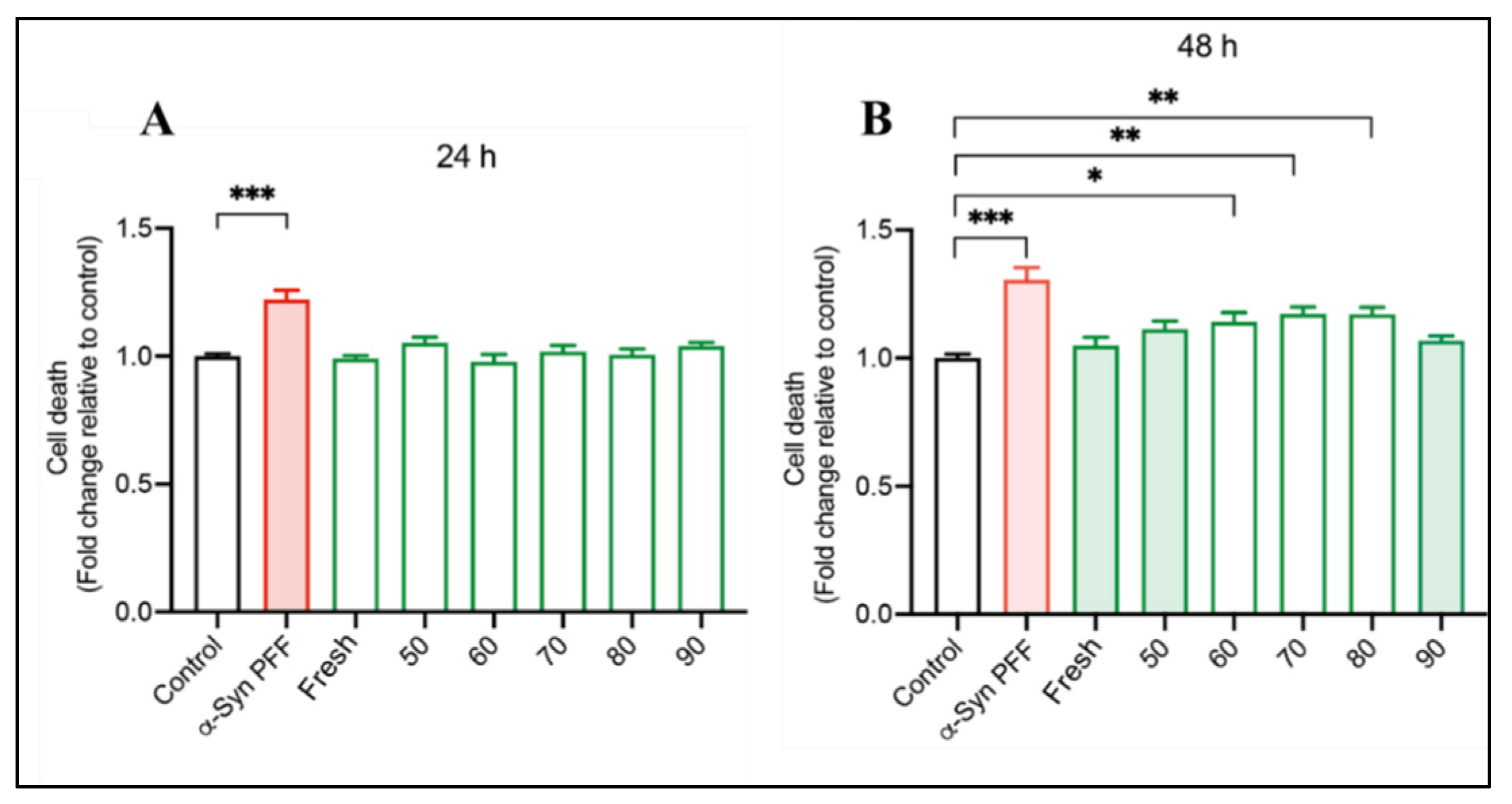
3.6. Neuroprotective Potential
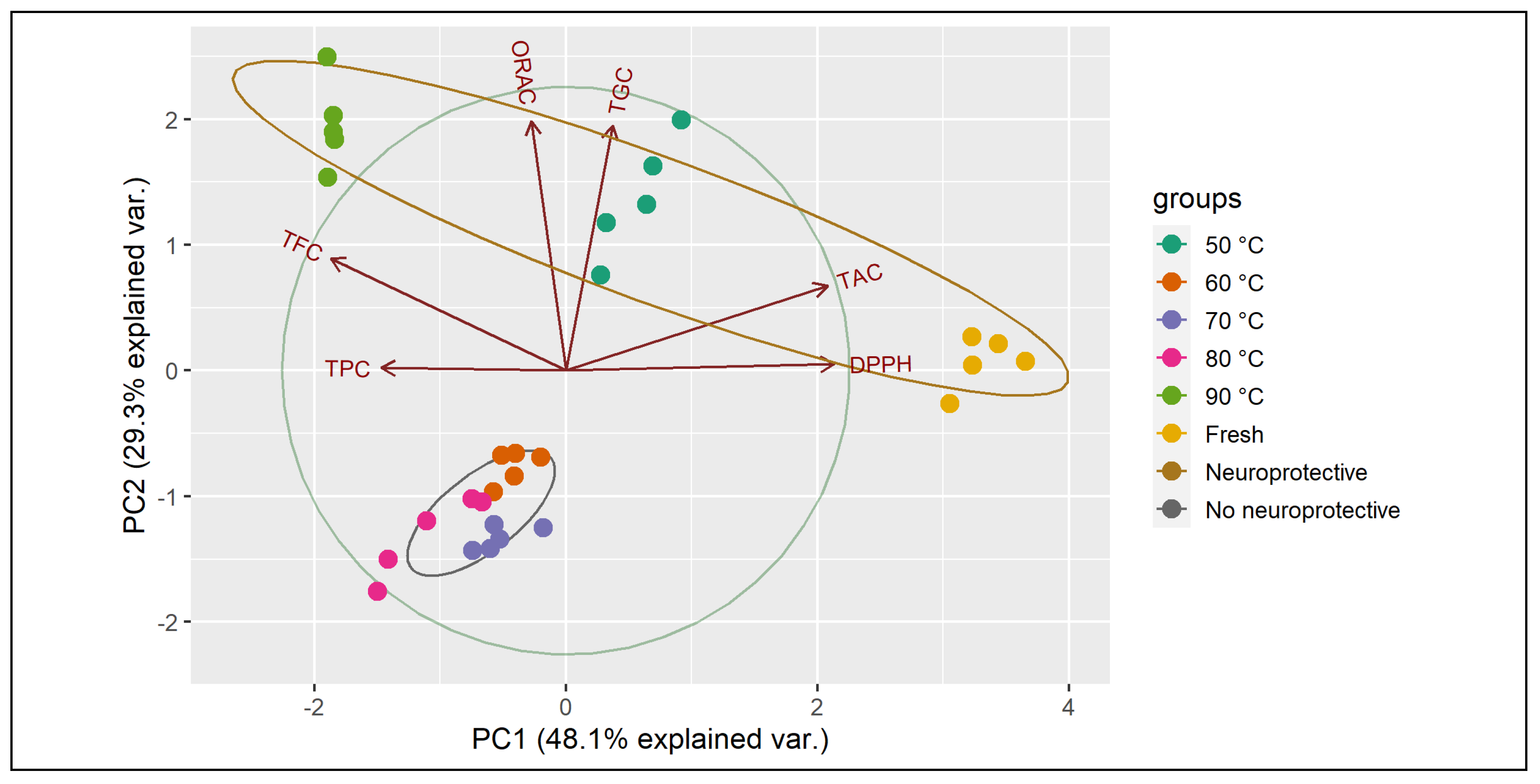
3.7. Principal Components Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ben Ticha, M.; Haddar, W.; Meksi, N.; Guesmi, A.; Mhenni, M.F. Improving Dyeability of Modified Cotton Fabrics by the Natural Aqueous Extract from Red Cabbage Using Ultrasonic Energy. Carbohydr. Polym. 2016, 154, 287–295. [Google Scholar] [CrossRef]
- Drozdowska, M.; Leszczyńska, T.; Koronowicz, A.; Piasna-Słupecka, E.; Domagała, D.; Kusznierewicz, B. Young Shoots of Red Cabbage Are a Better Source of Selected Nutrients and Glucosinolates in Comparison to the Vegetable at Full Maturity. Eur. Food Res. Technol. 2020, 246, 2505–2515. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wallig, M.A.; Jeffery, E.H. Dietary Broccoli Lessens Development of Fatty Liver and Liver Cancer in Mice given Diethylnitrosamine and Fed a Western or Control Diet. J. Nutr. 2016, 146, 542–550. [Google Scholar] [CrossRef]
- Kolonel, L.N.; Hankin, J.H.; Whittemore, A.S.; Wu, A.H.; Gallagher, R.P.; Wilkens, L.R.; John, E.M.; Howe, G.R.; Dreon, D.M.; West, D.W.; et al. Vegetables, Fruits, Legumes and Prostate Cancer: A Multiethnic Case-Control Study. Cancer Epidemiol. Biomarkers Prev. 2000, 9, 795–804. [Google Scholar] [PubMed]
- Veeranki, O.L.; Bhattacharya, A.; Tang, L.; Marshall, J.R.; Zhang, Y. Cruciferous Vegetables, Isothiocyanates, and Prevention of Bladder Cancer. Curr. Pharmacol. Rep. 2015, 1, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Schreiner, M. Isothiocyanates, Nitriles, and Epithionitriles from Glucosinolates Are Affected by Genotype and Developmental Stage in Brassica Oleracea Varieties. Front. Plant Sci. 2017, 8, 1095. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Yuan, J.M.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, 1700965. [Google Scholar] [CrossRef]
- Sankhari, J.M.; Thounaojam, M.C.; Jadeja, R.N.; Devkar, R.V.; Ramachandran, A.V. Anthocyanin-Rich Red Cabbage (Brassica Oleracea L.) Extract Attenuates Cardiac and Hepatic Oxidative Stress in Rats Fed an Atherogenic Diet. J. Sci. Food Agric. 2012, 92, 1688–1693. [Google Scholar] [CrossRef]
- Zielinska, M.; Lewandowska, U.; Podsedek, A.; Cygankiewicz, A.; Jacenik, D.; Sałaga, M.; Kordek, R.; Krajewska, W.; Fichna, J. Orally Available Extract from Brassica Oleracea Var. Capitata Rubra Attenuates Experimental Colitis in Mouse Models of Inflammatory Bowel Diseases. J. Funct. Foods 2015, 17, 587–599. [Google Scholar] [CrossRef]
- Abdul-Latif, R.; Stupans, I.; Allahham, A.; Adhikari, B.; Thrimawithana, T. Natural Antioxidants in the Management of Parkinson’s Disease: Review of Evidence from Cell Line and Animal Models. J. Integr. Med. 2021, 19, 300–310. [Google Scholar] [CrossRef]
- Zaltieri, M.; Longhena, F.; Pizzi, M.; Missale, C.; Spano, P.; Bellucci, A. Mitochondrial Dysfunction and Alpha-Synuclein Synaptic Pathology in Parkinson’s Disease: Who’s on First? Parkinsons. Dis. 2015, 2015, 108029. [Google Scholar] [PubMed]
- Yang, F.; Wolk, A.; Håkansson, N.; Pedersen, N.L.; Wirdefeldt, K. Dietary Antioxidants and Risk of Parkinson’s Disease in Two Population-Based Cohorts. Mov. Disord. 2017, 32, 1631–1636. [Google Scholar] [CrossRef]
- Parasram, K. Phytochemical Treatments Target Kynurenine Pathway Induced Oxidative Stress. Redox Rep. 2018, 23, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S.; Rajasankar, S. Parkinson’s Disease: Oxidative Stress and Therapeutic Approaches. Neurol. Sci. 2010, 31, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Bezard, E.; Zhao, B. Protective Effect of Green Tea Polyphenols on the SH-SY5Y Cells against 6-OHDA Induced Apoptosis through ROS-NO Pathway. Free Radic. Biol. Med. 2005, 39, 682–695. [Google Scholar] [CrossRef]
- Cho, H.S.; Kim, S.; Lee, S.Y.; Park, J.A.; Kim, S.J.; Chun, H.S. Protective Effect of the Green Tea Component, l-Theanine on Environmental Toxins-Induced Neuronal Cell Death. Neurotoxicology 2008, 29, 656–662. [Google Scholar] [CrossRef]
- Jaisin, Y.; Thampithak, A.; Meesarapee, B.; Ratanachamnong, P.; Suksamrarn, A.; Phivthong-ngam, L.; Phumala-Morales, N.; Chongthammakun, S.; Govitrapong, P.; Sanvarinda, Y. Curcumin I Protects the Dopaminergic Cell Line SH-SY5Y from 6-Hydroxydopamine-Induced Neurotoxicity through Attenuation of P53-Mediated Apoptosis. Neurosci. Lett. 2011, 489, 192–196. [Google Scholar] [CrossRef]
- Moosavi, M.; Owjfard, M.; Farokhi, M.R. Curcumin Prevents 6-Ohda Induced Cell Death and Erk Disruption in Human Neuroblastoma Cells. J. Knowl. Health Basic Med. Sci. 2018, 13, 1–7. [Google Scholar] [CrossRef]
- Oboh, G.; Ademiluyi, A.O.; Ogunsuyi, O.B.; Oyeleye, S.I.; Dada, A.F.; Boligon, A.A. Cabbage and Cucumber Extracts Exhibited Anticholinesterase, Antimonoamine Oxidase and Antioxidant Properties. J. Food Biochem. 2017, 41, e12358. [Google Scholar] [CrossRef]
- Ghareaghajlou, N.; Hallaj-Nezhadi, S.; Ghasempour, Z. Red Cabbage Anthocyanins: Stability, Extraction, Biological Activities and Applications in Food Systems. Food Chem. 2021, 365, 130482. [Google Scholar] [CrossRef]
- Mori, M.A.; Delattre, A.M.; Carabelli, B.; Pudell, C.; Bortolanza, M.; Staziaki, P.V.; Visentainer, J.V.; Montanher, P.F.; Del Bel, E.A.; Ferraz, A.C. Neuroprotective Effect of Omega-3 Polyunsaturated Fatty Acids in the 6-OHDA Model of Parkinson’s Disease Is Mediated by a Reduction of Inducible Nitric Oxide Synthase. Nutr. Neurosci. 2017, 21, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Sakulnarmrat, K.; Wongsrikaew, D.; Konczak, I. Microencapsulation of Red Cabbage Anthocyanin-Rich Extract by Drum Drying Technique. Lwt 2021, 137, 110473. [Google Scholar] [CrossRef]
- Song, C.F.; Cui, Z.W.; Jin, G.Y.; Mujumdar, A.S.; Yu, J.F. Effects of Four Different Drying Methods on the Quality Characteristics of Peeled Litchis (Litchi Chinensis Sonn.). Dry. Technol. 2015, 33, 583–590. [Google Scholar] [CrossRef]
- Tao, Y.; Han, M.; Gao, X.; Han, Y.; Show, P.L.; Liu, C.; Ye, X.; Xie, G. Applications of Water Blanching, Surface Contacting Ultrasound-Assisted Air Drying, and Their Combination for Dehydration of White Cabbage: Drying Mechanism, Bioactive Profile, Color and Rehydration Property. Ultrason. Sonochem. 2019, 53, 192–201. [Google Scholar] [CrossRef]
- Gómez-Pérez, L.S.; Navarrete, C.; Moraga, N.; Rodríguez, A.; Vega-Gálvez, A. Evaluation of Different Hydrocolloids and Drying Temperatures in the Drying Kinetics, Modeling, Color, and Texture Profile of Murta (Ugni Molinae Turcz) Berry Leather. J. Food Process Eng. 2020, 43, e13316. [Google Scholar] [CrossRef]
- Ke, Y.Y.; Shyu, Y.T.; Wu, S.J. Evaluating the Anti-Inflammatory and Antioxidant Effects of Broccoli Treated with High Hydrostatic Pressure in Cell Models. Foods 2021, 10, 167. [Google Scholar] [CrossRef]
- Uribe, E.; Gómez-Pérez, L.S.; Pasten, A.; Pardo, C.; Puente, L.; Vega-Galvez, A. Assessment of Refractive Window Drying of Physalis (Physalis Peruviana L.) Puree at Different Temperatures: Drying Kinetic Prediction and Retention of Bioactive Components. J. Food Meas. Charact. 2022, 16, 2605–2615. [Google Scholar] [CrossRef]
- Dini, I.; Tenore, G.C.; Dini, A. Antioxidant Compound Contents and Antioxidant Activity before and after Cooking in Sweet and Bitter Chenopodium Quinoa Seeds. LWT—Food Sci. Technol. 2010, 43, 447–451. [Google Scholar] [CrossRef]
- De Souza, V.R.; Pereira, P.A.P.; Da Silva, T.L.T.; De Oliveira Lima, L.C.; Pio, R.; Queiroz, F. Determination of the Bioactive Compounds, Antioxidant Activity and Chemical Composition of Brazilian Blackberry, Red Raspberry, Strawberry, Blueberry and Sweet Cherry Fruits. Food Chem. 2014, 156, 362–368. [Google Scholar] [CrossRef]
- Aghajanzadeh, T.; Hawkesford, M.J.; De Kok, L.J. The Significance of Glucosinolates for Sulfur Storage in Brassicaceae Seedlings. Front. Plant Sci. 2014, 5, 704. [Google Scholar] [CrossRef]
- Araya, M.; García, S.; Rengel, J.; Pizarro, S.; Álvarez, G. Determination of Free and Protein Amino Acid Content in Microalgae by HPLC-DAD with Pre-Column Derivatization and Pressure Hydrolysis. Mar. Chem. 2021, 234, 103999. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane, G. A Simple Method for the Isolation and Purificationof Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Hewavitharana, G.G.; Perera, D.N.; Navaratne, S.B.; Wickramasinghe, I. Extraction Methods of Fat from Food Samples and Preparation of Fatty Acid Methyl Esters for Gas Chromatography: A Review. Arab. J. Chem. 2020, 13, 6865–6875. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Uribe, E.; Lemus-Mondaca, R.; Vega-Gálvez, A.; Zamorano, M.; Quispe-Fuentes, I.; Pasten, A.; Di Scala, K. Influence of Process Temperature on Drying Kinetics, Physicochemical Properties and Antioxidant Capacity of the Olive-Waste Cake. Food Chem. 2014, 147, 170–176. [Google Scholar] [CrossRef]
- Vega-Galvez, A.; Uribe, E.; Pasten, A.; Camus, J.; Gomez-Perez, L.S.; Mejias, N.; Vidal, R.L.; Grunenwald, F.; Aguilera, L.E.; Valenzuela-Barra, G. Comprehensive Evaluation of the Bioactive Composition and Neuroprotective and Antimicrobial Properties of Vacuum-Dried Broccoli (Brassica Oleracea Var. Italica) Powder and Its Antioxidants. Molecules 2023, 28, 766. [Google Scholar] [CrossRef]
- Puente-Díaz, L.; Ah-Hen, K.; Vega-Gálvez, A.; Lemus-Mondaca, R.; Di Scala, K. Combined Infrared-Convective Drying of Murta (Ugni Molinae Turcz) Berries: Kinetic Modeling and Quality Assessment. Dry. Technol. 2013, 31, 329–338. [Google Scholar] [CrossRef]
- Saifullah, M.; McCullum, R.; McCluskey, A.; Vuong, Q. Effects of Different Drying Methods on Extractable Phenolic Compounds and Antioxidant Properties from Lemon Myrtle Dried Leaves. Heliyon 2019, 5, e03044. [Google Scholar] [CrossRef]
- Vu, H.T.; Scarlett, C.J.; Vuong, Q.V. Optimization of Ultrasound-Assisted Extraction Conditions for Recovery of Phenolic Compounds and Antioxidant Capacity from Banana (Musa Cavendish) Peel. J. Food Process. Preserv. 2017, 41, e13148. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Murtijaya, J. Antioxidant Properties of Phyllanthus Amarus Extracts as Affected by Different Drying Methods. LWT—Food Sci. Technol. 2007, 40, 1664–1669. [Google Scholar] [CrossRef]
- Martín-Cabrejas, M.A.; Aguilera, Y.; Pedrosa, M.M.; Cuadrado, C.; Hernández, T.; Díaz, S.; Esteban, R.M. The Impact of Dehydration Process on Antinutrients and Protein Digestibility of Some Legume Flours. Food Chem. 2009, 114, 1063–1068. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; Vargas, N.; Pasten, A.; Rodríguez, K.; Ah-Hen, K.S. Phytochemical Components and Amino Acid Profile of Brown Seaweed Durvillaea Antarctica as Affected by Air Drying Temperature. J. Food Sci. Technol. 2018, 55, 4792–4801. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, Y.; Corke, H.; Zhu, H. Dynamic Changes in Flavonoids Content during Congou Black Tea Processing. LWT 2022, 170, 114073. [Google Scholar] [CrossRef]
- Geng, Z.; Wang, J.; Zhu, L.; Yu, X.; Zhang, Q.; Li, M.; Hu, B.; Yang, X. Metabolomics Provide a Novel Interpretation of the Changes in Flavonoids during Sea Buckthorn (Hippophae Rhamnoides L.) Drying. Food Chem. 2023, 413, 135598. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Zhao, Y.; Li, L.; He, Y.; Zhang, X.; Zhu, Y.; Hong, G. Comparative Metabolomics of Flavonoids in Twenty Vegetables Reveal Their Nutritional Diversity and Potential Health Benefits. Food Res. Int. 2023, 164, 112384. [Google Scholar] [CrossRef]
- Orzechowska, B.U.; Wróbel, G.; Turlej, E.; Jatczak, B.; Sochocka, M.; Chaber, R. Antitumor Effect of Baicalin from the Scutellaria Baicalensis Radix Extract in B-Acute Lymphoblastic Leukemia with Different Chromosomal Rearrangements. Int. Immunopharmacol. 2020, 79, 106114. [Google Scholar] [CrossRef]
- Fan, X.; Fan, Z.; Yang, Z.; Huang, T.; Tong, Y.; Yang, D.; Mao, X.; Yang, M. Flavonoids—Natural Gifts to Promote Health and Longevity. Int. J. Mol. Sci. 2022, 23, 2176. [Google Scholar] [CrossRef]
- Mitra, S.; Lami, M.S.; Uddin, T.M.; Das, R.; Islam, F.; Anjum, J.; Hossain, M.J.; Emran, T. Bin Prospective Multifunctional Roles and Pharmacological Potential of Dietary Flavonoid Narirutin. Biomed. Pharmacother. 2022, 150, 112932. [Google Scholar] [CrossRef]
- Hosseini, S.; Gharachorloo, M.; Ghiassi-Tarzi, B.; Ghavami, M. Evaluation of the Organic Acids Ability for Extraction of Anthocyanins and Phenolic Compounds from Different Sources and Their Degradation Kinetics during Cold Storage. Polish J. Food Nutr. Sci. 2016, 66, 261–269. [Google Scholar] [CrossRef]
- Podsȩdek, A.; Majewska, I.; Kucharska, A.Z. Inhibitory Potential of Red Cabbage against Digestive Enzymes Linked to Obesity and Type 2 Diabetes. J. Agric. Food Chem. 2017, 65, 7192–7199. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Functional Properties of Anthocyanins and Betalains in Plants, Food, and in Human Nutrition. Trends Food Sci. Technol. 2004, 15, 19–38. [Google Scholar] [CrossRef]
- Buchweitz, M.; Brauch, J.; Carle, R.; Kammerer, D.R. Colour and Stability Assessment of Blue Ferric Anthocyanin Chelates in Liquid Pectin-Stabilised Model Systems. Food Chem. 2013, 138, 2026–2035. [Google Scholar] [CrossRef] [PubMed]
- Dyrby, M.; Westergaard, N.; Stapelfeldt, H. Light and Heat Sensitivity of Red Cabbage Extract in Soft Drink Model Systems. Food Chem. 2001, 72, 431–437. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Jampani, C.; Raghavarao, K.S.M.S. Process Integration for Purification and Concentration of Red Cabbage (Brassica oleracea L.) Anthocyanins. Sep. Purif. Technol. 2015, 141, 10–16. [Google Scholar] [CrossRef]
- Ekici, L.; Simsek, Z.; Ozturk, I.; Sagdic, O.; Yetim, H. Effects of Temperature, Time, and PH on the Stability of Anthocyanin Extracts: Prediction of Total Anthocyanin Content Using Nonlinear Models. Food Anal. Methods 2014, 7, 1328–1336. [Google Scholar] [CrossRef]
- Ikeura, H.; Kobayashi, F.; Hayata, Y. Optimum Extraction Method for Volatile Attractant Compounds in Cabbage to Pieris Rapae. Biochem. Syst. Ecol. 2012, 40, 201–207. [Google Scholar] [CrossRef]
- Oerlemans, K.; Barrett, D.M.; Suades, C.B.; Verkerk, R.; Dekker, M. Thermal Degradation of Glucosinolates in Red Cabbage. Food Chem. 2006, 95, 19–29. [Google Scholar] [CrossRef]
- Kaulmann, A.; Jonville, M.C.; Schneider, Y.J.; Hoffmann, L.; Bohn, T. Carotenoids, Polyphenols and Micronutrient Profiles of Brassica Oleraceae and Plum Varieties and Their Contribution to Measures of Total Antioxidant Capacity. Food Chem. 2014, 155, 240–250. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Wiczkowski, W.; Płatosz, N.; Bartoszek, A. Phytochemical Composition and Biological Activities of Differently Pigmented Cabbage (Brassica Oleracea Var. Capitata) and Cauliflower (Brassica Oleracea Var. Botrytis) Varieties. J. Sci. Food Agric. 2019, 99, 5499–5507. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, Y.; Xu, S.; Xu, W.; Chu, K.; Xu, W.; Zhao, H.; Lu, J. Identification and Quantification of Phenolic Compounds in Vitex Negundo L. Var. Cannabifolia (Siebold et Zucc.) Hand.-Mazz. Using Liquid Chromatography Combined with Quadrupole Time-of-Flight and Triple Quadrupole Mass Spectrometers. J. Pharm. Biomed. Anal. 2015, 108, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Radünz, M.; Hackbart, H.C.D.S.; Bona, N.P.; Pedra, N.S.; Hoffmann, J.F.; Stefanello, F.M.; Da Rosa Zavareze, E. Glucosinolates and Phenolic Compounds Rich Broccoli Extract: Encapsulation by Electrospraying and Antitumor Activity against Glial Tumor Cells. Colloids Surf. B Biointerfaces 2020, 192, 111020. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional Amino Acids in Growth, Reproduction, and Health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, Y.; Yue, J.; Liu, Z.; Zheng, Y.; Qian, B.; Zhong, Y.; Zhao, Y. Thermal Behavior, Microstructure and Protein Quality of Squid Fillets Dried by Far-Infrared Assisted Heat Pump Drying. Food Control 2014, 36, 102–110. [Google Scholar] [CrossRef]
- Zhao, Y.; Jiang, Y.; Zheng, B.; Zhuang, W.; Zheng, Y.; Tian, Y. Influence of Microwave Vacuum Drying on Glass Transition Temperature, Gelatinization Temperature, Physical and Chemical Qualities of Lotus Seeds. Food Chem. 2017, 228, 167–176. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, L.; Kranjac, M.; Marijanovic, Z.; Jerkovic, I.; Corell, M.; Moriana, A.; Carbonell-Barrachina, Á.A.; Sendra, E.; Hernández, F. Quality Attributes and Fatty Acid, Volatile and Sensory Profiles of “Arbequina” HydroSOStainable Olive Oil. Molecules 2019, 24, 2148. [Google Scholar] [CrossRef]
- Dowd, M.K. Identification of the Unsaturated Heptadecyl Fatty Acids in the Seed Oils of Thespesia Populnea and Gossypium Hirsutum. J. Am. Oil Chem. Soc. 2012, 89, 1599–1609. [Google Scholar] [CrossRef]
- Mensink, R.P. Effects of Saturated Fatty Acids on Serum Lipids and Lipoproteins: A Systematic Review and Regression Analysis; World Health Organization: Geneva, Switzerland, 2016; p. 63. [Google Scholar]
- Avallone, R.; Vitale, G.; Bertolotti, M. Omega-3 Fatty Acids and Neurodegenerative Diseases: New Evidence in Clinical Trials. Int. J. Mol. Sci. 2019, 20, 4256. [Google Scholar] [CrossRef]
- Pan, A.; Chen, M.; Chowdhury, R.; Wu, J.H.Y.; Sun, Q.; Campos, H.; Mozaffarian, D.; Hu, F.B. α-Linolenic Acid and Risk of Cardiovascular Disease: A Systematic Review and Meta-Analysis. Am. J. Clin. Nutr. 2012, 96, 1262–1273. [Google Scholar] [CrossRef]
- Tu, T.H.; Kim, H.; Yang, S.; Kim, J.K.; Kim, J.G. Linoleic Acid Rescues Microglia Inflammation Triggered by Saturated Fatty Acid. Biochem. Biophys. Res. Commun. 2019, 513, 201–206. [Google Scholar] [CrossRef]
- Ali, W.; Ikram, M.; Park, H.Y.; Jo, M.G.; Ullah, R.; Ahmad, S.; Abid, N.-B.; Kim, M.O. Oral Administration of Alpha Linoleic Acid Rescues Aβ-Induced Glia-Mediated Neuroinflammation and Cognitive Dysfunction in C57BL/6N Mice. Cells 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Qiu, B.; Jia, M.; Liu, W.; Guo, X.-F.; Li, N.; Xu, Z.-X.; Du, F.-L.; Xu, T.; Li, D. Effects of α-Linolenic Acid Intake on Blood Lipid Profiles: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Crit. Rev. Food Sci. Nutr. 2020, 61, 2894–2910. [Google Scholar] [CrossRef] [PubMed]
- López, J.; Vega-Gálvez, A.; Bilbao-Sainz, C.; Uribe, E.; Chiou, B.-S.; Quispe-Puentes, I. Influence of Vacuum Drying Temperature on: Physico-Chemical Composition and Antioxidant Properties of Murta Berries. Food Process Eng. 2017, 40, e12569. [Google Scholar] [CrossRef]
- Chaves, N.; Santiago, A.; Alías, J.C. Quantification of the Antioxidant Activity of Plant Extracts: Analysis of Sensitivity and Hierarchization Based on the Method Used. Antioxidants 2020, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xiao, Y.; Lagnika, C.; Li, D.; Liu, C.; Jiang, N.; Song, J.; Zhang, M. A Comparative Evaluation of Nutritional Properties, Antioxidant Capacity and Physical Characteristics of Cabbage (Brassica Oleracea Var. Capitate Var L.) Subjected to Different Drying Methods. Food Chem. 2020, 309, 124935. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Ellis, A. Dietary Antioxidants and Parkinson Disease. Antioxidants 2020, 9, 570. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Aryal, S.; Skinner, T.; Bridges, B.; Weber, J.T. The Pathology of Parkinson’s Disease and Potential Benefit of Dietary Polyphenols. Molecules 2020, 25, 4382. [Google Scholar] [CrossRef]
- Zhao, D.; Simon, J.E.; Wu, Q. A Critical Review on Grape Polyphenols for Neuroprotection: Strategies to Enhance Bioefficacy. Crit. Rev. Food Sci. Nutr. 2020, 60, 597–625. [Google Scholar] [CrossRef]
- Khalatbary, A.R.; Khademi, E. The Green Tea Polyphenolic Catechin Epigallocatechin Gallate and Neuroprotection. Nutr. Neurosci. 2020, 23, 281–294. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Zhang, X.; Jin, Q.; Li, J. Phenolic Profiles, Antioxidant Activities, and Neuroprotective Properties of Mulberry (Morus Atropurpurea Roxb.) Fruit Extracts from Different Ripening Stages. J. Food Sci. 2016, 81, C2439–C2446. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, D.Y.; Jeong, R.H.; Lee, D.S.; Kim, Y.E.; Hong, E.K.; Kim, Y.C.; Baek, N.I. Neuroprotective Effect of Prenylated Arylbenzofuran and Flavonoids from Morus Alba Fruits on Glutamate-Induced Oxidative Injury in HT22 Hippocampal Cells. J. Med. Food 2015, 18, 403–408. [Google Scholar] [CrossRef]
- Cirmi, S.; Maugeri, A.; Lombardo, G.E.; Russo, C.; Musumeci, L.; Gangemi, S.; Calapai, G.; Barreca, D.; Navarra, M. A Flavonoid-Rich Extract of Mandarin Juice Counteracts 6-Ohda-Induced Oxidative Stress in Sh-Sy5y Cells and Modulates Parkinson-Related Genes. Antioxidants 2021, 10, 539. [Google Scholar] [CrossRef]
- Neves, D.; Valentão, P.; Bernardo, J.; Oliveira, M.C.; Ferreira, J.M.G.; Pereira, D.M.; Andrade, P.B.; Videira, R.A. A New Insight on Elderberry Anthocyanins Bioactivity: Modulation of Mitochondrial Redox Chain Functionality and Cell Redox State. J. Funct. Foods 2019, 56, 145–155. [Google Scholar] [CrossRef]
- Ni, J.; Wu, Z.; Meng, J.; Zhu, A.; Zhong, X.; Wu, S.; Nakanishi, H. The Neuroprotective Effects of Brazilian Green Propolis on Neurodegenerative Damage in Human Neuronal SH-SY5Y Cells. Oxid. Med. Cell. Longev. 2017, 2017, 7984327. [Google Scholar] [CrossRef] [PubMed]
- Zaim, M.; Kara, I.; Muduroglu, A. Black Carrot Anthocyanins Exhibit Neuroprotective Effects against MPP+ Induced Cell Death and Cytotoxicity via Inhibition of Oxidative Stress Mediated Apoptosis. Cytotechnology 2021, 73, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Pakrashi, S.; Chakraborty, J.; Bandyopadhyay, J. Neuroprotective Role of Quercetin on Rotenone-Induced Toxicity in SH-SY5Y Cell Line Through Modulation of Apoptotic and Autophagic Pathways. Neurochem. Res. 2020, 45, 1962–1973. [Google Scholar] [CrossRef] [PubMed]
- Natella, F.; Maldini, M.; Leoni, G.; Scaccini, C. Glucosinolates Redox Activities: Can They Act as Antioxidants? Food Chem. 2014, 149, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Hurtado, F.; Gicquel, M.; Esnault, M.A. Evaluation of the Antioxidant Potential of Cauliflower (Brassica Oleracea) from a Glucosinolate Content Perspective. Food Chem. 2012, 132, 1003–1009. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal Component Analysis: A Review and Recent Developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | [M − H]− Theoretical Mass | [M − H]− Experimental Mass | Error ppm | MS/MS (m/z) | Tentative Identification | Abbreviation | |
|---|---|---|---|---|---|---|---|
| 1 | 353.0878 | 353.0880 | +0.55 | 191.0553–135.0440–179.0341 | Neochlorogenic acid | NChla | |
| 2 | 477.0674 | 477.0642 | −6.92 | 259.0129–96.9588 | Quercetin 3′-glucuronide | Q3G | |
| 3 | 193.0506 | 193.0499 | −3.62 | Ferulic acid | FA | ||
| 4 | 353.0878 | 353.0879 | +0.28 | 135.0441–191.0552–173.0445 | Chlorogenic acid | Chla | |
| 5 | unknown | 535.0223 | 336.9931–477.0638 | Quercetin unknown derivate | QUd | ||
| 6 | 179.0350 | 179.0341 | −5.03 | 135.0443 | Caffeic acid | CA | |
| 7 | 477.0675 | 477.0645 | −6.21 | 468.0269–96.9588 | Quercetin 4’-glucuronide | Q4G | |
| 8 | 223.0612 | 223.0608 | −1.79 | Sinapic acid | SA | ||
| 9 | 609.1461 | 609.1421 | −6.56 | Rutin | R | ||
| Tentative Compound | 50 °C | 60 °C | 70 °C | 80 °C | 90 °C | Control | |
| NChla | 24.54 ± 3.69 ab | 0.63 ± 0.11 d | 8.83 ± 0.11 c | 30.78 ± 5.57 a | 24.04 ± 1.26 ab | 19.47 ± 1.83 b | |
| Q3G | 16.09 ± 0.27 b | 14.36 ± 3.15 b | 12.81 ± 0.34 bc | 9.07 ± 2.84 c | 12.76 ± 0.06 bc | 26.52 ± 1.92 a | |
| FA | <LOQ | <LOQ | ND | <LOQ | <LOQ | ND | |
| Chla | 11.44 ± 0.96 a | 0.27 ± 0.05 b | ND | 12.51 ± 6.19 a | ND | ND | |
| QUd | 1.43 ± 0.10 a | 0.99 ± 0.53 abc | 0.44 ± 0.02 bc | 0.47 ± 0.41 bc | 0.30 ± 0.03 c | 1.12 ±0.01 ab | |
| CA | 1.23 ± 0.06 b | 0.48 ± 0.07 c | 0.95 ± 0.09 b | 2.29 ± 0.35 a | 2.13 ± 0.06 a | 1.27 ± 0.04 b | |
| Q4Ge | 11.73 ± 1.33 a | 12.13 ± 0.62 a | ND | 3.40 ± 2.26 b | ND | ND | |
| SA | 0.43 ± 0.10 c | <LOQ | <LOQ | 1.60 ± 0.03 a | 0.77 ± 0.03 b | <LOQ | |
| R | 0.69 ± 0.18 d | 0.90 ± 0.17 cd | 1.62 ± 0.02 a | 0.76 ± 0.24 d | 1.22 ± 0.10 bc | 1.33 ± 0.06 ab | |
| Amino Acids g/100 g d.m. | Drying Temperature (°C) | ||||
|---|---|---|---|---|---|
| 50 | 60 | 70 | 80 | 90 | |
| ASP | 0.93 ± 0.06 c | 0.60 ± 0.27 c | 1.81 ± 0.33 b | 2.68 ± 0.45 a | 2.06 ± 0.36 ab |
| GLU | 2.26 ± 0.14 cd | 1.25 ± 0.50 d | 3.87 ± 0.69 bc | 6.42 ± 1.17 a | 4.61 ± 0.59 ab |
| SER | 0.50 ± 0.10 cd | 0.35 ± 0.15 d | 0.97 ± 0.20 bc | 1.54 ± 0.28 a | 1.25 ± 0.19 ab |
| GLY | 0.35 ± 0.19 a | 0.39 ± 0.08 a | 0.65 ± 0.42 a | 0.94 ± 0.26 a | 0.72 ± 0.04 a |
| THR | ND | ND | 0.69 ± 0.57 a | 1.35 ± 0.22 a | 1.02 ± 0.11 a |
| ARG | 0.61 ± 0.31 c | 0.23 ± 0.08 c | 3.04 ± 0.24 b | 3.75 ± 0.54 ab | 3.90 ± 0.09 a |
| ALA | 0.32 ± 0.04 b | 0.25 ± 0.11 b | 0.52 ± 0.11 b | 1.14 ± 0.23 a | 1.31 ± 0.06 a |
| TYR | 0.67 ± 0.23 abc | 0.44 ± 0.15 bc | 0.29 ± 0.15 c | 0.93 ± 0.08 a | 0.86 ± 0.15 ab |
| VAL | 0.42 ± 0.09 bc | 0.25 ± 0.08 c | 0.71 ± 0.13 ab | 1.06 ± 0.24 a | 0.86 ± 0.12 a |
| PHE | 0.36 ± 0.00 b | 0.35 ± 0.18 b | 0.61 ± 0.09 b | 1.81 ± 0.42 a | 1.38 ± 0.17 a |
| ILE | 0.71 ± 0.24 bc | 0.39 ± 0.23 c | 1.38 ± 0.31 ab | 2.05 ± 0.36 a | 1.44 ± 0.07 a |
| LEU | 0.05 ± 0.05 b | ND | 0.36 ± 0.05 a | 0.45 ± 0.15 a | 0.35 ± 0.05 a |
| LYS | 0.39 ± 0.10 b | 0.29 ± 0.19 b | 0.91 ± 0.15 a | 1.32 ± 0.18 a | 1.04 ± 0.18 a |
| g/100 g FAMES | Drying Temperature (°C) | ||||
|---|---|---|---|---|---|
| 50 | 60 | 70 | 80 | 90 | |
| SFA | |||||
| C12:0 | ND | 0.26 ± 0.08 a | 0.12 ± 0.00 b | 0.1 ± 0.01 b | 0.12 ± 0.01 b |
| C14:0 | ND | 0.34 ± 0.02 a | 0.23 ± 0.01 b | 0.19 ± 0.00 b | 0.2 ± 0.01 b |
| C15:0 | 0.5 ± 0.07 a | 0.58 ± 0.09 a | 0.55 ± 0.04 a | 0.45 ± 0.03 a | 0.51 ± 0.01 a |
| C16:0 | 18.86 ± 0.88 a | 19.52 ± 1.43 a | 17.47 ± 0.42 a | 20.02 ± 0.85 a | 17.43 ± 0.21 a |
| C17:0 | 0.87 ± 0.10 ab | 0.52 ± 0.08 c | 0.71 ± 0.07 bc | 0.98 ± 0.07 a | 0.99 ± 0.04 a |
| C18:0 | 4.16 ± 0.10 b | 5.95 ± 0.97 a | 3.97 ± 0.05 b | 3.14 ± 0.15 b | 3.63 ± 0.05 b |
| C20:0 | 0.5 ± 0.07 a | 0.81 ± 0.19 a | 0.77 ± 0.03 a | 0.56 ± 0.10 a | 0.74 ± 0.01 a |
| C22:0 | 0.31 ± 0.00 a | 0.54 ± 0.18 a | 0.39 ± 0.04 a | 0.4 ± 0.04 a | 0.44 ± 0.08 a |
| MFA | |||||
| C16:1 | 0.32 ± 0.09 a | 0.41 ± 0.14 a | 0.32 ± 0.05 a | 0.32 ± 0.03 a | 0.42 ± 0.02 a |
| C17:1 | 0.24 ± 0.08 a | 0.35 ± 0.13 a | 0.3 ± 0.06 a | 0.2 ± 0.09 a | 0.26 ± 0.01 a |
| C18:1n9c/C18:1n9t | 1.42 ± 0.39 b | 2.26 ± 0.33 a | 1.77 ± 0.21 ab | 0.7 ± 0.26 b | 1.07 ± 0.06 b |
| C20:1n9 | 0.11 ± 0.04 a | 0.34 ± 0.13 a | 0.21 ± 0.03 a | 0.11 ± 0.03 a | ND |
| C22:1n9 | ND | 0.11 ± 0.00 a | 0.06 ± 0.04 a | 0.06 ± 0.04 a | 0.09 ± 0.05 a |
| C24:1n9 | 0.51 ± 0.04 a | 0.78 ± 0.30 a | 0.75 ± 0.22 a | 0.56 ± 0.04 a | 0.42 ± 0.20 a |
| PFA | |||||
| C18:2n6c | 20.7 ± 0.39 ab | 19.38 ± 2.40 ab | 22.36 ± 0.14 a | 17.34 ± 0.18 b | 18.53 ± 0.12 b |
| C18:3n3 | 51.19 ± 1.09 a | 48.39 ± 1.08 ab | 48.76 ± 0.36 ab | 54.17 ± 0.43 a | 54.31 ± 0.42 a |
| C20:2 | 0.13 ± 0.05 a | 0.2 ± 0.03 a | 0.23 ± 0.03 a | 0.19 ± 0.05 a | 0.19 ± 0.03 a |
| C20:3n3 | 0.11 ± 0.03 b | 0.31 ± 0.10 a | 0.2 ± 0.02 ab | 0.2 ± 0.06 ab | 0.19 ± 0.03 ab |
| C22:6n3 | 0.27 ± 0.00 a | 0.65 ± 0.24 a | 0.81 ± 0.42 a | 0.39 ± 0.11 a | 0.48 ± 0.25 a |
| Total SFA | 24.99 ± 0.87 a | 28.52 ± 3.04 a | 24.22 ± 0.34 a | 25.84 ± 0.57 a | 24.07 ± 0.24 a |
| Total MFA | 2.61 ± 0.55 b | 4.25 ± 1.03 a | 3.41 ± 0.18 ab | 1.91 ± 0.36 b | 2.23 ± 0.16 b |
| Total PFA | 72.39 ± 1.41 ab | 68.93 ± 3.85 b | 72.36 ± 0.40 ab | 72.28 ± 0.40 ab | 73.7 ± 0.35 a |
| ORAC | DPPH | TPC | TFC | TGC | TAC | |
|---|---|---|---|---|---|---|
| ORAC | 1 | |||||
| DPPH | −0.04512324 | 1 | ||||
| TPC | 0.1454054 | −0.38457781 | 1 | |||
| TFC | 0.38914863 | −0.88279979 | 0.27005582 | 1 | ||
| TGC | 0.54154351 | 0.18675585 | −0.02073628 | 0.17229989 | 1 | |
| TAC | 0.15248469 | 0.82962777 | −0.67074275 | −0.57943842 | 0.35656316 | 1 |
| PC1 | PC2 | |||||
| ORAC | −0.07164693 | 0.66194871 | ||||
| DPPH | 0.55538337 | 0.01775146 | ||||
| TPC | −0.38219393 | 0.00680021 | ||||
| TFC | −0.4869898 | 0.29703139 | ||||
| TGC | 0.09657944 | 0.65004107 | ||||
| TAC | 0.54208594 | 0.22512553 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega-Galvez, A.; Gomez-Perez, L.S.; Zepeda, F.; Vidal, R.L.; Grunenwald, F.; Mejías, N.; Pasten, A.; Araya, M.; Ah-Hen, K.S. Assessment of Bio-Compounds Content, Antioxidant Activity, and Neuroprotective Effect of Red Cabbage (Brassica oleracea var. Capitata rubra) Processed by Convective Drying at Different Temperatures. Antioxidants 2023, 12, 1789. https://doi.org/10.3390/antiox12091789
Vega-Galvez A, Gomez-Perez LS, Zepeda F, Vidal RL, Grunenwald F, Mejías N, Pasten A, Araya M, Ah-Hen KS. Assessment of Bio-Compounds Content, Antioxidant Activity, and Neuroprotective Effect of Red Cabbage (Brassica oleracea var. Capitata rubra) Processed by Convective Drying at Different Temperatures. Antioxidants. 2023; 12(9):1789. https://doi.org/10.3390/antiox12091789
Chicago/Turabian StyleVega-Galvez, Antonio, Luis S. Gomez-Perez, Francisca Zepeda, René L. Vidal, Felipe Grunenwald, Nicol Mejías, Alexis Pasten, Michael Araya, and Kong Shun Ah-Hen. 2023. "Assessment of Bio-Compounds Content, Antioxidant Activity, and Neuroprotective Effect of Red Cabbage (Brassica oleracea var. Capitata rubra) Processed by Convective Drying at Different Temperatures" Antioxidants 12, no. 9: 1789. https://doi.org/10.3390/antiox12091789